
H. Watanabe
Faculty of Agriculture, Niigata University, Ikarashi, Niigata, 950-2181, Japan
Y. Adachi
Faculty of Agriculture, Niigata University, Ikarashi, Niigata, 950-2181, Japan
M. Saigusa
Research Center for Advanced Agrotechnology and Biotechnology, Toyohashi University of Technology, Toyohashi, Aichi, 441-8580, Japan







ABSTRACT
Improving early seedling growth is crucial for direct-seeding rice cultivation. In this study, field and laboratory experiments were conducted to determine the effects of Plant Growth Regulators (PGRs) such as gibberellic acid (GA3) and ethephon (ET) on the early growth of direct-seeded rice. Seeds were treated with the PGRs and pre-germinated at 30°C in the dark. For the field experiment, two Flooding Depth (FD) regimes were established during the growth period: 0 cm (drained condition) and 2 cm (flooded condition). Treatment of seeds with ET or GA3 alone promoted the growth of seedling organs compared to that of the control at both the FDs in some cases. However, combined application of ET and GA3 (ET+GA3) synergistically improved the growth of different seedling organs at both the FDs in most cases with slight exception: the coleoptile and mesocotyl plus coleoptile (mesocotyl+coleoptile) length were not different across the PGR treatments under drained condition. Further, the growth of the second node tiller was stimulated by the application of ET alone under drained conditions. The laboratory experiment revealed that ET+GA3 increased the growth rate of mesocotyl+coleoptile and that of the first leaf. These results suggest that the combined application of ET and GA3 might be useful for improving the growth performance of rice seedlings in the direct seeding method.
PDF Abstract XML References Citation
Received: May 07, 2015;
Accepted: June 10, 2015;
Published: June 29, 2015
How to cite this article
H. Watanabe, Y. Adachi and M. Saigusa, 2015. Synergistic Effects of Ethephon and Gibberellic Acid on the Growth of Rice Seedlings Grown under Field and Environmentally Controlled Conditions. Journal of Agronomy, 14: 87-92.
DOI: 10.3923/ja.2015.87.92
URL: https://scialert.net/abstract/?doi=ja.2015.87.92
DOI: 10.3923/ja.2015.87.92
URL: https://scialert.net/abstract/?doi=ja.2015.87.92
INTRODUCTION
Direct seeding involves planting of seeds without prepare for seedbed and raising seedlings. This method is highly economical than transplanting system, since it reduces labor. For direct seeding of rice, ensuring early seedling growth, including seedling emergence and vigor, is crucial. Treatment of seeds with Plant Growth Regulators (PGRs) is known to improve early seedling growth. Plant growth regulators such as gibberellin, ethylene and abscisic acid are known to promote the elongation of rice seedling organs (Suge, 1974; Raskin and Kende, 1983; Takahashi and Kaufman, 1983, 1992; Furukawa et al., 1997; Watanabe and Takahashi, 1997; Watanabe et al., 2001). Suge (1974) indicated that combined application of ethylene and GA1, a naturally active gibberellin in rice plants, synergistically stimulated the growth of coleoptiles and the first leaf in rice. Furukawa et al. (1997) suggested that the synergistic actions of ethylene and gibberellins on leaf elongation were attributed to the increasing responsiveness to and turnover of GA1 by ethylene application. Previously, we showed that the combined application of ethephon (ET) and gibberellic acid (GA3) improved the early growth of rice cultivated at different growth temperatures and flooding depths (Watanabe et al., 2007). However, our previous study and other relevant studies on PGRs (Suge, 1974; Furukawa et al., 1997) were conducted in growth chambers; therefore, analyzing the effects of PGRs in field trials is necessary for their practical application agronomically. Elongation of seedling organs, including mesocotyl, coleoptile and the first leaf, play important roles in the better emergence and establishment of seedlings obtained using the direct seeding method (Watanabe et al., 2006, 2007; Ogiwara and Terashima, 2001; Mori et al., 2012); however, investigations on the growth kinetics of these organs treated with ET and GA3 are still lacking.
This study aimed to investigate the effects of single and combined application of ET and GA3 on the early growth of seedling organs of rice cultivated at different flooding depths under field conditions. Further, the growth-promoting effects of ET and GA3 on rice seedlings were assessed under environmentally controlled conditions.
