
Morteza Kamrani
Department of Plant Production, Moghan College of Agriculture and Natural Resource, University of Mohaghegh Ardabili, Ardabil, Iran
Asghar Ebadi
Department of Plant Production, Moghan College of Agriculture and Natural Resource, University of Mohaghegh Ardabili, Ardabil, Iran
Manoochehr Shiri
Department of Plant Production, Moghan College of Agriculture and Natural Resource, University of Mohaghegh Ardabili, Ardabil, Iran
Journal of Agronomy
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 227-233







ABSTRACT
The objective of the present study is to develop an efficient protocol for plant regeneration and Agrobacterium-mediated transformation of potato cultivars Desiree, Agria and Marfona grown under Iranian agricultural conditions. The regeneration efficiency from internode, leaf and petiole explants was examined. The results from all Shoot Induction Medium (SIM) combinations and all cultivars together indicate highly efficient shoot regeneration from internode (4.56 shoots per explant). The response of petiole and leaf explants was lower, 3.83 and 2.55 shoots per explant, respectively. The highest efficiency of shoot regeneration was achieved with internode explants of cultivar Desiree (6.64 shoots per explant) on Murashige and Skoog (MS) medium supplemented with thidiazuron (TDZ) 1 mg L–1 + 6-benzyladenine (BAP) 1 mg L–1. For plant transformation, internode explants were inoculated and co-cultivated with Agrobacterium tumefaciens strain LBA4404 harboring a binary vector pBI-121 containing β-glucuronidase (GUS) and nptII genes. Reverse transcriptase-polymerase chain reaction analysis and histochemical assay for β-glucuronidase indicated that the gene coding for this enzyme was integrated in the potato genome and could be expressed in potato tissues. The presence of nptII gene in the kanamycin resistant plants was verified by polymerase chain reaction analysis. The transformation frequency ranged from 22-42%.
PDF Abstract XML References Citation
Received: July 18, 2015;
Accepted: August 28, 2015;
Published: September 30, 2015
How to cite this article
Morteza Kamrani, Asghar Ebadi and Manoochehr Shiri, 2015. Effect of Explant, Genotype and Plant Growth Regulators on Regeneration and Agrobacterium-Mediated Transformation of Potato. Journal of Agronomy, 14: 227-233.
DOI: 10.3923/ja.2015.227.233
URL: https://scialert.net/abstract/?doi=ja.2015.227.233
DOI: 10.3923/ja.2015.227.233
URL: https://scialert.net/abstract/?doi=ja.2015.227.233
INTRODUCTION
Potato (Solanum tuberosum) is one of the most economically important annual vegetable crops of Solanaceae family. It is the fourth most cultivated food crop in the world after wheat, rice, maize and is the most important dicotyledonous tuber crop. The edible part of plant is tuber used as cheap food, industrial raw material, animal feed and seed tuber. Potato is sensitive to a number of biotic and abiotic factors. Several approaches including traditional breeding techniques have been made in the past to overcome these constraints. But traditional methods are considered to be complicated in nature and consumed comparatively a longer period to produce a variety with desired characters. To address some of these limitations, plant genetic engineering using desirable genes from various organisms has provided promising results. Most of the transformation studies were conducted with the intention to transfer genes/traits expected to increase the resistance of potato against predators and pathogens (Chye et al., 2005) or to modify common metabolic pathways, such as starch and sucrose synthesis (Edwards et al., 1999).
Genetic engineering of potato cultivars requires the development of transformation protocols and the establishment of efficient regeneration systems. Various important factors such as genotype, source of explants and plant growth regulators significantly influence regeneration of potato. Among the factors that influence growth, the type of cytokinin, such as thidiazuron (TDZ) and 6-benzyladenine (BAP), is critical for initiation of shoot primordia from cultured explants. TDZ, a phenylurea-type cytokinin, has been used as a plant growth regulator to induce organogenesis, including adventitious regeneration of transformed plants in many plant species that were thought to be recalcitrant to regeneration (Cuenca et al., 2000; Corredoira et al., 2008; Sriskandarajah and Lundquist, 2009).
