
Gunhan Erdem
College of Health, Onsekiz Mart University, �anakkale, Turkey
Sevil Oldacay
College of Health, Onsekiz Mart University, �anakkale, Turkey
Journal of Applied Sciences
Year: 2004 | Volume: 4 | Issue: 2 | Page No.: 277-281







ABSTRACT
The aim of this study was to examine the discriminatory power of random amplified polymorphic DNA (RAPD) markers in sunflowers, and to determine the effect of various physical exposures on two different sunflower genotypes at molecular level. These exposures are different levels of γ-radiation, magnetic fields and a combination of both. The effect of these exposures on sunflower varieties has been determined using the RAPD method. The results of this study showed that γ-radiation and magnetic fields altered the genetic markers of sunflowers.
PDF Abstract XML References Citation
How to cite this article
Gunhan Erdem and Sevil Oldacay, 2004. Employment of RAPD Technique to Assess the Genetic Stability of Helianthus annuus Treated with Different Mutagenic Agents. Journal of Applied Sciences, 4: 277-281.
DOI: 10.3923/jas.2004.277.281
URL: https://scialert.net/abstract/?doi=jas.2004.277.281
DOI: 10.3923/jas.2004.277.281
URL: https://scialert.net/abstract/?doi=jas.2004.277.281
INTRODUCTION
The use of radiation as a mutagen for inducing variation is well established. Although the desired variation is often lacking, radiation can be used to induce mutations and thereby generate genetic variation from which desired mutants may be selected. Like radiation, other physical and chemical mutagens such as X-ray and ethylmetansulphonate (EMS) are used in the area of plant breeding. Magnetic fields (MF) can also be used for plant breeding due to their biological effects[1].
Previous study showed that γ-radiation and magnetic field exposures create the alteration in the chlorophyll content and peroxidase activity in the sunflower genotypes. While the radiation reduced the chlorophyll contents and enhanced the peroxidase activities, magnetic fields had opposite effect when comparing to radiation. Furthermore, magnetic fields had a capacity to reduce the effect of γ-radiation[2].
Different methods are available to investigate the effect of mutagens on plants. Molecular markers allow a direct comparison of the effects on genotypes at the DNA level. A variety of molecular techniques have been developed and are widely used in many fields such as agriculture, biology, medicine etc. for various purposes. The exploration of random amplified polymorphic DNA (RAPD) as genetic markers has improved the effectiveness of recombinant DNA techniques. The RAPD method does not require DNA probes or prior sequence information. The method is simple, requires only small amounts of DNA and can be performed without the use of radioactivity[3]. RAPD fingerprinting is a modification of the polymerase chain reaction (PCR). The method utilises a single, arbitrarily-chosen primer to amplify a number of fragments from a given template of DNA to generate a discrete “fingerprint” when resolved by gel electrophoresis. The assessment of RAPD markers for various purposes has been demonstrated in a number of species such as soybean[4], wheat[5], rice[6], barley[7] and sunflowers[8].
The development of molecular markers which are based on polymorphisms found in proteins or DNA has greatly facilitated research in a variety of disciplines such as taxonomy, phylogeny, ecology, genetics and plant breeding.
In the present study, we treated two sunflower genotypes with radiation, magnetic fields and a combination of both using different doses and then analyzed the effect of these different exposures on the genotypes with RAPDs.
MATERIALS AND METHODS
The two sunflower genotypes used in the study, Nantio and AS 508, are widely grown in Turkey. In this study, the seeds obtained from 300 different generations for each genotype.
Mutagen exposures: For the exposure of γ-radiation on the seeds, 137Cs has been used. According to the testing exposures, GR50 dose that reduces the height of seedlings at the rate of 50% of γ-radiation has been determined 150 gray (Gy) for both genotypes. Therefore the starting dose of γ-radiation exposure has been applied at the level of 100 Gy.
| Table 1: | Sequences of randomly selected 10-mer primers used in the study |
 | |
| Table 2: | The effects of radiation, magnetic fields and the combined exposures on the germination |
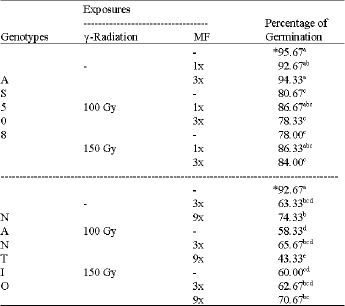 | |
| Control germination values for each genotype are given as * symbol. Values in the same column with different supercript letters are significantly different at P≤0.05 for AS508, at P≤0.01 for Nantio, based on Duncan’s Multiple Range Test | |
300 seeds obtained from each genotype in a polyethylene bag have been incubated at room temperature (RT) in the γ-radiation source for 13 and 19.5 min to apply 100 and 150 Gy doses, respectively. After the exposure, the seeds have been stored at +4°C until the sowing. For the exposure of magnetic fields, maximum MF levels that have not effect on germination rate of each genotype have been selected as the threshold value. AS 508’s seeds have been incubated at RT for 5 sec (1x-threshold value for AS508) and 15 sec (3x); Nantio’s seeds have been incubated at RT for 15 sec (3x-threshold value for Nantio) and 45 sec (9x) in the 5 mT magnetic fields. For the combinated exposures, MF have been exposed after the radiation.
