
L.C. Saalu
Department of Anatomy, College of Health Sciences, Igbinedion University Okada, University of Ibadan, Ibadan, Nigeria
V.A. Togun
Department of Animal Production and Health, Faculty of Agricultural Sciences, University of Ibadan, Ibadan, Nigeria
A.O. Oyewopo
Department of Anatomy, Faculty of Basic Medical Sciences, Ladoke Akintola University of Technology, Ogbomoso, University of Ibadan, Ibadan, Nigeria
Y. Raji
Department of Physiology, College of Medicine, University of Ibadan, Ibadan, Nigeria
Journal of Applied Sciences
Year: 2006 | Volume: 6 | Issue: 14 | Page No.: 2889-2894







ABSTRACT
The study investigated the effect of melatonin on artificial crytorchidism in Sprague-Dawley rats. Forty rats weighing 220-270 g were divided into five treatment groups A, B, C, D and E with group A as control. The rats were rendered unilaterally (B and D) or bilaterally (C and E) cryptorchid by anchoring the upper pole of the testis to the abdominal wall. Groups D and E in addition received 0.7 mg kg-1 body weight of melatonin intraperitoneally between 9.00-10.00 am daily for 56 days. The control group gained 40% of their initial body weight while the mean weight losses of the cryptorchids were 27 and 23% (unilateral and bilateral without melatonin) and 45 and 39% (unilateral and bilateral with melatonin). Mean Paired Testes Weight (PTW) of control rats did not differ significantly (p>0.05) from those of groups B and D (unilateral cryptorchids with and without melatonin respectively) and E (bilateral cryptorchid with melatonin) but was significantly higher than the mean PTW of group C (bilateral cryptorchid without melatonin respectively). The ipsilateral testis without melatonin was lower in weight than the contralateral testis (B) while the weights were equal in melatonin treated rats (D). The mean PTW of bilateral cryptorchids with melatonin (E) was higher than the PTW of bilateral cryptorchids without melatonin. There was no sperm cell in the ipsilateral cauda epididymis of group B and the bilateral epididymides of group C rats that did not receive melatonin treatment (azoospermia). The epididymis of groups D and E rats, with melatonin administration had significantly (p<0.05) lower spermatozoa concentration (oligospermia) than the control rats. There was no significant (p>0.05) difference between epididymal sperm concentration and motility of the control rats compared with the contralateral testis of groups B and D rats. Motility of cryptorchid epididymal sperm was slow and non-linear. It was concluded that artificial cryptorchidism negatively affected metabolic activities in male Sprague Dawley rats. The contralateral testes of artificial, unilateral cryptorchids were not significantly (p>0.05) affected by the negative effect of artificial cryptochidism on the ipsilateral testes. Artificial cryptorchidism did not affect blood plasma testosterone level. Melatonin alleviated the deleterious effect of artificial cryptorchidism.
PDF Abstract XML References Citation
How to cite this article
L.C. Saalu, V.A. Togun, A.O. Oyewopo and Y. Raji, 2006. Artificial Cryptorchidism and the Moderating Effect of Melatonin (N-acetyl. 5 methoxy tryptamin) in Sprague-Dawley Rats. Journal of Applied Sciences, 6: 2889-2894.
DOI: 10.3923/jas.2006.2889.2894
URL: https://scialert.net/abstract/?doi=jas.2006.2889.2894
DOI: 10.3923/jas.2006.2889.2894
URL: https://scialert.net/abstract/?doi=jas.2006.2889.2894
INTRODUCTION
Failure of the testis to descend into the scrotum is called cryptorchidism. The word cryptorchid literally means a testis, which is hidden from view (Senger, 1999). Cryptorchidism is generally associated with male infertility; the resulting testicular damage is in part, a consequence of increased generation of free radicals and oxidants (Grisham and Mc Cord, 1986). According to Waites and Setchell (1990), increasing testicular temperature above the normal level has been reported to alter spermatogenesis in mammals due to the effect of heat on the seminiferous epithelium. Surgically induced cryptorchidisn has been reported to result in time dependent reduction in cauda epididymal sperm concentration and motility (Oyewopo and Togun, 2005). Bilateral cryptorchidism results in sterility. Infertility, with cryptorchidism as one of its causes, is a personal and social problem in man as well as a major cause of reproductive losses in animal production. According to Reiter (1993) melatonin (N-acetyl. 5-methoxy tryptamine), secreted by the pineal gland, is the most effective antioxidant and free radical scavenger yet to be studied. It is non-toxic and has the capacity to easily cross morphophysiological barriers to enter subcellular components.