Transformation efficacy in potato is actually highly genotype-dependent, which is the main reason for the existence of many different protocols. The objective of this study is to establish an effective protocol for plant regeneration and transformation of local Iranian varieties, using different types of explants that would allow genetic engineering for important traits. Three cultivars, Agria, Marfona and Desiree were used for transformation with an Agrobacterium binary vector (pBIl21) which included genes for GUS and nptII.
MATERIALS AND METHODS
Plant materials and culture conditions: In vitro grown plants of potato cultivars Agria, Marfona and Desiree prepared from Genetic Engineering Laboratory, University of Tabriz, Tabriz, Iran. These cultivars were propagated using single-node segments on MS medium (Murashige and Skoog, 1962) containing 2% w/v sucrose and 0.8% w/v agar. The pH of the medium was adjusted to 5.8 before autoclaving and the cultures were grown under a 16 h day–1 photoperiod with a light intensity of 56 μmol m‾2 sec–1 provided by cool white fluorescent tubes and incubated at 22±2°C ambient temperature.
Regeneration experiments: Internode, petiole and leaf segments of in vitro grown potato cultivars were cut into sections (1 cm each) or small pieces (1 cm2) as explants for callus induction. The explants were cultured on callus induction medium [MS agar medium containing 2% sucrose supplemented with 2 mg L–1 6-benzyladenine (BAP) and 1 mg L–1α-naphthalene acetic (NAA)] and incubated at 22±2°C under a light intensity of 56 μmol m‾2 sec–1. The calli were transferred to the fresh callus induction medium about 21 days interval for further proliferation and maintenance. After 2-3 weeks of incubation, the well-developed compact calli were cultured in Shoot Induction Media (SIM) containing MS basal medium supplemented with various combinations of BAP (1 and 2 mg L–1), kinetin (1 and 2 mg L–1) and thidiazuron (1 and 2 mg L–1). Approximately 40-45 explants were cultured on each of the 8 types of SIM combination. Each experiment was repeated three times. The cultures were incubated in a growth chamber under same conditions of light and temperature, as that of callus induction. After 4-6 weeks, shoot formation was observed. The shoots were excised and transferred to root induction medium (MS medium containing 0.1 mg L–1NAA) for root induction.
Bacterial strains: Agrobacterium tumefaciens strain LBA4404 possessing binary vector pBI121 was inoculated on YEB medium and used for gene transfer into potato.
Agrobacterium tumefaciens-mediated transformation: Internodal (5-6 mm) explants were prepared from 4 weeks old plants for transformation. The internodes were wounded several times leaving about a one mm gap between the cuts. The internode pieces were placed in MS liquid medium containing 1 mg L–1NAA, 2 mg L–1BAP and 2% sucrose at pH 5.7, in Petri dishes for 48 h at room temperature for co-cultivation. After the dark incubation period, internode pieces were washed three times in sterile water and in a cefotaxime solution (250 mg L–1) to remove bacterial excess. The explants were blotted dry with a sterile filter paper and then placed in callus induction medium (MS medium containing 1 mg L–1 NAA, 2 mg L–1 BAP, 50 mg L–1 kanamycin, 250 mg L–1cefotaxime, 3% sucrose and 0.8% agar at pH 5.7). After 7 days of incubation, the explants were transferred to the shoot induction medium (MS medium, supplemented with 1 mg L–1 TDZ+2 mg L–1 BAP, 75 mg L–1 kanamycin, 250 mg L–1cefotaxime, 3% sucrose at pH 5.7 solidified with 0.8% agar). The explants were transferred every week to fresh shoot induction medium. After four weeks, shoots formed at the cut end of the internodes. Regenerated shoots of 2.0-2.5 cm in length were then excised and transferred into rooting medium (MS solidified medium supplemented with 0.1 mg L–1NAA, 2% sucrose, 100 mg L–1cefotaxime and 75 mg L–1kanamycin).
