
Tahany A. Elzainy
Department of Microbial Chemistry, National Research Centre, Dokki, Cairo, Egypt
Thanaa H. Ali
Department of Microbial Chemistry, National Research Centre, Dokki, Cairo, Egypt
Journal of Applied Sciences
Year: 2006 | Volume: 6 | Issue: 6 | Page No.: 1389-1395







ABSTRACT
Extracts of nitrate-grown mats of Aspergillus niger NRRL3 catalyzed, with comparable efficiency, hydrolytic deamidation of both L-glutamine and L-asparagine in a similar manner under different experimental conditions. Other amides such as acetamide, nicotinamide and nicotinamide adenine dinucleotide could not be deamidated by these extracts. Optimum deamidation of the two amino acids by the extracts was recorded at pH 6 of 0.08 to 0.16 M phosphate buffer and 60°C. Exposure of the extracts to 60°C for 20 min in absence of the substrate revealed about 70% increase in each of the two activities. Fractionation of the extract proteins using DEAE Sephadex A-25 column chromatography showed that the glutaminase and asparaginase activities were detected in the same protein fractions with the same peak and with almost constant ratios. The ratio of the glutaminase activity to that of the asparaginase was approximately 0.7 to 1.0. The activities of the pooled fractions towards L-glutamine and L-asparagine showed similar behaviors at different pH values and at different temperatures. Both activities exhibited hyperbolic substrate saturation kinetics and the apparent km values were found to be 5.05 and 2.57x10-3 M for L-glutamine and L-asparagine, respectively. The sum of the two activities was much less than additive on using an equimolar mixture of L-glutamine and L-asparagine as substrates. Each of the two activities was not inhibited by the products of the reaction but inhibited by Mg++ or Hg++. Both activities were inactivated at almost the same rate on exposure to temperatures above 40°C and were retained in the freezer for some months. The same type of enzyme was also suggested to occur in a strain of Penicillium chrysogenum and in a strain of Penicillium politans.
PDF Abstract XML References Citation
How to cite this article
Tahany A. Elzainy and Thanaa H. Ali, 2006. Detection of the Antitumor Glutaminase-Asparaginase in the Filamentous Fungi. Journal of Applied Sciences, 6: 1389-1395.
DOI: 10.3923/jas.2006.1389.1395
URL: https://scialert.net/abstract/?doi=jas.2006.1389.1395
DOI: 10.3923/jas.2006.1389.1395
URL: https://scialert.net/abstract/?doi=jas.2006.1389.1395
INTRODUCTION
To our knowledge, some microorganisms non of which belongs to the filamentous fungi, have been reported to contain the amidohydrolase (E C 3.5.1.38) which has dual specificity towards L-glutamine and L-asparagine and which proved to have substantial effectiveness against a variety of tumors that have abnormally high requirements for L- glutamine and L- asparagine. These organisms are: I. Different pseudomonads such as: Pseudomonas species (Ramadan et al., 1964; Broome and Schekein, 1971); Pseudomonas fluorescens (Nikolaev et al., 1971; Pekhov et al., 1983, Huser et al., 1999), Pseudomonas aeruginosa (Soda, 1972); Pseudomonas acidovorans (Davidson et al., 1977); Pseudomonas 7A (Roberts, 1976; Roberts and Mc Gregor, 1991; Lubkowski et al., 1994; Jacob et al., 1997; Ortlund et al., 2000); Pseudomonas boreopolis 526 (Pekhov et al., 1983, 1984, 1985; Pekhov and Zanin, 1987) and Pseudomonas aurantiaca BKMB 548 (Kabanova et al., 1985; Lebedeva et al., 1988; Lebedeva and Berezov, 1995, 1997 ). II. Acinetobacter glutaminasificans (Holcenberg et al., 1972; Holcenberg, 1979; Kien and Holcenberg, 1981; Steckel et al., 1983; Ammon, 1988).
III. Achromobacter (Roberts et al., 1972; Spiers and Wade, 1979; Abuchowski et al., 1979). IV. One yeast characterized as Cryptococcus nodaensis has also been suggested to contain this enzyme (Sato et al., 1999).
According to Ortlund et al. (2000), the most studied representatives of this class of enzymes are those of Pseudomonas 7A and Acinetobactr glutaminasificans.
