
Samira A. Barghouthi
Department of Chemistry, College of Arts and Sciences,
Arab American University at Jenin, P.O. Box 240, Jenin, Palestine
Journal of Applied Sciences
Year: 2005 | Volume: 5 | Issue: 3 | Page No.: 540-545







ABSTRACT
In this research we present binding studies of some selected antimalarial drugs such as primaquine and quinacrine with biomolecules. Ultraviolet-visible spectrophotometry and fluorescence spectrophotometry along with equilibrium dialysis techniques were used to monitor the interaction of these drugs with myoglobin, hemoglobin albumin and with phospholipid unilamellar vesicles. Fluorescence spectrophotometry, UV-visible difference spectrophotometry and equilibrium dialysis showed no evidence of any binding of these drugs to myoglobin or hemoglobin. However, binding to albumin was evident from a blue shift in UV-Vis absorption spectra. Both primaquine and quinacrine were found to bind to phospholipid vesicles with binding constants of 2.7 ± 0.4x102 and 8.1 ± 0.8x104, respectively. As for the number of molecules per binding site, we found that eight molecules of primaquine occupies ten binding sites and in case of quinacrine two drug molecule for each ten binding sites. Physical parameters for primaquine are determined via UV-Vis absorption at a wavelength of 350 nm. In case of quinacrine fluorescence intensity was employed to measure concentrations with an excitation wavelength of 425 nm and an emission wavelength at 497 nm.
PDF Abstract XML References Citation
How to cite this article
Samira A. Barghouthi, 2005. Thermodynamic Studies of Antimalarial Drugs and Their Interaction with Myoglobin, Hemoglobin and Phospholipid Model Membranes. Journal of Applied Sciences, 5: 540-545.
DOI: 10.3923/jas.2005.540.545
URL: https://scialert.net/abstract/?doi=jas.2005.540.545
DOI: 10.3923/jas.2005.540.545
URL: https://scialert.net/abstract/?doi=jas.2005.540.545
INTRODUCTION
Antimalarial drugs have been shown to bind to RNA, DNA, liposomes and some proteins[1-3]. However, most of these studies have emphasized the clinical aspects[4-6] rather than the molecular interactions between these drugs and proteins. Other researchers[7] have studied the complexes formed by reacting the mustard derivatives of these drugs with proteins; in these cases the drug has basically been used as an extrinsic fluorescence probe. Quinine dihydrochloride, specifically, is known to bind to plasma proteins[8]. However, it is not known whether any of these drugs bind to myoglobin or hemoglobin. As these two proteins are of such importance in living systems this study will answer the question of whether or not selected antimalarial drugs bind to myoglobin and hemoglobin. Binding of these drugs to lipids should affect the time of achieving medicinal effect when used to treat malaria. Binding of these drugs can be used to monitor any protein denaturing effects and possible cytotoxicity.
In order to achieve a better understanding of antimalarial drug protein or membrane interactions one must: (a) establish spectroscopic properties of the target drugs (absorption, emission and fluorescence lifetimes), (b) establish the spectroscopic properties of macromolecule of interest and © compare the properties of the pure substances to those of mixtures of the protein or phospholipid vesicles with antimalarial drug.
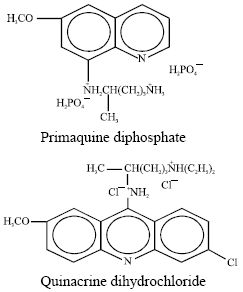 | |
| Fig. 1: | Chemical structures of antimalarial drug |
The structures of primaquine diphosphate and quinacrine dihydrochloride are shown in Fig. 1.
MATERIALS AND METHODS
Materials: Quinine sulfate, quinacrine dihydrochloride and primaquine diphosphate were purchased from Aldrich Chemical Company. Hemoglobin (2x crystallized), albumin, dimyristoyl phosphatidyl choline (DMPC), cholesterol and buffers were purchased from Sigma Chemical Company. Myoglobin (horse) was purchased from Calbiochem. All chemicals and proteins were used as purchased with no further purification.