Sowing: Sowing has been performed at greenhouse conditions. All of the seeds have been sowed at the beginning of April while the temperature of soil has been 10°C. Seeds have germinated after approximately 10 days from sowing.
DNA isolation: For each genotype and exposure, 10 seedlings that grow normally have been used for isolating DNA. High molecular weight (approx. 10 kb) plant genomic DNA has been isolated from the fresh leaves of each seedling in the 14th day of growing with reference to a modified method of Doyle and Doyle[9] by using the MBI Fermentas genomic DNA isolation system. The molecular weight and purity of DNA samples have been controlled by agarose gel electrophoresis. The concentration of DNA samples has been spectrophotometrically determined.
RAPD: The 6 different 10-mer oligonucleotides have been used as the primer for the RAPD (Table 1). A 25 μl reaction mixture contained 0.5 μg sunflower genomic DNA, 2.5 μl 10 x Promega PCR buffer, 2.5 mM MgCl2, 25 pmol primer, 200 μM dNTPs and 0.5 unit of Promega Taq polymerase. The conditions for denaturation, annealing and extension for 30 cycles are 94°C for 25 sec, 35°C for 45 sec and 72°C for 1 min, respectively. The RAPD samples have been loaded on 1.5% (w/v) agarose gel and electrophoresed at a constant current of 100 mA for about 3 h in a 0.5xTBE (5 mM tris-borate; 0.1 mM EDTA, pH 8.3) running buffer. After the electrophoresis, the gel has been stained for 10 min in 0.5 μg ml-1 ethidium bromide and examined with 254 nm UV light and photographed on Polaroid 667.
RESULTS
Germination: Number of the seeds sowed for each exposure are 300. Radiation exposures decreased the percentage of germinations in the both of genotypes at statistically significant level (for AS508 p≤0.05; for Nantio p≤0.01) (Table 2). MF exposures also reduced the germination rate at the level of 1x for AS508 and level of 3x and 9x for Nantio. Therefore, the combinated exposures showed that MF also reduced the radiation effect. After the 100 Gy exposure of radiation, MF exposures (1x for AS508 and 3x for Nantio) are effective to reduce the radiation effect, while MF exposure at the all doses that have been applied, reduced the effect of 150 Gy radiation exposure.
RAPD results: RAPD analysis has been performed for all of the DNA samples. RAPD results of controls for each primer are the same in each genotype. However, there are some differences in the exposed patterns. Because of this, we selected one DNA sample for each genotype randomly and performed RAPD analysis on the DNA samples with three repetitions. Repetitive results are the same for each one.
All the exposures examined could be distinguished on the basis of their RAPD profiles.
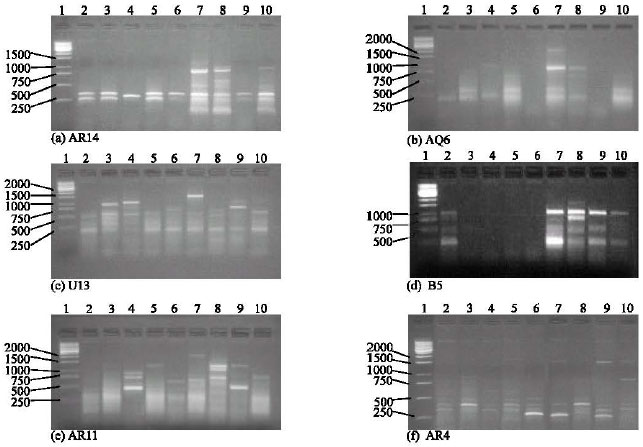 | |
| Fig. 1: | RAPD results of AS 508 genotype with different primers. In all gel images, lane 1 includes 1 kb marker DNA (MBI Fermentas) while other lanes, from 2 to 10, include RAPD samples as follows: lane 2: control, lane 3: 5 sec in 5 mT (1x5 mT), lane 4: 15 sec in 5 mT (3x5 mT), lane 5: 13 min in γ-rad source (for 100 Gy γ-rad), lane 6: 19.5 min in γ-rad source (for 150 Gy γ-rad), lane 7: 1x5 mT and 100 Gy γ-rad, lane 8: 3x5 mT and 100 Gy γ-rad, lane 9: 1x5 mT and 150 Gy γ-rad, lane 10: 3x5 mT and 150 Gy γ-rad. Primers: AR14 in (a); AQ6 in (b); U13 in (c); B5 in (d); AR11 in (e) and AR4 in (f) |