The present study provides data indicating detection, for the first time in a filamentous fungus of the glutaminase-asparaginase, and demonstrates its constitutive nature and some of its kinetic properties.
MATERIALS AND METHODS
Chemicals: L-glutamine, L-asparagine, acetamide, L-glutamic acid and L-aspartic acid are products of BDH Chemicals Ltd. Nicotinamide adenine dinucleotide (NAD) and nicotinamide are products of Sigma. DEAE Sephadex A-25 is from Pharmacia Fine Chemicals.
Organisms: The A. niger NRRL3, a strain of Penicillium chrysogenum and a strain of Penicillium politans were from the culture collection of the Department of Microbial Chemistry, National Research Centre, Cairo, Egypt.
Medium: The organisms were grown and kept on slants of solid modified Czapek Dox's medium containing (g L tap water): glucose, 30; NaNO3, 2.0; KH2 PO4, 1.0; Mg SO4 7 H2O, 0.5; KCl, 0.5 and agar 20.
Preparation of fungal extracts: The 4 days old mats, grown on liquid modified Czapek-Dox’s medium at 28°C were harvested by filtration, washed thoroughly with distilled water, blotted dry with absorbent paper then ground with cold washed sand in a chilled mortar and extracted with cold distilled water. The slurry so obtained was centrifuged at 5500 rpm for 10 min and the supernatant was used as the crude enzyme preparation.
Colorimetric determinations: NH3 was determined by direct nesslerization. Protein of the extracts was estimated by the method of Sutherland et al. (1949) and that of the eluted fractions was determined by UV absorption according to the method of Layne (1957).
Chromatographic analysis: Separation and identification of the amino acids were carried out by ascending paper chromatography technique as described by Smith and Seakins (1976) using Whatman No. 3 mm filter papers (46x57 cm ) and two solvent systems. Solvent I consisted of n-butanol: conc acetic acid: water; 60:15:30 and solvent II consisted of n-propanol: pyridine: water; 65:13:26. After spraying the dried chromatograms with a solution of 2% ninhydrine in absolute alcohol, the Rf values of the located spots were determined and were found to be: 0.24, 0.22, 0.33 and 0.27 in solvent I and 0.23, 0.18, 0.17 and 0.14 in solvent II for glutamine, asparagine, glutamic acid and aspartic acid, respectively.
Fractionation of the extract proteins using DEAE- Sephadex A-25 column chromatography: The crude extracts were heated at 60°C for 20 min, immediately cooled in ice, then centrifuged at 12000 rpm for 10 min at20°C. Ten milliliter of the supernatant was then loaded on a DEAE-Sephadex A-25 column (1.5x40 cm) which was equilibrated with 0.02 M phosphate buffer at pH 6. Elution of protein was then carried out by batch wise addition of 40 mL portions of increasing molarities (0.0-0.4 M ) of solutions of NaCl in 0.02 M phosphate buffer at pH 6. Fractions of 5 ml each were collected at room temperature (25°C) at a flow rate of about 27 mL h-1. At the end of the fractionation, the activity of each fraction was tested with L-glutamine and L-asparagine as substrates.
The assay reaction mixture of the eluted fractions contained (in 1 mL vol.): 0.4 mL of each fraction, 5.0 μmoles substrate and 80 μmoles phosphate buffer at pH 6. The time of the reaction was 30 min and the temperature was 40°C.
Specific activity was expressed as Fmoles NH3 released from 5.0 μmoles substrate per mg protein per min at 40°C.
Appropriate control reaction mixtures where the enzyme source or the substrate was omitted were used as blanks through out the work.
RESULTS AND DISCUSSION
Hydrolytic deamidation of L-glutamine and L-asparagine by extracts of Aspergillus niger NRRL3: Data presented in Fig. 1 show that when extracts of nitrate grown mats of A. niger NRRL3 were incubated with each of: L-glutamine, L-asparagine, L-glutamate, L-aspartate, acetamide, nicotinamide or NAD, as a substrate, at different pH values, ammonia was produced only in the two reaction mixtures that contained L-glutamine and L-asparagine even after prolonged (180 min) incubation period. This indicates that the amide linkages of only glutamine and asparagine were susceptible to hydrolysis by these extracts, while neither the same type of linkage of acetamide, nicotinamide and NAD nor the amino linkages of L-aspartate, L-glutamate and NAD could be hydrolyzed.