β-glucuronidase (GUS) histochemical assay: The β-glucuronidase assay was carried out essentially as described by Jefferson et al. (1987). Leaves from kanamycin resistant shoots were tested for histochemical GUS expression in X-gluc solution consisting of 2 mM X-gluc, 0.1 M phosphate buffer (pH 7.0) and 5 mM each of potassium ferricyanide and ferrocyanide with 0.1% (v/v) Triton X-100. After incubating overnight in the substrate solution at 37°C, the stained tissues were rinsed several times with 70% ethanol to remove chlorophyll.
Polymerase Chain Reaction (PCR) analysis: Genomic DNA was isolated from 100 mg of young leaves of transformed and untransformed potatoes by the method of Dellaporta et al. (1983). The PCR amplification was carried out using the gene specific primers (5’-GAACAAGATGGATTGCACGC-3’) and (5’-GAAGAACTCGTCAAGAAGGC-3’) for nptII generating 786 bp product. PCR was performed in a 25 μL reaction mixture containing 50 ng of genomic DNA as template, 1xTaq DNA polymerase buffer, 400 μM each dNTPs, 10 pmol of each oligonucleotide primer and 0.3 U of Taq DNA polymerase (ASTEC PC-818; Fukuka, Japan). DNA amplifications were done in a thermal cycler (Takara, Japan) using the following program: Initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 45 sec, annealing at 55°C for 45 sec and extension at 72°C for 2 min followed by final extension for 5 min at 72°C. The PCR products were separated by electrophoresis in a 1% agarose gel containing ethidium bromide and visualized under UV using Gel Documentation System OptiGo 600/650 (ISOGEN).
Reverse Transcriptase-PCR (RT-PCR): Total RNA was isolated from the transgenic as well as from untransformed control plants using TRIZOL reagent (Invitrogen, USA). Five milligram of total RNA was taken in a nuclease free tube. One microliter of oligo(dT)20 primer (50 μM), 1 μL of annealing buffer and DEPC treated water were added to it so that the final volume was made up to 8 μL. This mixture was incubated at 65°C for 5 min and then slowly cooled to room temperature so that the oligo(dT)s bind to polyA tail. About 10 μL of 2X First-Strand Reaction Mix (Invitrogen, USA) and 2 μL of SuperScriptR III/RNaseOUT™ Enzyme Mix were added. The enzyme mixture was gently mixed and incubated at 50°C for 1 h. The enzyme was deactivated by heating the reaction mixture at 85°C for 5 min. The cDNA (1 μL) synthesized from the transgenic and untransformed control lines were taken as template for PCR using the GUS gene specific primers (5’-CCCTTACGCTGAAGAGATGC-3’ and 5’-GAGCGTCGCAGAACATTACA-3’) with same PCR condition. Actin was used as a loading control with PCR primers described by Nicot et al. (2005). Reaction products were separated on 1% agarose gel stained with ethidium bromide and visualized under UV light.
Statistical analysis: The data pertaining to shoot regeneration was analyzed using two-way analysis of variance to examine the main and interaction effects of cultivar and SIM combination. The differences among means were compared by Least Significant Difference (LSD) at p<0.05 significance level. Analysis was performed using programs in MSTAT-C (Michigan State University, Ann Arbor, MI).