Melatonin has been reported to stimulate testicular growth, enhance puberty, increase quality and quantity of spermatozoa produced as well as improves the viability of post-thawed spermatozoa (Chemineau et al., 1992; 1996; 1998; Kaya et al., 2001). The reduction of oxidative stress by melatonin has been reportedly due to its being an effective scavenger of both highly toxic hydroxyl radical, produced by the electron reduction of oxygen and the peroxyl radical that is sufficiently toxic to propagate lipid peroxidation generated during oxidation of unsaturated lipids (Reiter, 1993).
The aim of the present study was to investigate the action of melatonin, an antioxidant with hypothermic characteristic, as a moderator of testicular damage in artificial cryptorchidism as well as establish the relative status of contralateral testis in the unilateral cryptorchid animal.
MATERIALS AND METHODS
Animals: Adult male Sprague-Dawley rats weighing 220-270 g were used for the study. The animals were housed in wire mesh cages under standard environmental conditions with the provision of 12 h light and 12 h darkness. Rat cubes (Pfizer feeds Nig. Ltd, Ibadan, Nigeria) and water were provided ad libitum.
Experimental protocol: Forty male Sprague-Dawley rats were weighed and divided into five groups of eight rats each. Group A served as the control and the rats were neither rendered cryptorchid nor treated with melatonin. Groups B, C, D and E served as the experimental groups in which rats were rendered unilaterally (B and D) or bilaterally (C and E) cryptorchid. Artificial cryptorchidism was performed by anchoring the upper pole of the testis to the anterior abdominal wall, using a suture passing through the connective tissue of the caput epididymis as described by Jegou et al. (1984). Groups D and E rats in addition had 0.7 mg kg-1 body weight of melatonin injected intraperitoneally, daily between 9:00-10:00 am for 56 days.
Autopsy and organ weights: At the end of the experimental period, each rat was weighed and sacrificed by decapitation. The testes and epididymides were dissected and weighed immediately.
Sperm characteristics: The testes from each rat were carefully exposed and removed. They were trimmed free of the epididymides and adjoining tissues. From each separated epididymis, the cauda part was removed and placed in a beaker containing 1 mL physiological saline solution. Each section was quickly macerated with a pair of sharp scissors and left for a few minutes to liberate its spermatozoa into the saline solution. Sperm motility, concentration and progressive motility were determined as earlier described (Raji et al., 2003, 2005, 2006). Semen drops were placed on the slide and two drops of warm 2.9% sodium citrate were added. The slide was covered with a cover slip and examined under the microscope using X40 objective for sperm motility. Sperm count was done under the microscope using improved Neubauer haemocytometer.
Estimation of plasma levels of testosterone: Plasma testosterone concentrations were estimated using the Enzyme Immunology Assay (EIA) method as earlier described (Raji et al., 2005, 2006). Plasma samples collected were stored at-20°C until assayed. The EIA kits used were obtained from Immunometrics (London U.K) and contained testosterone EIA substrate reagents and EIA quality control samples. A quality control sample was run for the hormone at the beginning and at the end of the assay to ascertain acceptability with respect to bias and within assay variation. The EIA kit used had a sensitivity level of 0.3 nmol L-1 (0.1 ng mL-1). The intra and inter assay variations were 11.00 and 10.10%, respectively.
Statistical analysis: Data were expressed as mean±SEM. They were subjected to analysis of variance (ANOVA). Statistical significance between the various groups was separated by t-test (SAS, 2002).