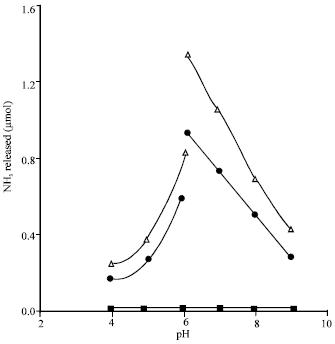 | |
| Fig. 1: | Hydrolytic deamidation of L-glutamine and L-asparagine by A. niger extracts at different pH values. Reaction mixture contained (in 1 mL vol): substrate, 5.0 μmol; extract protein, 2.73 mg; buffer, 80 μmol (citrate from pH 4 to pH 6 and phosphate from pH 6 to pH 9); temp, 40°C; time of the reaction, 30 min. NH3 formed: from L-asparagine (Δ), from L-glutamine (●), from L-glutamate, L-aspartate, acetamide, nicotinamide or NAD (■) |
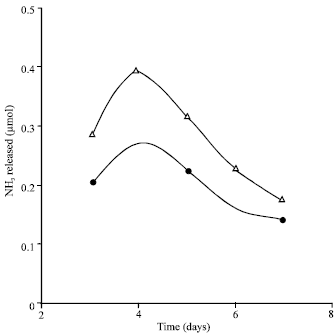 | |
| Fig. 2: | Activity with L-glutamine and L-asparagine as a function of the age of mats. Reaction mixture contained (in 1 mL vol): substrate, 5.0 μmol; extract protein, 2 mg. phosphate buffer pH 6, 80 μmol;. temp, 40°C; incubation period, 30 min. Specific activity: μmol NH3 formed per mg protein per 30 min; activity with L-asparagine (Δ), activity with L-glutamine (●) |
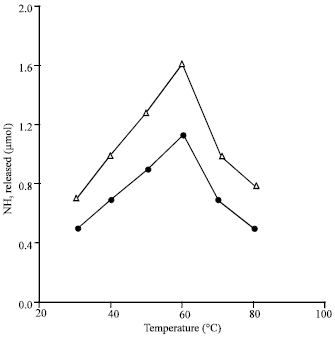 | |
| Fig. 3: | Dependence of the glutaminase and asparaginase activities on the temperature of the reaction. Reaction mixture contained (in 1 mL vol): substrate, 5.0 μmol; extract protein 2.4 mg. phosphate buffer at pH 6, 80 μmol; time of the reaction, 30 min and temp, as indicated; activity with L-asparagine (Δ), activity with L-glutamine (•) |
| Table 1: | Substrate specificity of the amidohydrolase pooled fractions |
 | |
| Reaction mixture substrate, 5 μmol; protein, 105 μg; phosphate buffer pH 6, 80 μmol; vol, 1 mL; temperature, 40°C; time, 30 min., *5 μmol L-asparagine+5 μmol L-glutamine were used as substrates in the reaction mixture that contained mixed substrates | |
| Table 2: | Pattern of inhibition and thermal inactivation of the glutaminase and sparaginase activities |
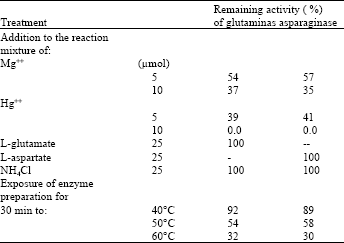 | |
| Note exposure of the enzyme preparation for 60 min to 40°C, 50°C or 60°C revealed about 85, 45 and zero % remaining activity, respectively | |
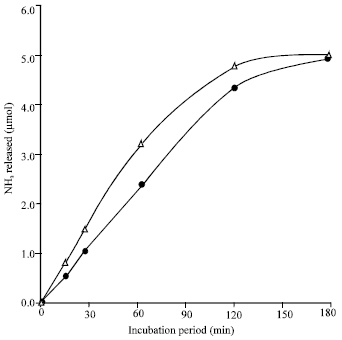 | |
| Fig. 4: | Completion of L-glutamine and L-asparagine deamidation reactions. Reaction mixture contained (in 1 mL vol): substrate, 5.0 μmol; extract protein, 2 mg; phosphate buffer at pH 6, 80 μmol; temp, 60°C; incubation period, as indicated. Activity with L-asparagine) (Δ), activity with L-glutamine (•) |
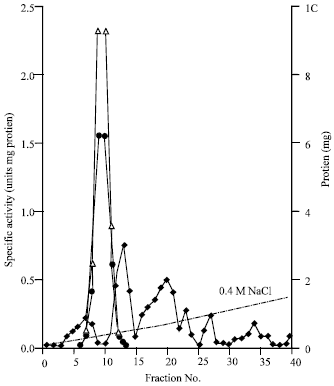 | |
| Fig. 5: | Elution diagram of the glutaminase and asparaginase using DEAE-Sephadex A-25 column chromatography. Asparaginase activity (Δ) glutaminase activity (●), protein (■) |
Chromatographic analysis of the two reaction mixtures in which NH3 was produced, after 3 h incubation, showed formation of glutamic acid and aspartic acid in the reaction mixture that contained L-glutamine and L-asparagine, respectively, as well as almost disappearance of the two substrates.