RESULTS
Plant regeneration: It is important to optimize the regeneration frequency of potato to increase the likelihood of recovery of transformants; therefore, different combinations of BAP with TDZ and Kin were evaluated for their regeneration ability of the leaf, internode and petiole explants of Marfona, Agria and Desiree cultivars. Figure 1 shows different developmental stages of regeneration of potato. Callus formation was observed from all types of explants after they were cultured on callus induction medium (Fig. 1a).
 | |
| Fig. 1(a-d): | In vitro plant regeneration of potato, (a) Callus formation on the internodal explants cultured on CIM medium for 4 weeks, (b) Shoot formation from the explants cultured on SIM medium, (c) Rooting of regenerated shoots on MS medium and (d) Hardening of rooted plants in the green house |
After 2 weeks, calli were transferred to Shoot Induction Media (SIM). Shoots were regenerated from nodular callus of all type of explants (Fig. 1b). The evaluation of shoot induction was based on the mean number of shoots per explant. Adventitious roots could be seen forming within 1 week after the regenerated shoots were placed on root induction medium (Fig. 1c). Finally, rooted plants were transferred to pots and grown under greenhouse conditions (Fig. 1d).
Table 1 gives the results of two-way ANOVA for the differences between SIM combinations and between cultivars with regard to number of shoots per explant from particular explants. All null hypotheses regarding the effect of SIM combination, cultivar and explants type were rejected at the 0.01 probability level. The SIM combination×cultivar interaction was significant for all explant sources, indicating that the cultivars differed in the various SIM combinations.
Averaging the results from all SIM combinations and all cultivars together indicates highly efficient shoot regeneration from internode (4.59 shoots per explant). Shoot regeneration from petiole and leaf explants were 3.83 and 2.55 shoots per explant, respectively (Table 2). The highest efficiency of shoot regeneration was achieved on MS medium supplemented with1 mg L–1 BAP+1 mg L–1 TDZ and 2 mg L–1 BAP+1 mg L–1 TDZ for all cultivars, respectively (Table 2).
Plant transformation: Among the different explants tried, internode explants were found to be best for shoot induction. For transformation, MS medium supplemented with 2 mg L–1BAP and 1 mg L–1 NAA (callus induction medium) followed by MS medium supplemented with 2 mg L–1BAP and 1 mg L–1 TDZ for subsequent shoot proliferation was used routinely. A concentration of 75 mg L–1 kanamycin was chosen for selection of transformants. The same parameters were used routinely for transformation studies in all the three varieties. A total of 150 internode explants were cocultivated in the three varieties (Table 3).
| Table 1: | Analysis of variance for effect of growth regulators on regeneration from Solanum tuberosum L. explant sources |
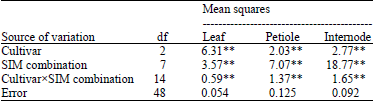 | |
| **Significant at the 0.01 probability level, df: Degree of freedom | |
| Table 2: | Effect of different plant growth regulator combinations, explants and cultivar on plant regeneration |
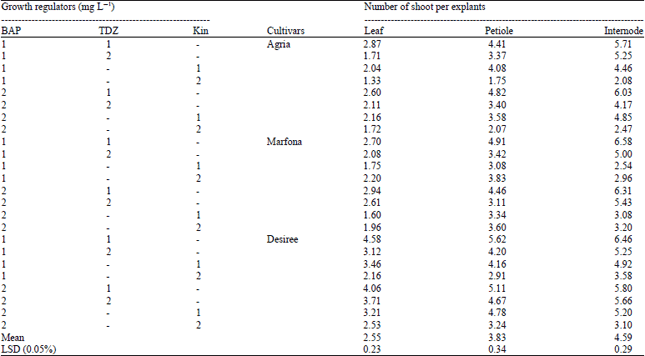 | |
| LSD: Least significant difference, BAP: 6-benzyladenine, TDZ: Thidiazuron, Kin: Kinetin | |
| Table 3: | Efficiency of potato internode transformation mediated by Agrobacterium tumefaciens |
 | |
The highest number of kanamycin resistant shoots was recorded with Desiree (122) followed by Marfona (108) and Agria (86). Although, the explants remained green, some of the shoots did not grow more than 10-15 mm. Only those shoots measuring 2.0-2.5 cm were transferred to rooting medium (MS medium supplemented with 0.1 mg L–1 NAA and 75 mg L–1 kanamycin). Similarly, highest transformation efficiency were recorded in Desiree (42%), followed by Marfona (30%) and lowest in Agria (22%) (Table 3). All rooted plants were planted in pots and molecular analysis was carried out to confirm the transgenic nature.