RESULTS
Body weight changes: Table 1 shows that rats in the control group significantly (p<0.05) increased in weight by 34% of their initial mean live weight. All cryptorchid groups lost weight up to between 23-45% of their initial weight. However the weight loss by each of the cryptorchid groups that received melatonin was higher than the losses by each of the cryptorchid groups that did not receive melatonin.
| Table 1: | Live weight changes in cryptorchid rats |
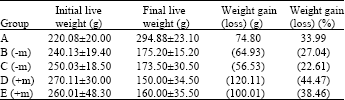 | |
| A = Control, B = Unilateral cryptorchid, C = Bilateral cryptorchid, D = Unilateral cryptorchid, E = Bilateral cryptorchid, (-m) = without melatonin administration, (+m) = with melatonin administration | |
| Table 2: | Mean testicular and mean relative testicular weights |
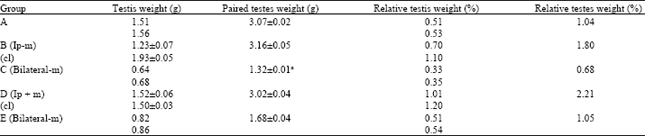 | |
| A = control, B = Unilateral cryptorchid, C = Bilateral cryptorchid, D = Unilateral cryptorchid, E = Bilateral cryptochid, (-m) = Without melatonin administration, (+m) = With melatonin administration, a: Significantly (p<0.05) lower than control | |
| Table 3: | Spermatozoa characteristics in cryptorchid rats’ cauda epididymides |
 | |
| A = Control, B = Unilateral cryptorchid C = Bilateral cryptorchid, D = Unilateral cryptorchid, E = Bilateral cryptochid, (-m) = without melatonin administration, (+m) = with melatonin administration, a1 = Rapid linear progressive motility, b1 = Show sluggish linear or non-linear motility, abMeans along each row with same superscript are not significantly different (*) significantly lower than control value (p<0.01) | |
| Table 4: | Testosterone levels in cryptorchid rat plasma (ng mL-1) means±SEM |
 | |
Testes weights and relative weights: Table 2 shows that there were no significant (p>0.05) differences between the Paired Testes Weights (PTW) of groups B and D rats compared to the control rats. The mean weight of the ipsilateral testes in the unilateral cryptorchid rats that did not receive melatonin was lower (1.23 vs 1.93 g) than the contralateral testis whereas, the mean weight of the ipsilateral testes of the rats that received melatonin was about equal (1.52 vs 1.50) to the mean weight of the contralateral testes. The mean weight of the ipsilateral testes of rats that did not receive melatonin was lower (1.23±0.07 vs 1.52±0.06) than that of rats that received melatonin. The mean PTW of the bilateral cryptorchids that did not receive melatonin was lower (1.32±0.01 vs 1.68±0.04 g) than the mean PTW of the group that received melatonin. The bilateral cryptorchid that did not receive melatonin had the least PTW and mean relative weight among the groups, being significantly (p<0.05) lower than the control.
Spermatozoa concentration: Table 3 shows that there were no significant differences between the sperm concentration of the control (group A) and those of contralateral groups B and D. The epididymides of the rats with ipsilateral testes and the bilateral cryptorchids that did not receive melatonin did not have any sperm cell (azoospermia). However the epididymides of ipsilateral testes and bilateral cryptorchids with melatonin treatment were oligospermatic (Table 3).
Spermatozoa motility: There were no significant (p<0.05) differences between the sperm motility observed in the epididymides of control rats and those of the contralateral testes (B or D) with or without melatonin treatment. The bilateral cryptorchids with melatonin treatment did not significantly (p>0.05) differ from the ipsilateral group with melatonin but they both had motility values significantly (p<0.05) lower than the control and the contralateral epididymides (Table 3).
Progressivity of spermatozoa motility: The sperm cells from the cauda epididymides of the control and contralateral testes showed progressive linear movement while those from the bilateral and ipsilateral with melatonin, showed sluggish, linear movement (Table 3).
Testosterone levels in plasma: There was no significant (p>0.05) difference between the control rats and any of the other groups, which did not significantly (p>0.05) differ from one another (Table 4).
