
R. Kader
Department of Food and Nutrition, National College of Home Economics, Dhaka, Bangladesh
A. Yousuf
Department of Chemical Engineermg and Polymer Science,
Shah Jalal University of Science and Technology, Sylhet-3 1 14, Bangladesh
M.M. Hoq
Department of Microbiology, University of Dhaka, Dhaka, Bangladesh
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 6 | Page No.: 855-860







ABSTRACT
Rhizopus sp. a mould of mucor family, excrete lipase when cultured on lipolytic media. The Rhizopus sp. produced a larger clear zone on tributyrin agar medium suggesting its esterase activity. It was further investigated in liquid medium in order to optimize the lipase production conditions under shake culture. Lipase production was found to be maximum with medium containing maltose (1%) and peptone (5%) as carbon and nitrogen sources, respectively with Rhizopus sp. The enzyme production was profoundly influenced by initial pH of the medium and optimum value of this parameter was found to be 6.0. Maximum enzyme production was obtained at 30 °C with a shaking rate of 200 rpm. Ca2+ was found to stimulate lipase production, while it was strongly inhabited by Hg2+. Lipase production was increased about 23.7% under optimized cultivation conditions over olive oil-peptone medium.
PDF Abstract XML References Citation
How to cite this article
R. Kader, A. Yousuf and M.M. Hoq, 2007. Optimization of Lipase Production by a Rhizopus MR12 in Shake Culture. Journal of Applied Sciences, 7: 855-860.
DOI: 10.3923/jas.2007.855.860
URL: https://scialert.net/abstract/?doi=jas.2007.855.860
DOI: 10.3923/jas.2007.855.860
URL: https://scialert.net/abstract/?doi=jas.2007.855.860
INTRODUCTION
Lipases (Triacylglycerol hydrolase, EC 3.1.1.3) are ubiquitous among microbes, plants and animals which catalyze the hydrolysis of ester linkages of triglycerides to form glycerol and fatty acids. Traditionally, these microbial lipases have been used in the food and detergent industry for the ripening of the cheese and as laundry detergent additives. Recently these enzymes have become of interest to the chemical, pharmaceutical and leather industries (Gulati et al., 2005; Gunstone, 1999) because of their ability to hydrolyze ester bonds, trans-esterify triglycerides, resolve racemic mixtures (Muralidhar et al., 2001; Reetz, 2002) and synthesize a variety of stereo-specific esters, sugar esters, thioesters and amides (Singh et al., 2003; Dellamora-Otiz, 1997).
Because of huge variation in applications, it has been renewed interest in the development of sources of lipase. Choo et al. (1998) reported that though numerous species of bacteria, yeasts and moulds produce lipase with different enzymological properties and specificities, moulds are known to be more potent lipase producer. These microorganisms produce lipase both by solid substrate and submersed fermentation (He et al., 2004). The best results in the production of lipase were obtained in the present of carbon and nitrogen sources (Fadiloğlu and Erkmen, 2002). Alonso et al. (2005) studied the lipase production by a brazilian wild srain of Yarrowia lipolytoca at different stirring speeds and air flow rates. They detected maximum lipase activity at 200 rpm.
Factor affecting microbial extracellular lipase production have been widely studied in bacteria (Lawrence et al., 1967; Mates and Sudakevitz, 1973), moulds (Chander et al., 1980; Chopra and Chander, 1983) and yeasts (Ota et al., 1968). Recently Kshmiri et al. (2006) cultivated a new strain Trichoderma viride for lipase production in shake flasks at 30±1°C and highest specific growth rate was observed between 6 and 18 h, though maximum fungal biomass was present at 13.6 g L–1 at 60 h. Few studies have been conducted with Rhizopus to examine the control of lipase production and the growth of the fungus. This study was, therefore, initiated to investigate the effect of different growth conditions supplemented with various nitrogen and carbon sources on the lipase production by Rhizopus MR12.