Therefore, the study has aimed at investigating whether RAPD markers can be used as a tool to detect mutant plants or not. We used RAPD markers to study genetic variation in the sunflower varieties that have been irradiated with different gamma doses exposed to different magnetic fields and a combination of both. We are able to obtain different and distinctive fingerprint patterns for all exposures. Polymorphisms has become evident as the presence and/or absence of DNA fragments between the samples. Control RAPD patterns of AS 508 genotype (Fig. 1, lanes 2) have amplified two bands (250 and 350 bases [b]) with primer AR14 (a), three bands (200, 300 and 500 b) with primer U13 (c), four bands (250, 500, 750 and 1000 b) with primer B5 (d) and two bands (300 and 400 b) with primer AR4 (f), while no band has been obtained with primer AQ6 (b) and AR11 (e). Control RAPD patterns of the Nantio genotype (Fig. 2, lanes 2) have amplified two bands (250 and 350 b) with primer AR14 (a), three bands (200, 300 and 1000 b) with primer U13 (c), single band (250 b) with primer B5 (d) and two bands (350 and 400 b) with primer AR4 (f) have been observed, while no band has been obtained with primer AQ6 (b) and AR11 (e) as in AS 508 genotype. The different results obtained from primer U13 and B5 have showed that the RAPD technique is efficient enough to distinguish between the two common genotypes (AS 508 and Nantio) of sunflower.
There are major differences among the RAPD results of exposed samples for each genotype. For AS508 genotype, MF exposure at the level of 1x changed the patterns of RAPD in comparison with controls (Fig. 1, lanes 4). Gamma radiation exposure has also changed the RAPD patterns in AS508 genotype, especially at the level of 150 Gy dose (Fig. 1, lanes 6). All of the combinated exposures is effective the alteration in RAPD patterns (Fig. 1, lanes 7 to 10). Same results have been obtained for Nantio genotype with the exposures of MF alone (at the level of 3x and 9x), γ-radiation alone (100 and 150 Gy doses) and combinated exposure (Fig. 2). As a result, the genetic stability is not high at all among the both genotypes of sunflower exposed either to 100-150 Gy γ-radiation or 5 mT MF or combined exposures.
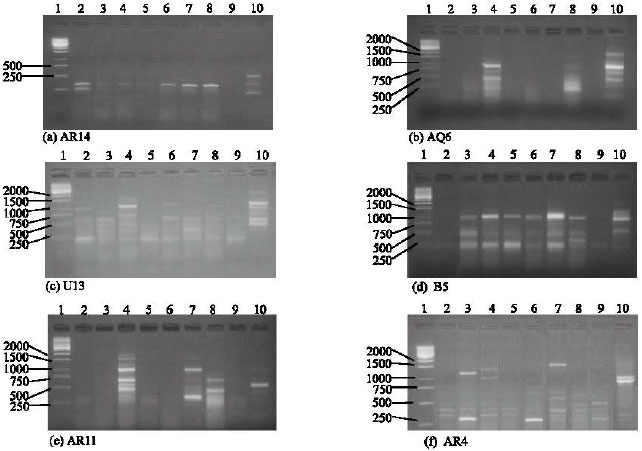 | |
| Fig. 2: | RAPD results of Nantio genotype with different primers. In all gel images, lane 1 includes 1 kb marker DNA (MBI Fermentas) while other lanes, from 2 to 10, include RAPD samples as follows: lane 2: control, lane 3: 15 sec in 5 mT (3x5 mT), lane 4: 45 sec in 5 mT (9x5 mT), lane 5: 13 min in γ-rad source (for 100 Gy γ-rad), lane 6: 19.5 min in γ-rad source (for 150 Gy γ-rad), lane 7: 3x 5 mT and 100 Gy γ-rad, lane 8: 9x5 mT and 100 Gy γ-rad, lane 9: 3x5 mT and 150 Gy γ-rad, lane 10: 9x5 mT and 150 Gy γ-rad. Primers: AR14 in (a); AQ6 in (b); U13 in (c); B5 in (d); AR11 in (e) and AR4 in (f) |
DISCUSSION
The RAPD patterns of the samples exposed to γ-radiation, magnetic fields and the combination of both are different from the controls of the two genotypes. This finding indicates that γ-radiation and the magnetic fields created polymorphic regions in the sunflower genome. This result has been expected for γ-radiation[10]. The similar effect of the magnetic fields on the sunflower genomes is original and unexpected. The magnetic fields have also created polymorphisms and altered the effect of γ-radiation.