DISCUSSION
The results showed that artificial cryptorchidism, both unilateral and bilateral, impaired growth and reproductive capacity of male Sprague Dawley rats. The 34% increase in live weight of rats in the control group at the end of the study probably indicates that the rats were still in the active growth phase. The loss in weight observed in all the cryptorchid rats (22.61-44.47%) implicates cryptorchidism with a negative effect on the body metabolic process in rats. This situation can be conjecturally attributed first to the physiological phenomenon of compensatory hypertrophy in the groups B and D rats, which were unilateral cryptorchids and where the contralateral testes were subjected to compensatory hypertrophy. This is supported by the report of Ezeasor (1985) that the single descended testicle undergoes compensatory hypertrophy. The process of hypertrophy would aid the descended testes to perform the functions of the normal two testes. In this case, there would be the diversion of available nutrients, which would be channeled towards the hypertrophy process of the testis in preference to body growth. The other explanation could be based on the fact that the procedure for achieving artificial cryptorchidism in the rats would confer a conspicuous level of stress on the rats. This stress could affect the metabolic process of all the cryptorchids, leading to losses in live weights observed in this study. The conspicuous increase in weight loss from 27.04% (group B) to 44.47% (group D) can be accounted for by the administration of melatonin to the unilateral cryptorchids in group D. According to Chemineau et al. (1992, 1996, 1998) melatonin implant resulted in significant stimulation of testicular growth to result in early maturity, mass increase in testis weight and sperm production. Such functions of melatonin would cause a synergistic effect with the compensatory hypertrophy process in unilateral cryptorchids, to increase the quantity of nutrients diverted from body growth to both testicular growth and functions in the melatonin treated, unilateral cryptorchids (group D) than those untreated (group B) rats.
The non-significant difference between the control group and the unilateral cryptorchids in Paired Testes Weight (PTW) despite the significantly lower live weight of the unilateral cryptorchids could probably be due to the compensatory hypertrophy phenomenon and the ameliorative effect of melatonin administration (group D), which enhanced the weights of the contralateral testes. This is in agreement with earlier findings (Ezeasor and Singh, 1989; Fred 1997, Inegedu et al., 2005). The lower testes weights in bilateral cryptorchids compared to the control also corroborate the report of Ono and Sofikitis (1997). Frandson, (1976), Guyton and Hall (1996) and Senger (1999) reported degenerative changes in the seminiferous epithelium of testes lying within the core body temperature, due to the effect of heat on the spermatogenic cells within the epithelium leading to loss in testicular weight.
That the mean PTW of group E rats, which had a lower mean final live weight and which received melatonin treatment was higher than the mean PTW of group C rat with higher final live weight is an indication of the ameliorating as well as testicular growth stimulating effects of melatonin administration. These effects also accounted for the relative testes weight (testes weight as a percent of body weight) of group E rats, which was equal to the value observed in the control rats contrary to that group C which was much lower. The same explanation could hold for the higher mean weight of the ipsilateral testes in group D than that of group B rats as well as their relative weights.
The non-significant difference between the values observed for epididymal sperm concentration and motility in the contralateral epididymides of groups B and D rats compared with the control, contradicts the report of Ono and Sofikitis (1997). Retention of the testes inside the abdomen results in disruption of spermatogenesis and impairment of steroidogenesis (Zhang et al., 2002). This results in poor testicular functions (Patkowski et al., 1992; Tellaloglus, 1994; Kawakami, 1995; Nambirajan et al., 2002). Attending degeneration of tubular epithelium, leaving only the interstitial structure of the testis, with consequential difficulty in the formation of viable spermatozoa have also been reported (Frandson, 1976; Guyton and Hall, 1996; Senger, 1999). That the epididymides of the ipsilateral (group B) and bilateral (group C) cryptorchid testes had no spermatozoa confirms the deleterious effect of cryprorchidism on the testes and their functions. Similar results on epididymal sperm concentration and motility have recently been reported (Oyewopo and Togun, 2005).
According to Pinart et al. (1999), the low sperm concentration indicated that unilateral cryptorchidism severely impaired sperm production of the scrotal testis. The observed low sperm concentration (oligospermia) and motility in the ipsilateral (group D) and bilateral (group E) epididymides with melatonin administration are significant improvements over the azoospermic group B (ipsilateral) and group C (bilateral) epididymides. This is a confirmation of the ameliorative effect of melatonin, which possesses hypothermic properties and highly effective anti-oxidant and free radical scavenger properties (Reiter, 1993, 1995). Thus the free radicals generated by the exposure of the testes to the core body temperature as well as the attending oxidative process conferred by artificial cryptorchidism have been alleviated by the administration of melatonin, which lowered the core body temperature that could reduce the detrimental effect of heat on the cryptorchid testes. This result supports the finding of Reiter (1998) that melatonin protected the tissues from oxidative damage.
