
Homa Mahmoodzadeh
Science and Research Center of Tehran, Iran Department of Biology, Science Faculty, Islamic Azad University of Mashad, Iran
Ahmad Majd
Department of Biology, Tarbiat Moallem University, Tehran, Iran
Mehrdad Lahouti
Department of Biology, Ferdowsi University, Mashad, Iran
Journal of Agronomy
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 366-369







ABSTRACT
In the present research structure and ultrastructure of shoot apical meristem of canola (Brassica napus cv. Symbol) under salinity conditions were investigated. The experiments were conducted in five groups (0, 3, 6, 9, 12 dS m-1) under greenhouse conditions. Sampling of apical meristem and TEM tissue preparation procedure were carried out. Semithin and ultrathin sections were prepared and viewed in light and electron microscopy, respectively. The results included reduction of meristem size, disorders in meristem structure. Also formation of autophagic vacuoles was observed that is probably one of the plant responses to salt stress for more water storage in these vacuoles and decreasing of cell water requirements.
PDF Abstract XML References
How to cite this article
Homa Mahmoodzadeh, Ahmad Majd and Mehrdad Lahouti, 2007. Effect of Salinity Stress on Structure and Ultrastructure of Shoot Apical Meristem of Canola (Brassica napus cv. Symbol). Journal of Agronomy, 6: 366-369.
DOI: 10.3923/ja.2007.366.369
URL: https://scialert.net/abstract/?doi=ja.2007.366.369
DOI: 10.3923/ja.2007.366.369
URL: https://scialert.net/abstract/?doi=ja.2007.366.369
INTRODUCTION
A heavy environmental concentration of salts unleashes various types of physical and chemical stress in plants, provoking complex responses that involve changes in plant morphology, physiology and metabolism (Cheesman, 1988; Sanders and Harper, 1999). It is commonly accepted that growth inhibition by salt stress is associated with alterations in the water relationships within the plant, caused by osmotic effects with specific ionic consequences (excesses or deficits) or energy availability related to carbohydrate concentrations (Lazof and Bernstein, 1998). Last investigations about meristems of growth inhibition and salt tolerance, often based on comparative studies between salt-tolerant and salt-sensitive cultivars, however, since most of the environmental stresses, influence tolerance level of various developmental processes, salt stress is considered a complex phenomenae (Sanders and Harper, 1999).
A high concentration of NaCl greatly reduces of both the shoot and root (Staples and Harper, 1984; Cheesman, 1988). Among the shoot cells, meristematic cells are especially interesting because mitotic activity and cell division indispausable for shoot growth and because meristematic cells are considered to be one of the salt-sensitive cells (Huang and Steveninek, 1988). Canola is a genetically altered form of rapeseed with low erucic acid, a 22-carbon chain fatty acid that is used in a variety of polymer and lubricant products. Interest in canola is increasing steadily among consumers due to its lowest content of saturated fatty acids (<70 g kg-1) among major oil seeds. Salt-tolerance of Brassicaceae family (including canola)is placed between Poaceae (Poa species) with the most tolerance and Fabaceae (Vicia faba) with the lowest tolerance. Ashraf et al. (2001) showed canola and tomato cv. Monkey maker continue to yielding under NaCl 200 mM. Stepphun and Volkman (2001) compared the effect of different salinity levels on seed emergency, germination rate, plant height, biomass and yield in canola, pea, bean and wheat. The results showed that pea and bean no yield but canola and wheat yield 0.4 and 21% of yielding at non-salinity conditions. In studding of primary growth response of Arabidopsis to salt stress, the important of cell cycle regulation was indicated. Growth decline of stressed roots was due to reducing in cell production and smaller mature cells (West, 2004). Sacks and Silk (1997) showed that salt stress resulted in short-term changes in cell cycle duration, however, replaced with a long-term response that meristem size is reduced but cell cycle duration turn back to initial state.
The objective of this study was to determine the structural and ultrastructural changes of shoot apical meristem of canola (Brassica napus cv. Symbol).
MATERIALS AND METHODS
The experiment was conducted in Faculty of Science, Islamic Azad University of Mashad, Iran (2004).
The seeds of canola (cv. Symbol) were supplied by the Agricultural Research Center of Khorasan, Iran. The study was conducted in greenhouse condition. There were a control group which received no solution of NaCl plus four other groups which treated with salt solutions of 3, 6, 9 and 12 dS m-1. Shoot apical meristems of control and treated plants at vegetative phase were collected and for TEM studies, specimens were fixed in 1% glutaraldehyde with 0. 1 M cacodilate buffer, pH = 7. 2, at 4 for 6 h; post-fixed in 1% osmium tetroxide at 4 for 6 h, then dehydrated in an acetone series, and embedded in spur resin. Semithin sections (0. 5) were prepared using ultramicrotome and stained with toloidine-blue and viewed in light microscopy. After primary distinguishing, ultrathin sections (60-80 A) were prepared and placed on 200 mes grids. Tissue sections were stained with 5% uranyle acetate in methanol for 20 min and observed and photographed by a LEO 914 AB TEM.