Experimental conditions: All experiments were performed at the physiological pH of 7.2 using 0.05 M phosphate buffer and a temperature of 37°C; pH 5 acetate buffer was used in the case of quinine sulfate because of low solubility at neutral pH. At pH 7.2 and based on the published pKa values[9] primaquine would be neutral (90%) and quinacrine would be positively charged (the tertiary nitrogen is 99% protonated and the ring nitrogen is 90% protonated). The concentrations were 5xl0-5 M for myoglobin and 2-10 mg mL-1 for hemoglobin. Fluorescence spectra of primaquine diphosphate were monitored using an excitation wavelength of 352 nm and an emission range of 400-650 nm. For quinacrine dihydrochloride, an excitation wavelength of 380 nm was used and the emission spectra were monitored from 400 to 700 nm.
Phospholipid vesicles were prepared using 90% DMPC and 10% cholesterol. Ten milliliter samples were prepared using phosphate buffer and pulsed sonification for 30 min.
Methods: The pKa, values for primaquine diphosphate were determined using both conventional potentiometric titration and spectrophotometry and found to agree with existing literature values[9]. Absorption and emission spectra were obtained at different pH values ranging from 1 to 12 at molar concentrations of 5 mmol for both primaquine and quinacrine using a Cary UV-visible spectrophotometer and a Spex fluoromax.
Fluorescence lifetime measurements were performed using a time correlated single photon counting fluorometer (TCPC) at an excitation wavelength of 295 nm.
The TCPC measurements were made on a similar instrument employing a Coherent laser Nd:YAG as the excitation source. The frequency doubled outputs of both Nd:YAG lasers were used to pump dye lasers (rhodamine 6G) and the outputs of the latter were then frequency doubled to yield an excitation wavelength range from 285 to 315 nm. The excitation pulse width was approximately 5 ps and excitation was conducted under magic angle conditions. Fluorescence emission was selected by use of an American Holographies DB-10 double-monochromator (with zero-time dispersion) and detected with Hamamatsu microchannel plate (model No. R2809U-07 PMT). The instrument response functions (IRF) were measured by use of Ludox AS-40 (colloidal silica) as a Rayleigh scatterer; the IRF values were stable with a full width at half maximum, typically about 40-60 ps. Time correlated single photon counting intensity decay data were analyzed by use of standard non-linear least squares based methodology. The statistical criteria for goodness of fit included assessment of residuals and of the reduced chi-square values such as the two values in Table 1. A global program (Global Unlimited, revision 3, Laboratory for Fluorescence Dynamics, University of Illinois at Urbana-Champaign, Urbana, IL 61801) was used to analyze the intensity decay data.
Once the fluorescence properties of the free drugs were characterized, the interaction of these drugs with myoglobin and with hemoglobin were studied;
| • | Fluorescence spectra of primaquine and quinacrine were obtained as a sample of the drug was titrated by adding aliquats of proteins. Any shifts in the spectra would give an indication of formation of a drug-protein complex. |
| • | Possible changes to the protein, as antimalarial drug is added, were monitored in the UV-visible 250-700 nm range. Difference absorption spectra were obtained by subtracting the absorption spectrum of the pure drug from that of the drug- protein mixture. Three spectra were acquired, free protein, free drug, protein/drug mixture and the effect of the drug on absorption spectra of protein is observed by subtracting spectrum of free drug from that of the protein-drug mixture. |
This procedure was repeated at increasing concentrations of the drug and sets of difference spectra were obtained. These difference spectra were then compared to the experimentally determined spectrum of the protein.
Possibility of these drugs binding to the targeted proteins was further studied using equilibrium dialysis[10] in a sodium phosphate buffer (pH 7.2). Solutions of 0.16-10 mM primaquine and 0.05-5 mM quinacrine were used versus myoglobin or hemoglobin. These studies were repeated at different protein concentrations ranging from 2-10 mg mL-1.
Binding studies with phospholipid vesicles were performed using Unilamellar vesicles of 90% DMPC and 10% cholesterol. These vesicles were prepared using an ultrasound sonifier[10]. Equilibrium dialysis was carried out at varied concentrations of the drug in question. Samples of free and bound drugs were monitored using both UV-visible and fluorescence spectrophotometry. Primaquine concentrations in the free and bound form were measured using UV-vis spectrophotometry at an absorption wavelength of 350 nm (absorptivity coefficient at pH 7.15 is 3.568x103). Quinacrine concentration of free and bound form was monitored using fluorescence spectrophotometry with an excitation wavelength of 425 nm and emission at 497 nm. Concentrations were then calculated using standard curves and baseline corrections as well as dilution factors were applied.