Radiation is one of the best known physical mutagens. It dissociates the atoms of water molecule and causes the generation of hydroxyl radicals that are the most reactive[11]. They react with most of the biomolecules include DNA and scavenge electrons from them. The oxidation of biomolecules by the radicals damages their structure and biological activity. By the way, genetic alterations occur on the DNA molecules. This is the cause of mutations depend on radiation.
There are a lot of biological effects include carcinogenesis attribute MF, but there is no clear data about the mutagenic effect of MF[12-14].
In general, the biological effects of MF depend on the alterations of electron spins of molecules, especially ionic forms. Some studies have suggested that MF exposure could be due to both the increase in the concentration[15] and oscillating of free radicals[16]. MF are known to affect radical pair recombination and they may increase the concentration of oxygen free radicals in living cells[15]. Increasing the concentration of free radicals creates oxidative stress, enhances stress response and some biological reactions, such as DNA damage occurs under this condition[17].
The reason of the increase in the oscillating of free radicals is the transformation of the radicals into more stabile and less reactive forms under the MF energy[16].
In the present study, it is observed that MF exposure of the sunflower seeds has altered the effect of radiation. MF energy can change the reactivity of the radicals generated by radiation. Therefore, the patterns of combined exposures for both genetic variations have differed from the alone exposures of MF and radiation for them.
In conclusion, the results obtained from the present study suggest that magnetic fields as a new physical mutagen could be used alone or with the combination of radiation for mutant selection in plant breeding.
ACKNOWLEDGMENT
This research has been supported by Çanakkale Onsekiz Mart University Research Fund (project no: 2001/17).
REFERENCES
- Matsuda, T., H. Asou, M. Kobayashi and M. Yonekura, 1993. Influences of magnetic fields on growth and fruit production of strawberry. Acta Hortic., 348: 378-380.
Direct Link - Oldacay, S. and G. Erdem, 2002. Evaluation of chlorophyll contents and peroxidase activities in Helianthus annuus genotypes exposed to radiation and magnetic fields. J. Applied Sci., 2: 934-937.
CrossRefDirect Link - Karp, A., O. Seberg and M. Buiatti, 1996. Molecular techniques in the assessment of botanical diversity. Ann. Bot., 78: 143-149.
Direct Link - Zhang, J.H., M.B. McDonald and P.M. Sweeney, 1996. Soybean cultivar identification using RAPD. Seed Sci. Technol., 24: 589-592.
Direct Link - Gang, D.R. and D.J. Weber, 1996. Using random amplified polymorphic DNA to analyze the genetic relationships and variability among three species of wheat smut (Tilletia). Bot. Bull. Acad Sinica, 37: 173-180.
Direct Link - Mezencev, N., A. Ghesquiere, P. Marmey, M.C. Combes and E. Guiderdoni, 1997. Assessment of RAPD markers to detect genetic change in protoplast-derived rice plants. J. Genet. Breed., 51: 97-102.
Direct Link - Svitashev, S., T. Bryngelsson, X.M. Li and R.R.C. Wang, 1998. Genome-specific repetitive DNA and RAPD markers for genome identification in elymus and hordelymus. Genome, 41: 120-128.
PubMed - Iida, S., M. Kusaba and T. Nishio, 1997. Mutants lacking glutelin subunits in rice: Mapping and combination of mutated glutelin genes. Theor. Applied Genet., 94: 177-183.
Direct Link - Leibovitz, B. and B.V. Siegel, 1980. Aspects of free radical reactions in biological systems: Aging. J. Gerontol., 35: 45-56.
PubMed - Schreiber, W.G., E.M. Teichmann, I. Schiffer, J. Hast and W. Akbari et al., 2001. Lack of mutagenic and co-mutagenic effects of magnetic fields during magnetic resonance imaging. J. Magnetic Resonance Imag., 14: 779-788.
PubMed - Novikov, V.V., I.M. Sheiman and E.E. Fesenko, 2002. Effect of weak and superweak magnetic fields on the intensity of asexual propagation of Planarians Dugesia tigrina. Biofizika, 47: 125-129.
PubMed - Jajte, J.M., 2000. Programmed cell death as a biological function of electromagnetic fields at a frequency of (50/60 Hz). Medycyna Pracy, 51: 383-389.
PubMed - Gutzeit, H.O., 2001. Biological effects of ELF-EMF enhanced stress response: New insights and new questions. Electro Magnetobiol., 20: 15-26.
Direct Link