MATERIALS AND METHODS
The Rhizopus sp. was isolated from decomposed of mushroom culture. The organism was grown on Potato Dextrose Agar (PDA) plates or slants at 30°C and pH 5.5 for 2-3 days and stored at 4°C, with sub culturing every 3 or 4 weeks interval. All experiments were conducted in the laboratory of the Department of Microbiology, University of Dhaka, Bangladesh.
Media and shake flask culture condition: The basal medium contained the nitrogen source 5%, (NH4)2SO4, 0.1%; K2HPO4, 0.1% and different concentration of carbon sources (Maltose, lactose, xylose, glucose and glycerol) in 100 mL distilled water. The initial pH of the medium was adjusted to pH 5.5.
A culture medium of 50 mL (in baffled-Erlenmeyer flask) was inoculated with the fungal mycelia from 23 cm2 area if a PDA plate containing a mat like mycelia of 2-3 days growth of Rhizopus MR12. For each treatment at least two replica flasks ware made. The flasks were then shaken at 30°C on an orbital shaker (Gallenkamp, Germany) at 150 rpm for 5 days. The culture filtrate was assayed for lipase activity and other compounds.
Enzyme assay: Lipase activity was assayed by titrating fatty acid liberated from olive oil due to lipase activity with 0.05 M NaOH as describe by Ota et al. (1968). One unit of lipase activity was defined as the amount of lipase which liberated 1 μmol of fatty acid per minute at 30°C.
Optimization of cultural conditions for lipase production: Lipase production was optimized by altering various physico-chemical and cultural conditions observing the effect after 8 h of incubation at different temperature.
| • | The effect of incubation period on lipase production from Rhizopus sp. was studied measuring the enzyme activity, growth and extra cellular protein at different incubation period (8, 16, 24, 32, 40, 48, 56 and 72 h). |
| • | The effect of various supplements on lipase production was studied by adding different carbon sources (glucose, maltose, lactose, sorbitol, xylose, glycerol at an amount of 1% and olive oil at an amount of 2%), nitrogen sources (peptone, yeast extract, meat extract, urea, casein, corn steep liquor at an amount of 5%), metal ions (CaCl2, ZnCl2, MgCl2, FeCl2, BaCl2, HgCl2, con. 1 mM) in the basal medium. |
| • | Since the lipase production was maximum in the maltose (carbon source) and peptone (nitrogen source), different concentration of maltose (0.5, 1, 2%) and peptone (1, 3, 5%) were used to determine optimum concentration of them. |
| • | The effect of incubation temperature on the production of lipase was observed at different temperature (25, 28, 30, 33, 35, 40°C) with 1% maltose as the carbon source and 5% peptone as the nitrogen source. |
| • | The effect of initial pH of the culture was investigated using shake flask cultivation with 1% maltose as the carbon source and 5% peptone as the nitrogen source. The initial pH of the culture medium was adjusted to (3.0, 4.0, 5.0, 5.5, 6.0, 7.0, 8.0 and 9.0) with 1 M HCl and 1 M NaOH. |
| • | The effect of shaking rates was carried out at different values (120, 140, 160, 180 and 200 rpm). |
Others analysis: Soluble protein in the culture filtrate was analyzed by the method of Bradford (1976). Total Dry Mass (TDM) in the filtrate medium was determined by the method of Hoq et al. (1984).
RESULTS AND DISCUSSION
The optimum incubation period for lipase production (Fig. 1a), growth and extracellular protein section was found to be 30 to 32 h (Fig. 1b). The kinetics of growth and protein secretion was correlated.