Confirmation of transgenic plants: In comparison with untransformed wild-type plants which did not contain any expression of GUS, the transgenic plant lines showed the typical dark-blue after the GUS assay. Upon histochemical staining of leaves from kanamycin resistant rooted plants, all lines showed expression of GUS gene (Fig. 2). Besides the GUS assay, the expression of GUS gene was also carried out by RT-PCR analysis. The presence of a band of 362 bp was amplified from the cDNA products of transformed plants while the cDNA products of untransformed plants (control) cDNA showed no amplification (Fig. 3).
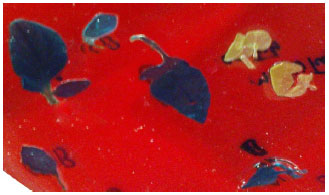 | |
| Fig. 2: | GUS histochemical staining in leaves of transgenic and nontransformed potato plants |
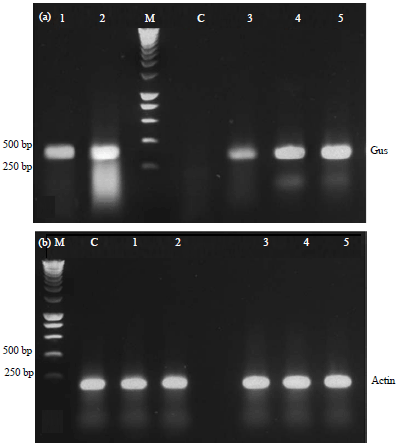 | |
| Fig. 3(a-b): | Reverse transcription (RT)-PCR analysis of transgenic potato lines. RT-PCR amplification of the 362 and 101 bp fragments corresponding to (a) GUS gene and (b) Actin as an internal control, respectively. Lanes 1-5: Transgenic lines, Lane C: Nontransformed plant (control), Lane: M,1 kb BLUE DNA Marker |
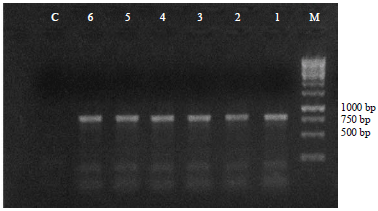 | |
| Fig. 4: | PCR analysis of putative transgenic potato lines for the nptII gene. Lanes 1-6: Transgenic lines, Lane C: Nontransformed plant (control), Lane M:1 kb BLUE DNA Marker |
The presence of the transgenes was further confirmed by PCR analysis using total genomic DNA isolated from leaves of transformed and untransformed (control) plants using specific primers designed to amplify nptII gene. The GUS-positive lines were used for PCR analysis with one non-transgenic line as negative control. Fragments of 786 bp (Fig. 4) were amplified from genomic DNA of all the transgenic plants, whereas, corresponding band was not detected in the untransformed control plant. Rooting on selective (kanamycin) medium was found to be a good indicator of transformation, as 100% of rooted shoots were positive when tested by PCR.
DISCUSSION
The objective of the present study is to develop an efficient Agrobacterium tumefaciens-mediated transformation protocol for important Iranian potato cultivars Agria and Marfona. For this purpose, two separate experiments, plant regeneration and transformation were carried out.
In plant regeneration experiment, we studied the effects of various concentrations and combinations of plant growth regulators on plant regeneration from different explant sources (leaf, petiole and internode) of potato cultivars. First, all explants were placed on callus induction medium (1 mg L–1 NAA and 2 mg L–1BAP). Calli were obtained from all three types of explant of the three investigated cultivars. The BAP and NAA combination have been shown to be an efficient combination for induction of callus in potato (Beaujean et al., 1998). Medium combination and explant source had an important influence on plant regeneration. In the present study, BAP in combination with Kin induced a lower plant regeneration but it promoted the highest plant regeneration in combination with TDZ (4.86 shoot per explant). The highest efficiency of shoot regeneration was obtained in internode explants of cultivar Marfona on MS medium supplemented with 1 mg L–1 TDZ+1 mg L–1 BAP (Table 2). Abd Elaleem et al. (2009) and Khalafalla et al. (2010) reported that MS medium containing 5 mg L–1 TDZ was the best for days to shoot initiation, the highest percentage of callus with shoot and highest number of shoot per callus in potato.