MATERIALS AND METHODS
General procedure: A rice (Oryza sativa L.) Japonica cultivar Hitomebore was used in the study. The seeds were treated with PGRs as described previously (Watanabe et al., 2007). Briefly, the seeds were sterilized with thiuram and benomyl solution for 24 h according to instructions, immersed in water for 24 h and then soaked in the test solution for 48 h. Ethephon (2-chloroethylphosphonic acid; Ishihara Sangyo Kaisha, Ltd., Osaka, Japan) was used as an ethylene-producing compound (Arteca, 1996) and GA3 (Sigma Chemical Co., MO, USA) was used for gibberellin. The components of the test solution were as follows: (1) water (control), (2) 50 mg L–1 ethephon (ET), (3) 100 mg L–1 GA3 and (4) 50 mg L–1 ET+100 mg L–1 GA3 (ET+GA3). After the seeds were treated with the PGRs, they were immersed in water for 48 h to remove any excess test solution. Seed sterilization and treatments were performed at 15°C. The imbibed seeds were germinated in water maintained at 30°C in the dark.
Field experiments: In 2004, a field trial was conducted at the Field Science Center of Tohoku University, Miyagi Prefecture, Japan (38°44’N, 140°15’E; altitude, 220 m). This area has non-allophanic volcanic ash soil (Andisol and Alic Pachic Mellanudand). Nitrogen, potassium and phosphorus were applied as chemical fertilizers at the rates of 8 g m–2. The fertilizers were applied to the paddy, 7 days before seeding. Metal flames (30 l×30 w×15 h cm) were buried in the soil and then the paddy within the flame was divided into 4 parts for the PGR treatments by using thin plastic plates. The size of each treatment plot was 0.0225 m–2 (15×15 cm). On May 11, 25 uniformly germinated seeds were sown in each treatment plot at a depth of 1.0 cm. After seeding, two flooding depth regimes were established throughout the growth period: 0 cm (drained conditions) and 2 cm (flooded conditions). The seedlings were grown for 30 days.
After the seedlings were sampled, the plant length and organ lengths were measured. The numbers of tillers on the second node (the second node tiller) were counted and their lengths were measured using a ruler. The emergence of tillers suggests that their tips had emerged from the subtending leaf (second leaf) sheath of the mother stem. The length of the tillers included the leaf blade and leaf sheath.
Laboratory experiment: The rice cultivar, method of PGR treatments and preparation of seeds were the same as described above. The laboratory experiment was performed as described previously (Watanabe et al., 2007) unless otherwise stated. The PGR-treated and germinated seeds were sown at a seeding depth of 1.0 cm in seedling pots with small compartments containing fertilized granulated soils (Kureha Chemical Co., Ltd., Tokyo, Japan). Twenty one uniformly germinated seeds were used for each treatment. The seeds were grown at 20°C with a 12 h light and dark photoperiod. Fluorescent lamps were used to supply photosynthetic photon flux of 120 μ mol m–2 sec–1. The flooding depth was 2 cm. The seedlings were sampled at 22 Days After Seeding (DAS) and the plant length and the length of each organ were measured.
Experimental design and statistical analysis: A randomized complete block design with four replicates was used. Data is presented as Mean±Standard Error (SE) and were analyzed using analysis of variance (ANOVA). Pairwise multiple comparison was performed using Tukey test at the 5% significance level. Statistical analysis was performed using JMP for Windows, version 4.0. (SAS Institute, Cary, NC, USA).
RESULTS AND DISCUSSION
Field experiment|Air temperature condition: The air temperature during the experimental period is shown in Fig. 1. The temperature was higher than the average temperature in May during sowing and continued to increase up to the seedling emergence stage. Thereafter, the temperature gradually began to reduce and was only slightly higher than the average temperature until the end of June.
Stimulating effects of ET and GA3 on shoot elongation: The PGR treatments significantly affected the morphological parameters of seedlings grown under both drained and flooded conditions (Fig. 2). Under drained conditions, treatment with ET or GA3 alone, did not affect the growth of seedling organs such as the mesocotyl, coleoptile, mesocotyl+coleoptile and the first and second leaves (Fig. 2a).