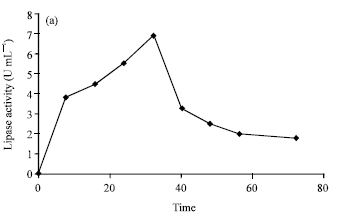 | |
| Fig. 1a: | Effect of incubation period on the production of lipase by Rhizopus sp. |
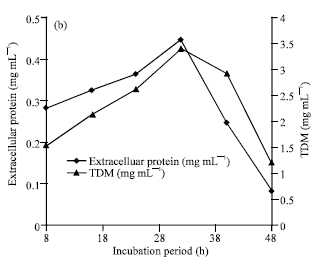 | |
| Fig. 1b: | Effect of incubation period on the production of extracellular protein and growth (Total Dry Mass- TDM) by Rhizopus sp. |
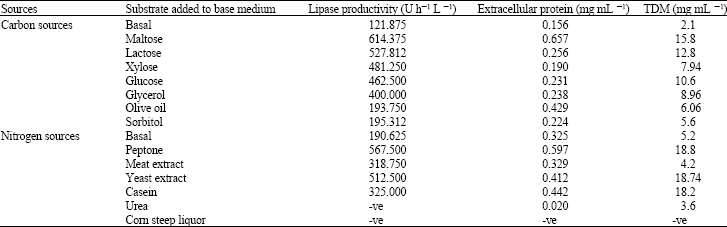
| Table 1: | Effect of various carbon and nitrogen sources on the production of lipase and extracellular protein by Rhizopus sp. |
 | |
The production of enzyme was found to be growth associated as the lipase activity increased with the increase in growth. This type of shorter incubation period is unique as of the previous reports indicated that optimum incubation period for lipase production by most fungi was around 72 h or longer (Samad et al., 1990; Akhter et al., 1980).
The effect of carbon sources on the production of lipase by Rhizopus was tested by using glucose, maltose, lactose, sorbitol, xylose, glycerol at an amount of 1% and olive oil at an amount of 2% as depicted in Table 1. In the present study, lipase productivity, production of soluble protein and biomass production was observed maximum with maltose followed by lactose. This finding agreed with Nakashima et al. (1988), who observed similar effect with Rhizopus chinensis. Lipase production in the media on olive oil was much lower than in media containing carbohydrates. This decrease of lipase production is comparable with that reported by Chander et al. (1980) who concluded that butter oil and olive oil inhibited lipase activity by 53 and 63%, respectively, similar observation were made both Lawrence et al. (1967) and Eittenmiller et al. (1970). As maltose supported high enzyme production than olive oil or fat, so mass production in water soluble media will be advantageous, because keeping the lipid substrate under emulsified condition needs vigorous agitation as opposed to soluble substrates. Olive oil was found to decrease lipase production similarly might be due to the inhibition by fatty acid concentration, which was liberated during the hydrolysis of triglycerides (Omer et al., 1987).
Among the nitrogen sources tested the maximum lipase (567.5 U h–1 L–1) and extracellular protein (0.597 mg mL–1) production and growth (18.8 g mL–1) was obtained on peptone (Table 1). Urea and ammonium sulphate failed to support lipase production by the fungi as they did with R. oligosporus (Nahas, 1988) and Acremonium stricum (Okele and Okolo, 1990) respectively. These indicate that organic nitrogen sources were preferred to inorganic ones for lipase production.
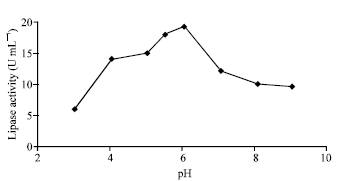 | |
| Fig. 2: | Effect of pH on the lipase activity |
In the present study corn steep liquor was failed to support both growth and synthesis of the enzyme, although it was a good nitrogen sources for lipase production by Humicola lanuginosa (Omar et al., 1987). So from these observations it can be stated that the lipase production was highly substrate specific.
The pH dependency of the culture played an important role for both growth of the fungi and the production of enzymes. The optimum pH for lipase production by Rhizopus sp. was found to be 6.0 (Fig. 2). During the growth of Rhizopus the pH was found to fluctuate in a narrow range. The change in medium pH may be compounds in the culture conditions employed.
Incubation temperature has been found to be significant controlling factor for enzyme production. The optimum incubation temperature for growth and lipase production by the fungi was found to be 30°C (Fig. 3). Most of the industrial application of microbial lipases for the hydrolysis of glycerides, interesterification of fatty acid moieties and so on, basically involved the use of mesophilic enzymes. Lipases of various species Rhizopus have been purified and characterized, including Rhizopus delemar (Iwai and Tsuijisaka, 1974), R. chinensis (Nakashima et al., 1988), R. arrhizus (Benzonana, 1974) and R. rhizopodifornis (Samad et al., 1990).