The Fig. 1 also shows that the two pH-activity profiles, the two pH optima (at pH 6) and the glutaminase -asparaginase ratio at the different pH values were almost similar. In addition, it appears that at pH 6, the two activities were higher in phosphate buffer than in citrate buffer.
Testing the glutaminase and asparaginase activities in some buffer systems including phosphate, Tris-acetate, phthalate and citrate revealed data indicating that phosphate buffer was somewhat more suitable for the two activities. Also testing the two activities at different molarities of the phosphate buffer (from 0.04 to 0.4) showed that optimum activities were obtained at a concentration of 0.08-0.16 M.
From these results it can be suggested that the extracts either contain one constitutively expressed amidohydrolase capable of hydrolyzing the two amino acids or contain two distinct deamidases which behaved similarly under the different experimental conditions tested.
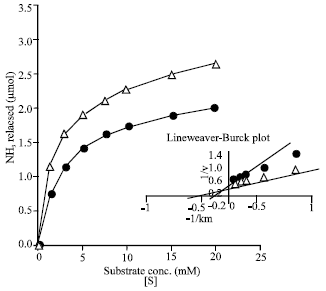 | |
| Fig. 6: | Effect of substrate concentration on the deamidase (s) activities. Reaction mixture contained (in 1 mL vol): substrate, as indicated; protein, 200 μg; phosphate buffer at pH 6, 80 μmol; temp, 40°C; incubation period, 30 min; activity with L-asparagine) (Δ), activity with L-glutamine (•) |
Dependence of the glutaminase and asparaginase specific activities on the age of the A. niger mats: Figure 2 shows that the highest specific activity with each of L-glutamine and L-asparagine was obtained on using extracts of four days old mats and also shows similarity of the two profiles and of the glutaminase- asparaginase ratio through out the growth period.
Temperature-activities relationships: Figure 3 demonstrates the pattern of dependence of L-glutamine and L-asparagine deamidation on the temperature of the reaction. From this figure, it appears that both L- glutamine and L-asparagine were optimally deamidated at 60°C and with almost the same activity profile. In this connection, when the extracts were exposed in absence of the substrate to 60°C, for 20 or 60 min the two activities were about 70% increased. These results indicate that presence of the substrate has no role in obtaining higher activity at 60°C (Fig. 3). Increase in activity due to exposure of the extracts to 60°C can be attributed either to the chemical nature of the enzyme(s), to elimination of certain inhibitory factor(s), to participation of certain activating factor(s) or to all these factors collectively.
Completion of the deamidation reactions: Figure 4 shows that by the end of three hours incubation of L-glutamine or L-asparagine with the extracts at the optimum pH and temperature, almost all the amounts of L-glutamine and L-asparagine were hydrolyzed. These results indicate that the products of each reaction did not inhibit the action of the deamidating enzyme.
Detection of the glutaminase and asparaginase activities in the same protein fractions during column fractionation of the extract proteins: Figure 5 demonstrates the elution diagrams of the glutaminase and asparaginase during fractionation of the extract proteins using DEAE Sephadex A-25 (Methods). As it appears from the diagram, the two activities were detected in the same protein fractions and with the same peak. In addition, the ratio of the glutaminase activity to the asparaginase activity in all these fractions was found to be almost constant and represents about 0.7 to 1.0. The highest specific activity recorded corresponds to about 160 fold purification.
