
Mashark S. Abdulai
Savanna Agricultural Research Institute, P.O. Box 52, Nyankpala, Tamale, Ghana
Journal of Agronomy
Year: 2006 | Volume: 5 | Issue: 4 | Page No.: 634-640







ABSTRACT
This study utilized 400 S1 progenies or families and cycle bulks from two random mating populations planted in sole and in intercropping with soybean variety Pella-84. The objectives were to compare half-sib and S1 methods of selection for developing sorghum varieties and the nature of genotypes suitable for different cropping systems. Statistical significance tests showed that the progenies were different both within populations and between the populations. The progenies or families selected for sole cropping systems were later to flower than those selected for intercropping system. However, intercropping increased the days to flowering and maturity. Total yields were higher in intercropping due to the additional harvest of the soybean components. The ranking of the S1-families based on total grain yield in sole cropping system showed that the first twenty families were made of equal numbers of families from the two populations, an indication that a short term selection program in sole cropping system may produce genotypes suitable for both cropping systems. GE interaction effects for most of the characters evaluated were not significant suggesting that many more cycles were needed to create diversity between the two populations to reveal GE interaction effects. Estimates of heritability and the expected responses due to selection confirmed that much would be gained with further selection. Evaluation of bulks from different cycles of selection showed that higher yields were obtained from half-sib bulks than S1-bulks probably due to heterosis. Inbreeding depression was less in the bulks selected in intercropping systems. This suggests that homozygosity is likely to be achieved earlier in sole cropping than in intercropping, hence genotypes suitable for intercropping systems are likely to be more variable than those more suitable to sole cropping systems.
PDF Abstract XML References
How to cite this article
Mashark S. Abdulai, 2006. Genotypic Variation of Sorghum bicolor (L.) Moench in Response to Different Cropping Systems. Journal of Agronomy, 5: 634-640.
DOI: 10.3923/ja.2006.634.640
URL: https://scialert.net/abstract/?doi=ja.2006.634.640
DOI: 10.3923/ja.2006.634.640
URL: https://scialert.net/abstract/?doi=ja.2006.634.640
INTRODUCTION
The growing of two or more cultivars, simultaneously on the same area of land is termed intercropping (Andrews and Kassam, 1976). The use of intercropping in traditional farming systems is very old, yet is still being practiced by more than half the farmers of the world. Intercropping produces higher yields, and is characterized by low levels of purchased inputs, intensive labor and more uniform distribution of the workload throughout the year. Since the research by Andrews (1978, 1974) and the review of intercropping by Willey (1978a, b), many reports have been published recognizing these advantages many of them agree that besides increased yield per unit area for sorghum based systems (Singh et al., 2000), winter rapeseed systems (Reddy et al., 2001) and cotton-based systems (Khan et al., 2001), the practice is advantageous because it may reduce risk of crop failure, crop diseases, pests and weeds (Litsinger and Moody, 1976; Altieri and Liebman, 1986), erosion, soil water loss and spreads labor demands.
Recurrent selection schemes have been very successful in improving populations of sorghum (Doggett and Eberhart, 1968; Doggett, 1972; Ross and Gardner, 1983) to develop maize varieties for tolerance to drought (Monneveux et al., 2006). In sorghum breeding, the use of recessive genetic male sterility, and with harvest of male sterile plants each generation, ensures cross-pollination so that laborious hand crossing is not required to obtain population genetic recombination. These varieties are often developed for sole cropping systems. However, farmers grow them in different cropping systems other than what is recommended by the breeders.
The reviews on developing or adapting cultivars specifically for intercropping systems (Francis et al., 1978a; Francis, 1985; Smith and Francis, 1986) have emphasized the importance of genotype by cropping systems (GE) interactions, i.e., when the ranking of genotypes change from one environment to the other when evaluated. Significant genotype-cropping systems interactions have been reported by Badu-Apraku et al. (2003) for maize, Asante and Dixon (2002) for cassava and Shafii et al. (1992) for winter rapeseed yield and oil content. The basic causes of GE are believed to be due to biochemical pathways of certain physiological processes taking place in plants. Even though genotypes may be similar phenotypically, they may still differ by a few nucleotide sequences. Certain genes may be expressed in some environments while others may not, due to the sequence differences. As a result, these genotypes may react differently to conditions varying from one environment to the other as reported by Langridge and Griffing (1959) for hybrid Arabidopsis thaliana plants. The evidence of improved, early and/or short genotypes producing higher land equivalent ratio (above 1.51) while tall and late genotypes giving poor yields reported by Andrews (1974) suggests that different crop varieties may be adapted to different growing conditions.
Many of the studies on intercropping systems have concentrated on agronomic aspects, and have used cultivars developed for sole cropping systems. Nearly all reports pertain to those types of cultivars. To make a rational decision as to whether a special selection program is necessary (or possible) for developing sorghum genotypes better adapted to intercropping systems, this project was designed to test S1-families for genotype-cropping systems interactions and also to investigate the type of populations suited for intercropping systems. This information may be useful in the design of suitable breeding and selection systems towards the genetic improvement of sorghum varieties for use in intercropping systems.