RESULTS AND DISCUSSION
This study answers the question of whether or not selected antimalarial drugs will bind to myoglobin or hemoglobin. Under our experimental conditions, there is no evidence of any binding of either primaquine, quinacrine, or quinine to either myoglobin or hemoglobin. Any difference in spectra is easily explained as a dilution effect. The fluorescence lifetime decay (Fig. 2) of both primaquine diphosphate and quinacrine dihydrochloride show a multiexponential fluorescence decay.
Fluorescence lifetime analysis (Table 1) are shown to exhibit multiexponential decay indicative of a multicomponent system. Both primaquine and quinacrine have long fluorescence lifetimes, 8.626 and 9.012 ns, respectively which correspond to the fraction of the drug in its neutral form and a shorter fluorescence lifetime, 2.589 and 3.152 ns, respectively, which is associated with the protonated fraction of the drug. The third small fraction of short fluorescence lifetime, 433 ps for primaquine and 564 ps for quinacrine, is probably due to the partially protonated primaquine and quinacrine. Since none of the drugs is binding to myoglobin or hemoglobin and due to the complexity of fluorescence lifetime; existence of three fractions in the free form of the drug, measurements were not performed on the protein-drug mixture and lipid-drug complexes.
Fluorescence emission spectra of primaquine or quinacrine in the presence of myoglobin or hemoglobin do not show any detectable shift in the fluorescence at our experiments concentrations of tested antimalarial drugs. While the fluorescence signal for either of the tested drugs in presence of hemoglobin was complicated by the fluorescence signal from hemoglobin, the fluorescence emission spectra for either drug with hemoglobin are unaffected by the presence of any of the tested antimalarial drugs at our experimental conditions.
| Table 1: | Fluorescence lifetimes of primaquine-diphosphate and qumacrine-dihydrochloridea |
| (a) Conditions, pH 7.2 phosphate buffer and 25°C; (b)Fluorescence lifetime; (c)Fractions | |
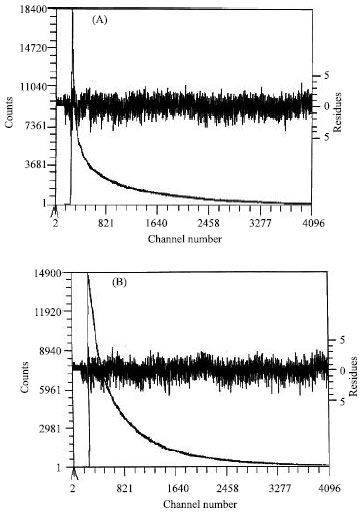 | |
| Fig. 2: | Fluorescence lifetimes of both primaquine diphosphate (A) and quinacrine dihydrochloride (B) in aqueous buffer of pH 7.15 |
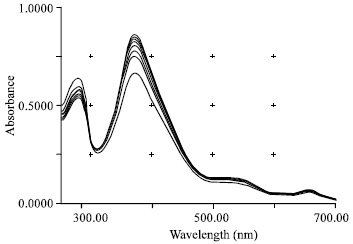 | |
| Fig. 3: | Absorption spectra, absorbance versus wavelength, for myoglobin in the absence (top) and presence of the antimalarial drug primaquine-HCl (20-1,000 μL of 1x10-4M stock solution). Spectra are presented without dilution factor corrections for the sake of visual clarity |
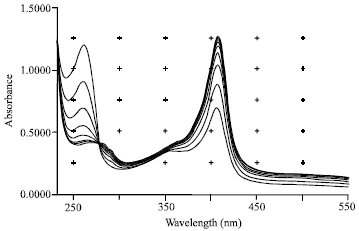 | |
| Fig. 4: | Absorption spectra, absorbance versus wavelength, for hemoglobin (0.25 mg mL-1) in the absence (top) and presence of the antimalarial drug primaquine diphosphate (25-1,000 μL of 1x10-4M stock solution). Spectra are presented without dilution factor corrections for the sake of visual clarity |
 | |
| Fig. 5: | Subtracted absorption spectra, absorbance versus wavelength, for hemoglobin (0.25 mg mL-1) in the absence (top) and presence of the antimalarial drug primaquine diphosphate (25-1,000 μL of 1x10-4M stock solution) |
Inspection of the absorption spectra of myoglobin as primaquine is added (Fig. 3) show that there is no binding of primaquine to myoglobin. Also, no effect is observed on myoglobin when quinine or quinacrine is added. When dilution factors are applied absorption spectra are superimposable. Here we present spectra without the application of dilution factors, only for visual clarity.