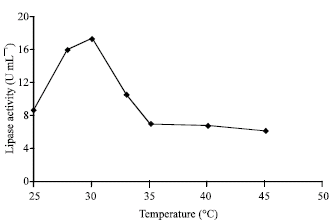 | |
| Fig. 3: | Effect of temperature on the production of lipase by Rhizopus sp. |
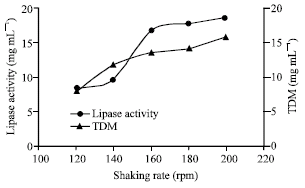 | |
| Fig. 4: | Effect of agitation rates on growth and lipase production |
However with the exception of R. rhizopodiformis (Nahas, 1988; Razak et al., 1989) reported that most of the Rhizopus species are mesophilic fungi.
Usually shaking rate i.e., agitation/aeration had profound influenced on the lipase production by aerobic microorganisms as increase in shaking rate increase the availability of dissolved oxygen. In addition shaking may also create condition of higher availability of the carbon sources to microorganisms (Nahas, 1988). In the present study, enzyme production was increased with increase in shaking rates (Fig. 4). This type of result was observed in case of mesophilic R. oryzae (Razak et al., 1989), A. oryzae (Ohnishi et al., 1994). But incase of R. oligosporus static incubation was required for maximum lipase production. This may be due to the denaturing effect of stress on the enzyme protein. The presence of Ca2+ in the culture medium was shown to stimulate lipase activity. Other ions such as Fe3+, Zn2+ slightly increased lipase production while Hg2+, Ba2+ and Mg2+ decreased the lipase production (Fig. 5) as happened with Humicola lanuginosa on sorbitol corn steep liquor (Omar et al., 1987). Metal ions generally form complexes with ionized fatty acids, changing their solubility and behavior at the interface. Lipase production was maximum in maltose and peptone and their optimum concentration as shown in Fig. 6 and 7.
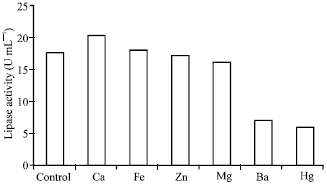 | |
| Fig. 5: | Effect of metal ions on the production of lipase by Rhizopus sp. |
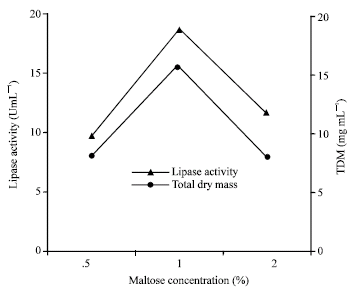 | |
| Fig. 6: | Effect of different concentration of maltose on the growth and production of lipase by Rhizopus sp. |
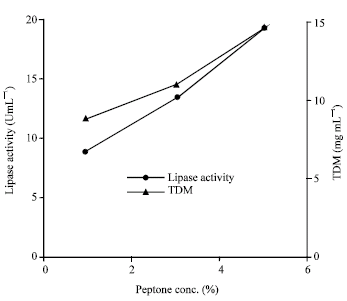 | |
| Fig. 7: | Effect of different concentration of peptone on the growth and production of lipase by Rhizopus sp. |
With the optimized medium (1% maltose and 5% peptone with CaCl2) maximum amount of lipase production (24.32 U mL–1) was possible within 32 h cultivation at 30°C with the pH maintained within 6.0-7.0 (Fig. 8).