The non-significant difference between the groups in plasma testosterone suggests that although all cell types of the testes were affected by cryptorchidism, the most prominent damage occurred in the seminiferous epithelium. Thus the Leydig cells, which secrete testosterone, were minimally affected by artificial cryptorchidism. This agrees with earlier findings (Frandson, 1976; Guyton and Hall, 1996; Senger, 1999).
In conclusion, artificial cryptorchidism negatively affected normal body weight of male Sprague Dawley rats. The contralateral testes of artificial, unilateral cryptorchids were not significantly affected by the negative effect of artificial cryptorchidism on the ipsilateral testes. Artificial cryptorchidism did not affect blood plasma testosterone level. Melatonin, to some extent, alleviated the deleterious effect of artificial cryptorchidism, whether unilateral or bilateral.
REFERENCES
- Chemineau, P. and B. Malpaux, 1998. Melatonine et reproduction chez les mammiferes d'elevage: La melatonine-Melatonin and reproduction in farm domestic mammals: Melatonin. Comptes Rendus Seances Societe Biologie Ses Filiales, 192: 669-682.
Direct Link - Jegou, B., R.A. Peake, D.C. Iraby and D.M. De Krester, 1984. Effects of the induction of experimental cryptorchidism and subsequent orchidopexy on testicular function in immature rats. Biol. Reprod., 30: 179-187.
Direct Link - Kawakami, E., T. Tsutsui, S. Saito, T. Kakimoto and A. Ogasa, 1995. Changes in peripheral plasma leutinizing hormone quality in normal and maturation. Lab. Anim. Sci., 45: 258-263.
Direct Link - Kaya, A., M. Aksoy, N. Baspinar, C. Yildiz and M.B. Ataman, 2001. Effect of melatonin implantation to sperm donor rams on post-thaw viability and acrosomal integrity of sperm cells in the breeding and non-breeding season. Reprod. Domestic Anim., 36: 211-215.
Direct Link - Nambirajan, L., S. Agarwala, A.K. Dinda and D.K. Mitra, 2002. Fertility and Unilateral undescended testis in the rat model. III. Ultrastructural changes in the contralateral descended testis. Pediatr. Surg. Int., 18: 276-280.
CrossRefDirect Link - Patkowski, D., M. Jelen and K. Creni, 1992. The natural course of cryptorchidism in rats. J. Pediatr. Surg., 27: 870-873.
Direct Link - Raji, Y., U.S. Udoh, O.O. Mewoyeka, F.C. Ononye and A.F. Bolarinwa, 2003. Implication of reproductive endocrine malfunction in male antifertility efficacy of Azadirachta indica extract in rats. Afr. J. Med. Sci., 32: 159-165.
PubMedDirect Link - Reiter, R.J., 1993. Interactions of the pineal hormone melatonin with oxygen-centered free radicals. Braz. J. Med. Biol. Res., 26: 1141-1155.
Direct Link - Reiter, R.J., 1995. A review of the evidence supporting melatonin's role as an antioxidant. J. Pineal Res., 18: 1-11.
Direct Link - Reiter, R.J., 1998. Oxidative damage in the central nervous system: Protection by melatonin. Prog. Neurobiol., 56: 359-384.
CrossRefDirect Link - Tellaloglu, S., A. Kadioglu, I. Kilicaslan, M. Alipour, T. Kadioglu and O. Ziylan, 1994. Cryptorchidism: Is orchidopexy always preventive treatment for infertility? Acta Chirurgica Hungarica, 34: 195-201.
Direct Link - Zhang, R.D., X.H. Wen, L.S. Kong, X.Z. Deng and B. Peng et al., 2002. Quantitative study of the effect of experimental unilateral cryptorchidism and subsequent orchidopexy on spermatogenesis in adult rabbit testis. Reproduction, 124: 95-105.
Direct Link - Raji, Y., O.S. Akinsomisoye and T.M. Salman, 2005. Antispermatogenic activity of Morinda lucida extract in male rats. Asian J. Androl., 7: 405-410.
CrossRefDirect Link