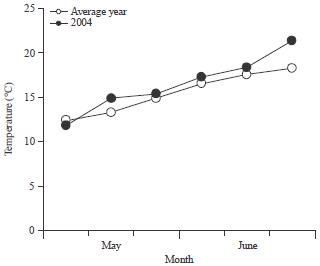 | |
| Fig. 1: | Air temperature conditions during the experimental period |
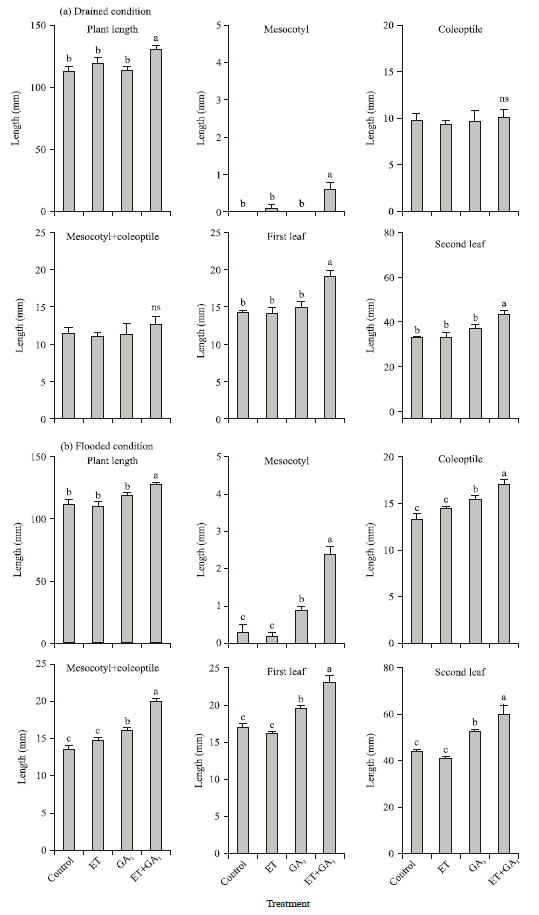 | |
| Fig. 2(a-b): | Comparison of growth parameters of control and plant growth regulator-treated seedlings grown under different flooding regimes in the field, (a) Drained condition and (b) Flooded condition. Means±SE (n = 4) for each treatment are shown. Different letters indicate significant differences by Tukey test at the 5% significance level, ns: Not significant. Comparisons valid within the same flooding regimes |
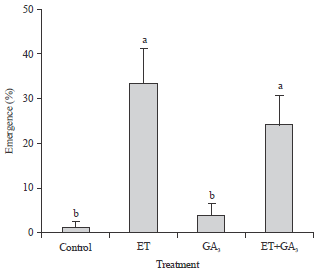 | |
| Fig. 3: | Effects of plant growth regulators on the percent emergence of second node tillers under drained conditions. Means±SE (n = 4) for each treatment are shown. Different letters indicate significant differences by Tukey test at the 5% significance level |
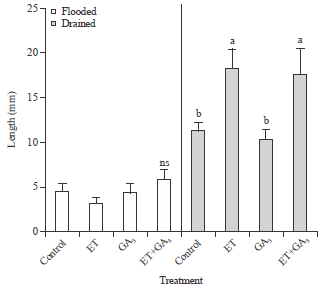 | |
| Fig. 4: | Effects of plant growth regulators on the length of the second node tillers. Means±SE (n = 4) for each treatment are shown. Different letters indicate significant differences by Tukey test at the 5% significance level, ns: Not significant. Comparisons valid within the same flooding regimes |
However, ET+GA3 treatment significantly elongated the mesocotyl and improved the growth of the first and second leaves and plant length. Under flooded conditions, combined treatment with ET and GA3 produced the most prominent effect on seedling growth. However, the responses of seedling organs to each PGR treatment under flooded condition were slightly different from those of seedlings grown under drained conditions. Under flooded condition, GA3 alone also significantly improved the growth of mesocotyl, coleoptiles, mesocotyl+coleoptile and the first and second leaves compared to those in the control (Fig. 2b); these effects were slightly improved after treatment with ET+GA3. Yamauchi and Biswas (1997) also suggested that the growth of seedling organs differed between flooded and drained conditions.
Thus, even under field conditions, the synergistic effects of ET and GA3 on seedlings growth were noted; however, coleoptile and mesocotyl+coleoptile growth under drained conditions were slightly different from those obtained in the previously reported growth chamber experiments (Watanabe et al., 2007).