Investigating the effects of pH and temperature on the two activities of the pooled fractions revealed data indicating that both activities were optimally effected, with the same profile, at pH 6 and at 50°C.
Substrate specificity of the deamidase (s): Data of Table 1 show that the amidohydrolase(s) of the pooled fractions exhibited activities only with L-glutamine and L-asparagine out of the tested compounds. The table also shows that the activity obtained on using equimolar mixture of L-glutamine and L-asparagine was non additive, suggesting possible binding of both substrates to the same catalytic site.
Substrate saturation kinetics of the amidohydrolase (s): Figure 6 demonstrates hyperbolic substrate saturation kinetics on using either L-glutamine or L-asparagine as a substrate for the pooled fractions enzyme. This Michael's-Menten relationship as well as proportionality of enzyme concentration-activity relationship with each of the two substrates (not cited data) indicate that the amidohydrolase can not be considered as an enzyme which has cooperative binding of any of the two substrates to multiple sites. The apparent km values for L-glutamine and L-asparagine were calculated from the Line-weaver Burk plots and were found to be 5.05 and 2.57x10-3 M, respectively (Fig. 6).
Pattern of inhibition and inactivation of the glutaminase and asparaginase activities: Results of Table 2 show that the glutaminase and aparaginase activities were moderately inhibited (in almost identical manner) in presence of Mg++ and severely inhibited in presence of Hg++. Addition of any of the products of the two reactions, at a concentration five fold that of the substrate, was found to have no inhibitory effect on any of the two activities, a result which is in accordance with what has been suggested from data presented in Fig. 4.
Also similar percentages of inactivation of the two activities were recorded when the pooled fractions were exposed to 50 or 60°C.
It is also worthy to mention that both the extracts and the pooled fractions retained their activities when kept in the freezer for some months without thawing then freezing.
Indication for the presence of a constitutive glutaminase-asparaginase in extracts of a strain of Penicillium chrysogenum and a strain of Penicillium politans.
Extracts of nitrate-grown mats of a strain of Penicillium chrysogenum and a strain of Penicillium politans catalyzed deamidation of L-glutamine and L-asparagine optimally at pH 6-7 and 50°C and at pH 8 and 60-70°C, respectively. Neither acetamide, nicotinamide nor NAD could be deaminated by these two extracts. The ratio of the glutaminase activity to the asparaginase activity was found to be about 1.4 to 1.0 for extracts of the first organism and about 0.8 to 1.0 for extracts of the second organism.
The interesting aspect of the present work is demonstration, for the first time, of the glutaminase-asparaginase activities in extracts of organisms which belong to the filamentous fungi. These organisms are A. niger NRRL3, a strain of Penicillium chrysogenum and a strain of Penicillium politans.
The assumption that the A. niger NRRL3 extracts contain one amidohydrolase having, with comparable efficiency, dual specificity towards both L-glutamine and L-asparagine was based on a number of given criteria which suggest that one enzyme was involved in the hydrolysis of the two amino acids. Each one of these criteria may possibly be non absolutely conclusive by itself, but providing very strong evidence when all taken together; as it is inconceivable that two different amidohydrolases: would be eluted in the same protein fractions, with the same activity peaks and having almost constant L-glutaminase-L-asparaginase ratio in all these fractions, would be affected in almost identical manner on subjection to different factors and would reveal non additive activity in the mixed substrate experiment.
A rather unique feature of the glutaminase-asparaginase of the filamentous fungi tested is their constitutive nature as, the enzyme has been found in mats grown on synthetic medium containing glucose plus mineral salts only. On the other hand, the bacterial enzymes and the suggested yeast enzyme were detected in cells grown in media containing inducers for the enzyme biosynthesis such as glutamine or glutamate and/or yeast extracts.
Constitutivity of the fungal enzyme indicates that it plays an essential role in maintaining the balance between glutamine and glutamate as well as between asparagine and aspartate, thus, regulating the intracellular concentrations of these four amino acids during the dynamics of cell metabolism.