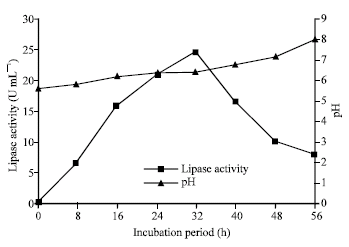 | |
| Fig. 8: | Lipase production and change of pH in the culture under optimized cultivation conditions (30°C) |
The results of the present study revealed that the maximum amount of enzyme is produced after 30 to 32 h of incubation period at 30°C. This shorter incubation period and ambient temperature of lipase production offers biotechnological exploitation of this organism. Purified lipase of this fungus can be used in tanneries and detergent as cleansing aid in our industries.
CONCLUSIONS
Modification and improvement of lipase production by Rhizopus sp. will lead to new and useful applications for this organism. More over it was observed that enzyme production was highly growth associated. Hence further study relating to increase in growth rate may be helpful to increase the enzyme production. It was also observed that cultivation conditions greatly affected lipase production and optimization of the cultivation factors improved lipase production. Thus optimized conditions for maximum enzyme production in the shake culture study will be useful for lipase production in controlled conditions of bioreactor.
REFERENCES
- Akhtar, M.W., A.Q. Miraz and M.D.I. Chughtai, 1980. Lipase induction in Mucor hiemalis. Applied Environ. Microbiol., 18: 257-263.
Direct Link - Alonso, F.O.M., E.B.L. Oliveira, G.M. Dellamora-Otiz and F.V. Perira-Meirelles, 2005. Improvement of lipase production at different stirring speeds and oxygen levels. Braz. J. Chem. Eng., 22: 9-18.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Choo, D.W., T. Kurihara, T. Suzuki, K. Soda and N.A. Esaki, 1998. Cold adapted lipase of an Alaskan psychrotroph, Pseudomonas sp., strain B11-1: Gene cloning and enzyme purification and characterization. Applied Environ. Microbiol., 64: 486-491.
Direct Link - Dellamora-Otiz, G.M., R.C. Martins, W.L. Rocha and A.P. Dias, 1997. Activity and stability of a Rhizomucor miehei lipase in hydropkobic media. Biotechnol. Applied Biochem., 26: 31-37.
Direct Link - Fadilo-Lu, S. and O. Erkmen, 2002. Effects of carbon and nitrogen sources on lipase production by Candida rugosa. Turk. J. Eng. Environ. Sci., 26: 249-254.
Direct Link - Gulati, R., J. Isar, V. Kumar, A.K. Prasad, V.S. Parmar and R.K. Saxena, 2005. Production of a novel alkaline lipase by Fusarium globulosum using neem oil and its applications. Pure Applied Chem., 77: 251-262.
CrossRefDirect Link - Gunstone, F.D., 1999. Enzymes as biocatalysts in the modification of natural lipids. J. Sci. Food Agric., 79: 1535-1549.
Direct Link - Kshmiri, M.A., A. Adnan and B.W. Butt, 2006. Production, purification and partial characterization of lipase from Trichoderma viride. Afr. J. Biotechnol., 5: 878-882.
Direct Link - Lawrence, R.C., T.F. Fryer and B. Reiter, 1967. Rapid method for the quantitative estimation of microbial lipase. Nature, 25: 1264-1265.
CrossRefDirect Link - Muralidhar, R.V., R. Marchant and P. Nigem, 2001. Lipases in racemic resolutions. J. Chem. Technol. Biotechnol., 76: 3-8.
Direct Link - Ohnishi, K., Y. Yoshida and J. Sekiguchi, 1994. Lipase production by Aspergillus oryzae. J. Ferment. Bioeng., 77: 490-495.
Direct Link - Okeke, C.N. and B.N. Okolo, 1990. The effect of cultural conditions on the production of lipase by Acremonium stricum. Biotechnol. Lett., 12: 747-750.
Direct Link - Reetz, M.T., 2002. Lipases as practical biocatalysts. Curr. Opin. Chem. Biol., 6: 145-150.
CrossRefDirect Link - Samad, M.Y.A., A.B. Salleh, C.A.N. Razak, B.K. Ampon, W.M.Z. Yunus and M. Basri, 1990. A lipase from a newly isolated thermophilic Rhizopus rhizopodiformis. World J. Microbial. Biotechnol., 6: 390-394.
Direct Link