Ethephon affected the growth of the second node tiller: Tiller number can significantly contribute to the yield potential; however, information on the hormonal regulation of tillering is scarce (Hanada, 1993; Kariali and Mohapatra, 2007). Hence, the effects of PGR on tiller number were determined using the different flooding regimes. Since, the second node tillers were not noted under flooded conditions, the percent emergence of tillers under drained conditions was recorded (Fig. 3). The ET and ET+GA3 treatments significantly increased the percent emergence of second tillers than those noted in control and GA treatment alone (Fig. 3). The lengths of the tillers were not significantly different across the PGR treatments under flooded conditions; however, ET and ET+GA3 treatments significantly increased the tiller length under drained condition compared to those noted in control and GA3 treatments under drained conditions (Fig. 4). Further, under drained condition the tiller length was not different after treatment with ET and ET+GA (Fig. 4). These results suggested that ET promoted tillering but not synergistically with GA under drained conditions.
Tillering is known to be affected by environmental factors such as nutrient supply, plant spacing and flooding depth (Hanada, 1993; Yoshida, 1981); it is adversely affected with increasing water depth in paddy fields (Hanada, 1993). In this study, the second node tiller number was not increased after treatment with ET and ET+GA3 under flooded conditions; this suggested that the growth-promoting effect of ET on tillering was eliminated by flooding. This finding was consistent with that of Sekiya (1951), who indicated that the growth of the second tillering primordium and tillering bud were remarkably influenced by water depth and that their growth was the best at a water depth of 0 cm.
ET has been used to increase tillering and prevent lodging in many cereal crops, including maize, wheat and barley (Cox and Andrade, 1988; Cox and Otis, 1989; Foster et al., 1991). However, the growth-promoting effect of ET on tillering in rice has not yet been investigated. To our knowledge, this is the first report suggesting that ET promotes tiller growth in rice. Further studies are warranted to elucidate the mechanisms underlying the effect of ET on tiller growth in rice.
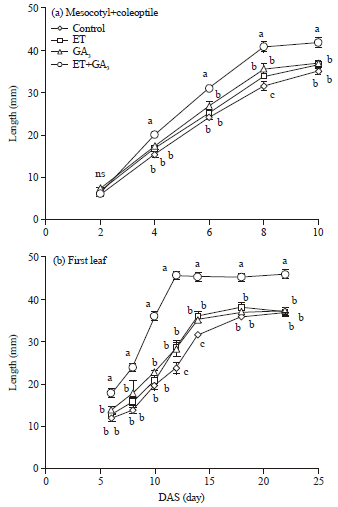 | |
| Fig. 5(a-b): | Differences in the growth of organs between the plant growth regulators-treated and control plants, (a) Mesocotyl+coleoptile and (b) First leaf. Means±SE (n = 4) for each treatment are shown. The seedlings were grown in a growth chamber at 20°C. Different letters indicate significant differences by Tukey test at the 5% significance level, ns: Not significant |
Laboratory experiments: The length of coleoptile plus mesocotyl and the first leaf are important factors that determine successful emergence and establishment of seedlings (Andrews et al., 1997; Hoshikawa, 1993; Ries and Hofmann, 1995; Alibu et al., 2011). Since, the final length of seedling organs were different across the PGR treatments (Fig. 2) and ET+GA3 improved the emergence and establishment of seedlings in the field experiments, the growth kinetics of seedling organs treated with PGRs were analyzed (Fig. 5).
The mesocotyl+coleoptile length was not different between the control and treated seedlings at 2 DAS; however, their length significantly increased after treatment with PGRs from 4 DAS. ET+GA3 had the most significant effect on mesocotyl+coleoptile lengths from 4-10 DAS. The final length of mesocotyl+coleoptile after treatment with ET+GA3, increased approximately by 18% compared with the other treatments. Further, the elongation rates of mesocotyl+coleoptile were the highest after treatment with ET+GA3 (Fig. 5a).
The synergistic effects of ET and GA3 were also noted for the growth of the first leaf (Fig. 5b). ET+GA3 significantly increased the first leaf length compared to those of the other treatments at 6 DAS. The growth of the first leaf after treatment with ET+GA3 was noted only until 12 DAS. In other treatments, the first leaf continued to grow even after 14 DAS. The final length of the first leaf after treatment with ET+GA3 was increased around by 24% than those of the other treatments. Nonetheless, the highest growth rate of the first leaf, as well as that of the coleoptile+mesocotyl and the longest first leaf length were noted after treatment with ET+GA3. These results suggested that the elongation rate and speed of mesocotyl+coleoptile and first leaf in ET+GA3 contribute to early seeding growth and vigor.