RESULTS
Shoot apical meristem of canola is small and cone-shape. tunica consists of two cellular layers and corpus is recognizable. Initial Ring (IR) of the meristem includes active cells with intense chromophilly and medulary meristem is also recognizable. Leaf primary base, Leaf primordium and leaf primary pattern is formed due to mitosis divisions of initial ring. Prepared semithin sections using ultramicrotome, after staining with toloidine_blue and observing with light microscopy, showed structure of shoot apical meristem in details (Fig. 1 and 2). In treated specimens, the shoot apical meristam was underwent disorder and the meristem size was declined (Fig. 3).
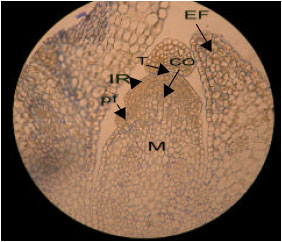 | |
| Fig. 1: | Semithin section of shoot apical meristem of canola (control plant). Toloidine-blue staining (x 20). The cells of central part of shoot meristem are vacuolated and converted to medulary meristem (M), otherwise the cells of tunica and corpus have dense cytoplasm. T: Tunica, CO: Corpus, M: Medulary meristem, IR: Initial Ring, pf: Leaf primordium and EF: Leaf primary pattern |
TEM studies on shoot apical meristam of canola and areas adjacent to the meristem showed that meristematic cells in control plants (Fig. 4) are small and every cell has thin cell wall, nucleus, bulky nucleolus and mitochondria are recognizable. Cytoplasm is densed and osmiophyll due to abundance of ground ribosomes. A part of cytoplasm is occupied with vacuole. In treated plants (12 dS m-1) cell wall is thickened and suitable with cell demands for more relations, a considerable numbers of plasmodesmata have formed in the cell walls (Fig. 5). The important note is that a major part of cytoplasm is occupied with bulky vacuole and many of destructing organells are observed in this autophagic vacuole.
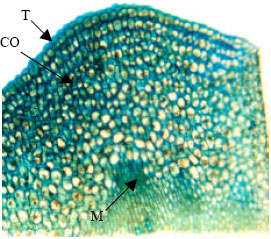 | |
| Fig. 2: | Semithin section of shoot apical meristem of canola (control plant). Toloidine-blue staining (x 40). Two cellular layers of tunica are regular. Underlying the tunica, corpus cells with bulky vacuoles and less chromophilly are recognizable. T: Tunica, CO: Corpus and M: Medulary meristem |
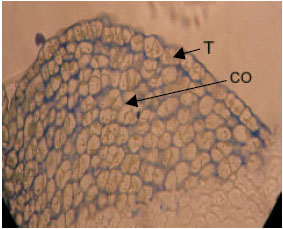 | |
| Fig. 3: | Semithin section of shoot apical meristem of canola (12 dS m-1 treatment). Toloidine-blue staining (x 40). Disorders in tunica organization are observable. T: Tunica and CO: Corpus |
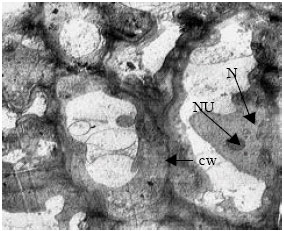 | |
| Fig. 4: | TEM micrograph of shoot apical meristem of control plant (EC = 0). The meristematic cells contain recognizable nucleus and nucleolus, thin cell wall and dense cytoplasm (due to abundance of ribosomes). N: Nucleus, NU: Nucleolus and cw: cell wall |
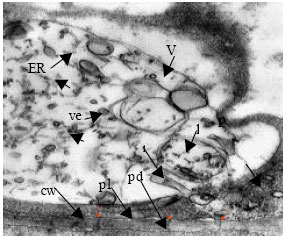 | |
| Fig. 5: | TEM micrograph of shoot apical meristem of stressed plant (12 dS m-1 treatment). One of the meristematic cells contain relatively thick wall with abundant plasmodesmata, plasmalemma, cytoplasm with ribosomes and endoplasmic reticulum. A bulky autophagic vacuole which surrounded most of organells and destroyed them. Cw: cell wall, pl: plasmalemma, pd: plasmodesmata, m: mitochondrium, p: plastid, ve: vesicle, ER: Endoplasmic Reticulum |
Traces of mitochondria, plastids, endoplasmic reticulum and cytoplasmic vesicles are viewed in this vacuole. In the medulary meristam cells, in addition of more vacuolizing of cytoplasm, differentiation of some organells including convert of proplastids to chloroplasts and formation of thylakoids and grana is observed (Fig. 6).