Figure 4 show absorption spectra for hemoglobin in the absence and presence of the antimalarial drug primaquine. Also, no effect is seen in case of quinacrine or quinine. Difference spectra show no change in absorption spectrum of hemoglobin as the different drugs are added, thus indicating that none of the above mentioned drugs is binding to hemoglobin.
We obtained absorption spectra of hemoglobin after subtracting absorption signal of the primaquine. It is obvious that although primaquin is present, no effect is observed on the hemoglobin absorption spectra. When dilution factors are applied spectra are superimposable however, we show spectra without correction for dilution only for the sake of visual clarity (Fig. 5).
Silamut et al.[5] had reported binding of quinine to whole plasma proteins and speculated that this type of binding might explain the rarity of quinine cell toxicity in severe falciparum malaria as compared to other quinine derivatives. Other groups studied interaction with enzymes Kristensen et al.[11], lipids Moreau et al.[12] and other molecules[13-18].
We plan to further monitor protein-antimalarial interactions by studying the binding of quinine derivatives to several plasma proteins, especially to the most abundant plasma protein, serum albumin. Our initial spectrophotometric studies of the binding of primaquine and quinacrine show that quinacrine binds to bovine serum albumin as detected from the blue shift in absorption spectra of albumin in the presence of quinacrine. On the other hand absorption spectra of albumin are only slightly affected by the presence of primaquine, or quinine which suggests a non-binding process or a much weaker binding than that of quinacrine.
Binding to phospholipid unilamellar vesicles was determined for primaquine and quinacrine. Binding constants were determined using methods of Barghouthi et al.[10] and via scathchard plot analysis of saturation binding isotherms.
Figure 6A and 7A are saturation binding isotherms obtained by ploting (v), concentration of drug in the bound form/concentration of phospholipid versus concentration of the drug in the free form as obtained from experiments of binding using equilibrium dialysis techniques[10] scathchard plots are obtained by ploting (v)/concentration of the drug in the free form versus v.
Figure 6B and 7B show a negative cooperativity for the binding of promaquine diphosphate and quinacrine dihydrochloride to phospholipid vesicles. Figure 6B is complicated because of the difficulty we experienced in trying to achieve saturation with primaquine.
Results show a larger binding constant for quinacrine and also show that a quinacrine molecule occupies more binding sites. Both of these findings are consistent with the larger size of quinacrine molecule compared to primaquine and with the protonation state of these molecules at the experimental pH of 7.15.
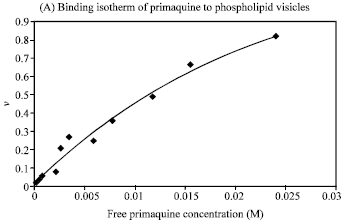 | |
| Fig. 6A: | Binding isotherm for the antimalarial drug primaquine diphosphate to phospholipid vesicles |
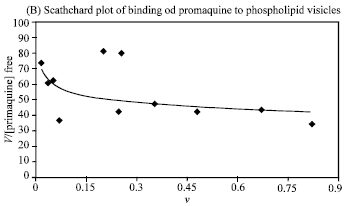 | |
| Fig. 6B: | Binding scathchard plot for the antimalarial drug primaquine diphosphate to phospholipid vesicles |
At this point it is important to state the complications of the experimental procedure as the primaquine concentrations of stock solution could not be prepared to match those of quinacrine, unfortunately that fact minimizes the significance of the difference in binding isotherms (Fig. 6A and 7A). As we mentioned in experimental procedures we were not able to determine binding data for both drugs via the same experimental procedures. Therefore present data and analysis are limited to what we can observe at our experimental conditions and concentrations of the antimalarial drugs.
Quinacrine has a positive charge on the tertiary nitrogen and the ring nitrogen resulting in a stronger electrostatic attraction to the partially negative phospholipid bilayer. These charges will also hinder quinacrine from partitioning to the inner layer of the bimembrane of phospholipid vesicles and results in less accessable binding sites.