CONCLUSION
Our results showed the synergistic effects of ET and GA3 on the growth of rice seedlings in the field experiment. In the future, we intend to perform experiments to determine the effects of ET+GA3 on the later growth stages of rice seedlings, as well as on the grain yield and rice quality. To our knowledge, this is the first study showing that ET increased the growth of the second node tiller under drained conditions. The findings of this study might form the basis to determine the mechanism of how ET promotes tiller growth and facilitate the use of ET in rice production.
REFERENCES
- Alibu, S., Y. Saito, H. Shiwachi and K. Irie, 2011. Relationship between coleoptile and mesocotyl elongation of upland rice (Oryza sativa L.) seedlings under submergence and soil-sand culture. Afr. J. Agric. Res., 6: 6463-6472.
Direct Link - Andrews, M., A. Douglas, A.V. Jones, C.E. Milburn, D. Porter and B.A. McKenzie, 1997. Emergence of temperate pasture grases from different sowing depths: Importance of seed weight, coleoptiele plus mesocotyl length and shoot strength. Ann. Applied Biol., 130: 549-560.
CrossRefDirect Link - Cox, W.J. and H.F. Andrade, 1988. Growth, yield and yield components of maize as influenced by ethephon. Crop Sci., 28: 536-542.
CrossRefDirect Link - Cox, W.J. and D.J. Otis, 1989. Growth and yield of winter wheat as influenced by chlormequat chloride and ethephon. Agron. J., 81: 264-270.
CrossRef - Foster, K.R., D.M. Reid and J.S. Taylor, 1991. Tillering and yield responses to ethephon in three barley cultivars. Crop Sci., 31: 130-135.
CrossRef - Furukawa, K., Y.Y. Yang, I. Honda, T. Yanagisawa, A. Sakurai, N. Takahashi and Y. Kamiya, 1997. Effects of ethylene and gibberellins on the elongation of rice seedlings (Oryza sativa L.). Biosci. Biotech. Biochem., 61: 864-869.
CrossRefDirect Link - Kariali, E. and P.B. Mohapatra, 2007. Hormonal regulation of tiller dynamics in differentially-tillering rice cultivars. Plant Growth Regul., 53: 215-223.
CrossRefDirect Link - Mori, S., H. Fujimoto, S. Watanabe, G. Ishioka, A. Okabe, M. Kamei and M. Yamauchi, 2012. Physiological performance of iron-coated primed rice seeds under submerged conditions and the stimulation of coleoptile elongation in primed rice seeds under anoxia. Soil Sci. Plant Nutr., 58: 469-478.
CrossRefDirect Link - Ogiwara, H. and K. Terashima, 2001. A varietal difference in coleoptile growth is correlated with seedling establishment of direct Seeded rice in submerged field under low-temperature conditions. Plant Prod. Sci., 4: 166-172.
CrossRef - Raskin, I. and H. Kende, 1983. Regulation of growth in rice seedlings. J. Plant Growth Regul., 2: 193-203.
CrossRefDirect Link - Ries, R.E. and L. Hofmann, 1995. Grass seedling morphology when planted at different depths. J. Range Manage., 48: 218-223.
CrossRefDirect Link - Sekiya, F., 1951. Studies on the tillering primordium and tillering bud in rice seedlings: 1. The influence of water depth on the development of tillering primordium and tillering bud. Jpn. J. Crop Sci., 20: 63-66.
CrossRef - Watanabe, H. and K. Takahashi, 1997. Effects of plant growth regulators on the appearance of MC type rice seedlings. Jpn. J. Crop Sci., 66: 318-324.
CrossRef - Watanabe, H., K. Takahashi and M. Saigusa, 2001. Morphological and anatomical effects of abscisic acid (ABA) and fluridone (FLU) on the growth of rice mesocotyls. Plant Growth Regul., 34: 273-275.
CrossRefDirect Link - Watanabe, H., M. Saigusa and S. Morita, 2006. Identification of Casparian bands in the mesocotyl and lower internodes of rice (Oryza sativa L.) seedlings using fluorescence microscopy. Plant Prod. Sci., 9: 390-394.
CrossRef - Watanabe, H., S. Hase and M. Saigusa, 2007. Effects of the combined application of ethephon and gibberellin on growth of rice (Oryza sativa L.) seedlings. Plant Prod. Sci., 10: 468-472.
CrossRef - Yamauchi, M. and J.K. Biswas, 1997. Rice cultivar difference in seedling establishment in flooded soil. Plant and Soil, 189: 145-153.
CrossRefDirect Link