Deore and Johnson (2008) suggested that TDZ and BAP together had a synergistic effect in adventitious shoot-bud induction in Jatrophacurcas. Also they reported that TDZ promoted higher regeneration from leaf explants than Kinetin. Huetteman and Preece (1993) reported that TDZ, a synthetic phenylurea, is considered to be one of the most active cytokinins for shoot induction in plant tissue culture. Husain et al. (2007) suggested that TDZ induces shoot regeneration better than other cytokinins. Apart from cytokinin-like activity, TDZ has been suggested to be a modulator of the endogenous auxin levels. There is experimental evidence that TDZ stimulates de novo synthesis of auxins by increasing the levels of IAA and its precursor, tryptophan (Murthy et al., 1995). Increases in endogenous auxin, cytokinin and ethylene have been seen in response to TDZ treatment (Murthy et al., 1995). Also, TDZ has been shown to be useful for rapid plant regeneration in several recalcitrant species through organogenesis (Malik and Saxena, 1992). Similarly, in our studies, it was observed that TDZ is essential for the high-frequency induction of adventitious shoots from all kind of explants.
Transformation efficiency of 22-42% was observed and true transformed shoots were rooted within 1 week in selection medium. This efficiency is high, as can be seen when comparing this internode transformation protocol with direct selection in kanamycin to other potato transformation protocols that employ leaf segments as explants (Barrell et al., 2002; Soto et al., 2007). Soto et al. (2007) reported higher transformation efficiency (68%) in potato cultivar Desiree using internodal stems as explant and phosphinothricin as a selection agent. While De Block (1988) obtained a transformation efficiency of only 30% with the cultivars Bintje and Desiree using phosphinothricin as selection marker. Moreover, the leaf explants were easily injured during the manipulation, which resulted in a low percentage of transformation (De Block, 1988) while the internodal explants are much more resistant during manipulation and more amenable to in vitro conditions. It was also observed that the transformed cells were located essentially at the section and in the cambium/pericycle cell (Higgins et al., 1992). Longitudinal sectioning of internodes considerably increased the section, with the vascular bundle zone being fully accessible to Agrobacterium and this allowed a large number of transgenic buds to be obtained (Beaujean et al., 1998). Analysis of all putative transgenic lines using PCR established the presence of the nptII gene in all regenerated lines (Fig. 4), thereby confirming their transgenic status and a high rate of success for the Agrobacterium-mediated gene transfer system of potato using kanamycin resistance as a selectable marker (Barrell et al., 2002).
Based on the data of the present study, it was concluded that the reported regeneration system is repeatable and can be easily used to regenerate transgenic potato plants expressing the genes present in the Agrobacterium binary vector T-DNA. Using this regeneration and transformation protocol, transgenic potato with higher contents of pharmaceutical compounds or modified secondary metabolic profiles will be produced in the future by approaches of specific metabolic engineering.
ACKNOWLEDGMENT
Dr. B. Baghban kohnehrouz (University of Tabriz, Tabriz, Iran) is greatly acknowledged for providing potato Agria, Marfona and Desiree cultivars.
REFERENCES
- Abd Elaleem, K.G., R.S. Modawi and M.M. Khalafalla, 2009. Effect of plant growth regulators on Callus induction and plant regeneration in tuber segment culture of potato (Solanum tuberosum L.) cultivar Diamant. Afr. J. Biotechnol., 8: 2529-2534.