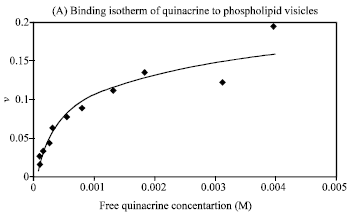 | |
| Fig. 7A: | Binding isotherm for the antimalarial drug quinacrine dihydrochloride to phospholipid vesicles |
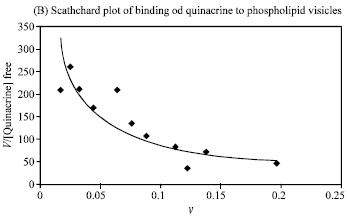 | |
| Fig. 7B: | Binding scathchard plot for the antimalarial drug quinacrine dihydrochloride to phospholipid vesicles |
We think that the unusually large number of primaquine molecules per binding site is a result of both the relatively smaller size of primaquine compared to quinacrine and to the hydrophobicity of the molecule. These two factors result into allowing primaquine molecules crossing to the less polar inner layer of the phospholipid vesicles. This is clear in the binding isotherm for primaquine that exhibits a slightly sigmoidal shape; characteristics of two types of binding sites (Fig. 6A).
Results of such investigations should help in understanding the difference in toxicity of different quinine derivatives and would either complement available clinical studies[18] or shed a new light on the presence of some other effects of these drugs on proteins. The binding of quinines to other macromolecules such as glycoproteins and lipoproteins should also be investigated in an attempt to further understand their toxicity.
ACKNOWLEDGMENTS
We are grateful to the Office of Sponsored Research at Southeastern Louisiana University for their partial support to this project. Many Thanks to Dr. Sedarous at Mayo Clinic for his help with the fluorescence decay. We thank Professor Steinbrecht at University of Rostock, Germany for facilitating the completion of this project. Partial support to this project was made possible, year 2003, by Deutscher Akademischer Austausch Dienst (DAAD); German Academic Exchange Service.
REFERENCES
- Valenzuela, C.F., J.A. Ken and D.A. Johnson, 1992. Quinacrine binds to lipid-protein interface of torpedo acetylcholine receptor: A fluorescence study. J. Biol. Chem., 267: 8238-8244.
Direct Link - Zidovetzki, R., W. Sherman, A. Atiya and H. de Boeck, 1989. A nuclear magnetic resonance study of the interactions of the antimalarials chloroquine, quinacrine, quinine and mefloquine with dipalmitoylphosphatidyl choline bilayers. Mol. Biochem. Parasitol., 35: 199-207.
PubMedDirect Link - Zamora, J., H.L. Pearce and W.T. Beck, 1988. Physical-chemical properties shared by compounds that modulate multidrug resistance in human leukemic cells. Mol. Pharmacol., 33: 454-462.
Direct Link - Jamaludin, A., M. Mohamed, V. Navaratnam, N. Mohamed, E. Yeoh and W. Wernsdorfer, 1988. Single-dose comparative kinetics and bioavailability study of quinine hydrochloride, quinine sulfate and quinine bisulfate sustained-release in healthy male volunteers. Acta leidensia, 57: 39-46.
PubMedDirect Link - Capomacchia, A.C. and S.G. Chulman, 1975. Electronic absorption and fluorescence spectrophotometry of quinacrine. Anal. Chimica Acta, 77: 79-85.
PubMedDirect Link - Kristensen, S., R.H. Wang, H.H.J. Tonnesen, J. Dillon and J. Roberts, 1995. Photoreactivity of biologically active compounds. Viii. Photosensitized polymerization of lens proteins by antimalarial drugs in vitro. Photochem. Photobiol., 61: 124-130.
Direct Link - Moreau, C., S. Jean-Claude, P.F. Devaux and A. Zachowski, 1997. Drug-induced transmembrane lipid scrambling in erythrocytes and in liposomes requires the presence of polyanionic phospholipids. Mol. Membr. Biol., 14: 5-12.
Direct Link - Mustonen, P., Y.A.J. Lehtonen and P.K.J. Kinnunen, 1998. Binding of quinacrine to acidic phospholipids in pancreatic phospholipase A2. Effect on the catalytic activity of the enzyme. Biochemistry, 37: 12051-12057.
CrossRefDirect Link - Arias, H.R., 1997. The high-affinity quinacrine binding site is located at a non-annular lipid domain of the nicotinic acetylcholine receptor. Biochem. Biophys. Acta, 1347: 9-22.
CrossRefDirect Link - Gregory, R.B. and G.J. Barritt, 1996. Store-activated Ca2+ inflow in xenopus laevis oocytes: Inhibition by primaquine and evaluation of the role of membrane fusion. Biochem. J., 319: 755-760.
PubMedDirect Link - Borissova, R., P. Stjaernkvist, M. Karlsson and I. Sjoeholm, 1995. Biodegradable microspheres. 17. Lysosomal degradation of primaquine-peptide spacer arms. J. Pharm. Sci., 84: 256-262.
PubMedDirect Link