A second distinct feature of the A. niger enzyme resides in the ratio of the glutaminase activity to the asparaginase activity. This ratio was found to be less than one (about 0.7 to 1.0) for the A. niger enzyme, while it was more than one for the previously reported enzymes. Thus, a ratio of : 2.2/1.0, 1.45/1.0, 1.5/1.0, 1.2/1.0 and 2.0/1.0 was reported for the enzyme of: Cryptococcus nodaensis (Sato et al., 1999), Pseudomonas acidovorans (Davidson et al., 1977), Pseudomonas boreopolis 526 (Pekhov et al., 1985), Achromobacter (Roberts et al., 1972) and Pseudomonas 7A (Roberts, 1976), respectively.
The preliminary work carried out with extracts of Penicillium Chrysogenum and Penicillium Politans showed a ratio of about 1.4/1.0 and 0.8/1.0 for the activities of these two extracts, respectively.
The pH activity profiles of the A. niger amidohydrolase with L-glutamine and L-asparagine were almost identical, having their optima at pH 6. similar pH-activity profiles with the two amino acids were also obtained on using extracts of either Penicillium Chrysogenum or Penicillium Politans, however, optimum activities of extracts of the first organism was observed at pH 6-7 while that of the second was at pH 8. A higher pH optimum (pH 9.5) for both activities was reported for the enzyme of Psuedomonas acidovorans (Davidson et al., 1977). On the other hand two different profiles were reported for Psuedomonas 7A enzyme (Roberts, 1976) as well as for Pseudomonas aurantaica 548 enzyme (Lebedeva and Berezov, 1995); thus , the L-glutaminase had optimum activity at pH 6-8 while the L-asparaginase activity increased as the pH increased.
The A. niger glutaminase-asparaginase showed non additive activity on using mixed substrate (L-glutamine+L-asparagine) in the same reaction mixture, a result which suggests that both substrates compete for the same catalytic site of one enzyme. Similar observation was reported for the Acinetobacter enzyme (Steckel et al., 1983).
Recording no product inhibition for the A. niger glutaminase-asparaginase on addition of any of the two products of each reaction to the corresponding reaction mixture is similar to what has been reported for the analogous enzyme of an Achromobacter soil organism (Roberts et al., 1972). In contrast, L-glutamate and L-aspartate showed competitive inhibition with respect to L-glutamine or L-asparagine, as substrate, for the enzyme from Acinetobacter glutaminasificans (Steckel et al., 1983).
The substrate concentration kinetic of the A. niger enzyme exhibited similar hyperbolic rather than sigmoid relationship with each of L-glutamine and L-asparagine. Also proportional enzyme concentration-activity relationship was recorded for each of the two activities. These data indicate that the amidohydrolase has no cooperative binding sites for each of the two amino acids. Similar suggestion was reported for the enzyme from Pseudomonas aurantiaca BKMB-548 (Kabanova et al., 1985).
In conclusion, the present basic study may indicate that the main biochemical properties of the A. niger enzyme could be in favour of its possible utilization, after further extensive studies, as an antitumor therapeutic agent.
Experiments employing variations in culture media are currently in progress and a search for other fungi having this type of enzyme is being continued.
REFERENCES
- Abuchowski, A., T. van Es, N.C. Palczuk, J.R. McCoy and F.F. Davis, 1979. Treatment of L 5178Y tumor-bearing BDF1 mice with a nonimmunogenic L-glutaminase-L-asparaginase. Cancer Treat. Rep., 63: 1127-1132.
PubMedDirect Link - Ammon, H.L., I.T. Weber, A. Wlodawer, R.W. Harrison and G.L. Gilliland et al., 1988. Preliminary crystal structure of Acinetobacter glutaminasificans glutaminase-asparaginase. J. Biol. Chem., 263: 150-156.
Direct Link - Davidson, L., D.R. Brear, P. Wingard, J. Hawkins and G.B. Kitto, 1977. Purification and properties of L-glutaminase-L-asparaginase from Pseudomonas acidovorans. J. Bactiol., 129: 1379-1386.
PubMedDirect Link - Holcenberg, J.S., D.C. Teller, J. Roberts and W.C. Dolowy, 1972. Physical properties of Acinetobacter glutaminase-asparaginase with antitumor activity. J. Biol. Chem., 247: 7750-7758.
PubMedDirect Link - Holcenberg, J.S., B.M. Camitta, L.D. Borella and B.J. Ring, 1979. Phase I study of succinylated Acinetobacter L-glutaminase-L-asparaginase. Cancer Treat. Rep., 63: 1025-1030.