Direct Link - Barrell, P.J., S. Yongjin, P.A. Cooper and A.J. Conner, 2002. Alternative selectable markers for potato transformation using minimal T-DNA vectors. Plant Cell Tissue Organ Cult., 70: 61-68.
CrossRefDirect Link - Beaujean, A., R.S. Sangwan, A. Lecardonnel and B.S. Sangwan-Norreel, 1998. Agrobacterium-mediated transformation of three economically important potato cultivars using sliced internodal explants: An efficient protocol of transformation. J. Exp. Bot., 49: 1589-1595.
CrossRefDirect Link - Corredoira, E., A. Ballester and A.M. Vieitez, 2008. Thidiazuron-induced high-frequency plant regeneration from leaf explants of Paulownia tomentosa mature trees. Plant Cell Tissue Organ Cult., 95: 197-208.
CrossRefDirect Link - Cuenca, B., A. Ballester and A.M. Vieitez, 2000. In vitro adventitious bud regeneration from internode segments of beech. Plant Cell Tissue Organ Cult., 60: 213-220.
CrossRefDirect Link - De Block, M., 1988. Genotype-independent leaf disc transformation of potato (Solanum tuberosum) using Agrobacterium tumefaciens. Theoret. Applied Genet., 76: 767-774.
CrossRefDirect Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Deore, A.C. and T.S. Johnson, 2008. High frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: An important biodiesel crop. Plant Biotech. Rep., 2: 7-11.
Direct Link - Edwards, A., D.C. Fulton, C.M. Hylton, S.A. Jobling and M. Gidley et al., 1999. A combined reduction in activity of starch synthases II and III of potato has novel effects on the starch of tubers. Plant J., 17: 251-261.
CrossRefDirect Link - Higgins, E.S., J.S. Hulme and R. Shields, 1992. Early events in transformation of potato by Agrobacterium tumefaciens. Plant Sci., 82: 109-118.
CrossRefDirect Link - Huetteman, C.A. and J.E. Preece, 1993. Thidiazuron: A potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult., 33: 105-119.
CrossRefDirect Link - Husain, M.K., M. Anis and A. Shahzad, 2007. In vitro propagation of Indian Kino (Pterocarpus marsupium Roxb.) using Thidiazuron. In vitro Cell. Dev. Biol. Plant., 43: 59-64.
CrossRefDirect Link - Jefferson, R.A., T.A. Kavanagh and M.W. Bevan, 1987. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J., 6: 3901-3907.
Direct Link - Khalafalla, M.M., K.G.A. Elaleem and R.S. Modawi, 2010. Callus formation and organogenesis of potato (Solanum tuberosum L.) cultivar Almera. J. Phytol., 2: 40-46.
Direct Link - Malik, K.A. and P.K. Saxena, 1992. Thidiazuron induces high-frequency shoot regeneration in intact seedlings of pea (Pisum sativum), chickpea (Cicer arietinum) and lentil (Lens culinaris). Aust. J. Plant Physiol., 19: 731-740.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nicot, N., J.F. Hausman, L. Hoffmann and D. Evers, 2005. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot., 56: 2907-2914.
CrossRefDirect Link - Sriskandarajah, S. and P.O. Lundquist, 2009. High frequency shoot organogenesis and somatic embryogenesis in juvenile and adult tissues of seabuckthorn (Hippophae rhamnoides L.). Plant Cell Tissue Organ Culture, 99: 259-268.
CrossRefDirect Link - Soto, N., G.A. Enriquez, A. Ferreira, M. Corrada, A. Fuentes, K. Tiel and M. Pujol, 2007. Efficient transformation of potato stems segments from cultivar Desiree using phosphinothricin as selection marker. Biotecnologia Aplicada, 24: 139-144.
Direct Link - Chye, M.L., K.J. Zhao, Z.M. He, S. Ramalingam and K.L. Fung, 2005. An agglutinating chitinase with two chitin-binding domains confers fungal protection in transgenic potato. Planta, 220: 717-730.
CrossRefPubMedDirect Link