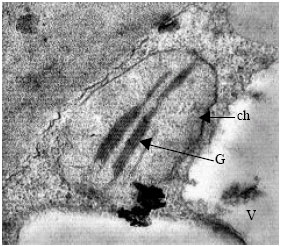 | |
| Fig. 6: | TEM micrograph of shoot apical meristem. One of the medulary meristem cells : Extension of vacuolar system and formation of chloroplast with internal grana is observed. The cytoplasm is granular and contain abundance ribosomes. ch: chloroplast, G: grana, V: Vacuole |
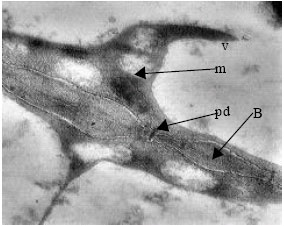 | |
| Fig. 7: | TEM micrograph. One of the cells which is differentiating to protophloem in adjacent to meristematic zone of shoot apical. Plasmodesmata and mitochondria are recognizable. Thick cell wall with number of plasmodesmata that are differentiating to sieve pores. Thickened zones of cell wall (bars) are observed. m: mitochondrium, V: vacuole, pd: plasmodesm, B: Bar |
In areas adjacent to the shoot apical meristem, one of the differentiations that rapidly occurred, is formation of protophloem (Fig. 7). In these cells, remarkable extension of vacuolar system and destruction of a major of cellular contents can be observed. Moreover the intermediate wall including vast plasmodesmata which are differentiating to sieve pores. In the distance among them, the cell wall has thickened and formed the bars. A number of mitochondria in the cells are observed.
DISCUSSION
The results of our studies suggest that salt stress induced structural and ultrastructural changes in the shoot apical meristem of canola. Reduction of meristam size, disorders in the structure of shoot apical meristem, increasing of cell vacuolation and formation of autophagic vacuoles were all responses to salinity condition.
Similar to our results, previous investigations on the effects of salinity on apical meristem structure (Jbir et al., 2003; Hanif and Davies, 1998) showed that the meristem size which is almost the same in the two wheat species (salt-tolrant and salt-sensitive), is decreased under salt stress especially in the salt-sensitive species. Sacks and Silk (1997) showed that water stress cause meristematic cells to be longer and reduced the rates of cell division that result in reduction of meristem size. One of the salt stress-induced changes in the ultrastructure of the shoot apical meristem was cell vacuolation. Jbir et al. (2003) reported that the cell vacuolation is increased by salt stress, especially in the salt-sensitive species. They showed that the salt stress-induced changes in the cells are characterized by further development of destructive processes expressed in changes of the ultrastructure of all cell organells. Balnokin (1995) showed that the decrease in water content is accompanied by supply of Na+ in the cells. This can be explained by an increased permeability of the plasmalemma and other cell membranes due to a decrease in their barrier properties, which are realized through the function of ion pumping systems in the membranes.
An excess of ions in the cytoplasm induced degradation of all organells, except the nucleus, which is the most resistant to stress, but its structure is also affected (Werker et al., 1983; Avetisova and Kadykov, 1985).
In conclusion, it can said that salt stress induced a surge of changes in structure (reduction of size meristem and disorders in the meristem) and the ultrastructure of the shoot apical meristem (the formation of autophagic vacuoles that including damaged organells) in canola. These changes result in the reduction of water demands of meristematic cells.
REFERENCES
- Ashraf, M., N. Nazir and T. McNeilly, 2001. Comparative salt tolerance of amphidiploid and diploid Brassica species. Plant Sci., 160: 683-689.
Direct Link - Cheeseman, J.M., 1988. Mechanisms of salinity tolerance in plants. Plant Physiol., 87: 547-550.
CrossRefDirect Link - Hanif, M. and M.S. Davies, 1998. Effects of NaCl on meristem size and proximity of root hairs to the root tips in Secale cereale (cv. K2) and Triticum aestivum (cv. Chinese spring). Pak. J. Biol. Sci., 1: 15-18.
CrossRefDirect Link - Huang, C.X. and R.F.M. van Steveninck, 1988. Effect of moderate salinity on patterns of potassium, sodium and chloride accumulation in cells near the root tip of barley: Role of differentiating metaxylem vessels. Physiol. Plant, 73: 525-533.
CrossRefDirect Link - Jbir, N., A. Said and A. Ayadi, 2003. Root cell characteristics of two species differing in their sensitivity of NaCl in response to salt stress. J. Trace Microprobe Tech., 21: 707-712.
Direct Link - Sacks, M.M., W.K. Silk and P. Burman, 1997. Effect of water stress on cortical cell division rates within the apical meristem of primary roots of maize. Plant Physiol., 114: 519-527.
CrossRefDirect Link - Sanders, D., C. Brownlee and J.F. Harper, 1999. Communicating with calcium. Plant Cell, 11: 691-706.
CrossRefDirect Link - Stephen, H., K.M. Volkmar and P.R. Miller, 2001. Comparing canola, field pea, dry bean and durum wheat crops grown in saline media. Crop Sci., 41: 1827-1833.
Direct Link - Werker, E., H.R. Lerner, R. Weimberg and A. Poljakoff-Mayber, 1983. Structural changes occurring in nuclei of barley root cells in response to a combined effect of salinity and ageing. Am. J. Bot., 70: 222-225.
Direct Link - West, G., D. Inze and G.T.S. Beemster, 2004. Cell cycle modulation in the response of the Primary root of Arabidopsis to salt stress. Plant Physiol., 135: 1050-1058.
Direct Link