PubMedDirect Link - Huser, A., U. Kloppner and K.H. Rohm, 1999. Cloning, sequence analysis and expression of ans B from Pseudomonas fluorescens, encoding periplasmic glutaminase/asparaginase. FEMS Microbiol. Lett., 178: 327-335.
Direct Link - Jakob, C.G., K. Lewinski, M.W. LaCount, J. Roberts and L. Lebioda, 1997. Ion binding induced closed conformation in Pseudomonas 7A glutaminase-asparaginase (PGA): Crystal structure of the PGA-SO4 (2-)-NH4+ complex at 1.7A resolution. Biochemistry, 36: 923-931.
CrossRefDirect Link - Kien, C.L. and J.S. Holcenberg, 1981. Nitrogen utilization in mice bearing Ehrlich ascites tumor treated with Acinetobacter glutaminase-asparaginase. Cancer Res., 41: 2051-2055.
Direct Link - Lebedeva, Z.I. and T.T. Berezov, 1995. Molecular and catalytic properties of bacterial glutamin-(Asparagin-) ase. Vestn. Ross Akad. Med. Nauk., 2: 57-61.
PubMedDirect Link - Lubkowski, J., A. Wlodawer, H.L. Ammon, T.D. Copeland and A.L. Swain, 1994. Structural characterization of Pseudomonas 7A glutaminase-asparaginase. Biochemistry, 33: 10257-10265.
PubMedDirect Link - Nikolaeve, A.Y., N.N. Sokolov and S.R. Mardashev, 1971. Asparaginase and glutaminase activity of the AG fraction of Pseudomonas fluorescens AG. Biokhimiya, 36: 834-840.
PubMedDirect Link - Oberts, J. and W.G. McGregor, 1991. Inhibition of mouse retroviral disease by bioactive glutaminase-sparaginase. J. Gen. Virol., 72: 299-305.
Direct Link - Ortlund, E., M.W. Lacount, K. Lewinski and L. Lebioda, 2000. Reactions of Pseudomonas 7A glutaminase-asparaginase with diazo analogues of glutamine and asparagine result in unexpected covalent inhibitions and suggests an unusual catalytic triad Thr-Tyr- Glu. Biochemistry, 39: 1199-1204.
PubMedDirect Link - Pekhov A.A., O.S. Zhukova, V.A. Zanin, V.A. Dobrynin and T.T. Berezov, 1984. The effect of preparations of microbial glutamin-(Asparagin)ase on cultured tumor cells. Eksp. Onkol., 6: 55-57.
PubMedDirect Link - Pekhov, A.A., V.A. Zanin, N.N. Magretova and T.T. Berezov, 1985. Isolation, purification and physicochemical properties of glutamin-asparaginase from Pseudomonas boreopolis 526. Biull. Eksp. Biol. Med., 99: 557-560.
PubMedDirect Link - Roberts, J., J.S. Holcenberg and W.C. Dolowy, 1972. Isolation, crystalization and properties of Achromobacteraceae glutaminase-asparaginase with antitumor activity. J. Biol. Chem., 247: 84-90.
PubMedDirect Link - Roberts, J., 1976. Purification and properties of highly potent antitumor glutaminase-asparaginase from Pseudomonas 7A. J. Biol. Chem., 7: 2119-2123.
Direct Link - Sato, I., H. kobayashi, Y. Hanya, K. Abe, S. Murakami, G. Scorzetti and J.W. Fell, 1999. Cryptococcus nodaensis sp. nov, a yeast isolated from soil in Japan that produces a salt-tolerant and thermostable glutaminase. J. Ind. Microbiol. Biotechnol., 22: 127-132.
Direct Link - Steckel, J., J. Roberts, F.S. Philips and T.C. Chou, 1983. Kinetic properties and inhibition of Acinetobacter glutaminase-asparaginase. Biochem. Pharmacol., 32: 971-977.
PubMedDirect Link - Sutherland, E.W., C.R. Cori, R. Haynes and N.S. Olson, 1949. Purification of the hyperglycemic glycogenolytic factor from insulin and gastric mucosa. J. Boil. Chem., 180: 825-834.
PubMedDirect Link