MATERIALS AND METHODS
Two populations of sorghum (PSC and PIC) used in this study came from two cycles of recurrent selection for high grain yield in sole and intercropping systems by Dr. W.C. Youngquist in 1986. They were developed from a population previously developed by Dr. C.A. Francis (1986) who had derived them from the RP2B population of Dr. W.M. Ross (1981) at the University of Nebraska, Lincoln. Evaluations were done at the same site at the Agricultural Research and Development Center in Mead, Nebraska. The soil type is a sharpsburg silty clay loam (Mesic Typic Argiudoll). The blocks-in-replications design outlined by Ross and Gardner (1983) for the evaluation of S1 lines of sorghum was used for the S1 family evaluation. Cropping systems were used as whole plots with two replications. Two hundred families were selected from each of the two populations (PSC and PIC). Ten families from each population chosen at random were assigned to a set and there were a total of 20 sets. The families per set and the sets per cropping system (whole plot) remained together in the second replication, but were again randomized.
In order to estimate the gain due to selection, entries were also derived from the same populations but were bulks of equal quantities of seed from all the heads harvested from the isolated plots of cycle zero, cycle one sole cropping system (sterile and fertile) and cycle two sole cropping system (sterile and fertile). In addition to the bulk of the families, the bulks of the S1 families were included. These lines had been maintained in isolations of sole cropping and intercropping systems. In all, ten bulks were formed by the procedure described above. A randomized complete block design with eight replications was used for evaluating the bulk family trial. Each replication consisted of sole cropped and intercropped whole plots with ten entries each.
In both trials, the sorghum families and soybeans were planted in single-row plots 8.0 m long with 0.76 m spacing between rows. Excess seed was planted in each plot and stands were later thinned to about 0.15 m between plants for the sole plots of sorghum. The intercropped plots were thinned to one plant of sorghum to two plants of soybeans. No thinning was done to the soybeans but it was expected that the intercropping will give plant densities in replacement series of 1/3 sorghum and 2/3 soybeans. The commercial soybean variety used for this study was Pella-86.
The rate at which the seedlings were growing was estimated by visually assessing the height of the seedlings two weeks after germination. The seedling vigor was rated on a scale of 1 to 5, 1 being excellent and 5 being very poor.
Days to 50% flowering (DFF) for each plot was determined by visual estimations. It was noted as the date on which at least half of the main heads in a plot had reached 50% anthesis and was converted to the number of days from planting to fifty percent bloom.
The number of plants in a plot and the number of heads with grain were counted. These were adjusted to plants and heads per square meter. The grain weight per head was calculated from the grain weight per plot divided by the number of plants per plot. The height of plants per plot was measured as a visual estimate of the mean height of the top of the main heads in meters.
Harvesting was started after the main sorghum heads matured. Both the sorghum and soybeans were combined harvested together. The harvest was taken to the laboratory and dried for two days, then the beans were separated. The weights and moisture content of grain of each crop was determined.
The data were entered into a personal computer and analyzed with SAS (1996). The data from each cropping system was analyzed separately. Scatter diagrams of each character measured in the intercrop was plotted against the same character measured in sole cropping to find the relationship of the character in each system. The grain yields of sorghum and soybeans of the intercrop plots were converted to a combined yield. This was done by first standardizing the yields of both sorghum and soybeans of both cropping systems to a mean of zero and a standard deviation of one. The standard yields were summed in each plot within the cropping systems. These were then re-standardized to the monoculture mean yield of 2.484 and standard deviation of 0.872. That resulted in a combined yield which is equally correlated to both component crops.
The S1-family evaluation trial was analyzed as an incomplete blocks-in-replications design considering a mixed model. Cropping systems were considered fixed while genotypes being random. The bulk family testing was analyzed using randomized complete block design with eight replications and two cropping systems. Analysis of variance within each cropping system was performed on the variables measured in each plot and the genotypic variation was partitioned into between and within-population components. Estimates of heritability on progeny mean basis were calculated.
RESULTS AND DISCUSSION
Evaluation of S1 families: In the random mating populations there were many plant types in each population owing to the genetic variation that was created through selection. The mean squares for the analyses of the sorghum traits measured within each cropping system, and analyses of combined cropping systems are presented in Table 1 and 2, respectively. There were numerous signs of water stress in the intercrop plots on hot and dry days more than their counterparts in sole. That resulted in stunting and stem lodging in seriously affected plots. Genotypes which could not efficiently use the limited resources, were weak and slow-growing during the seedling stage so were almost completely smothered by the soybeans. The reports of Andrews (1972) and Wahua and Miller (1978) suggest that the efficiency of production in cereal-legume intercrop systems could be improved by minimizing inter-specific competition between the component crops for growth factors by growing component crops with contrasting maturities so that they complement rather than compete for the same resources at the same time. For this study, an objective was to create conditions that will exert competition on the sorghum so that genotypes that are selected may be better adapted to intercropping systems.
The mean height of the families population one (PSC) was larger perhaps because it was selected in favorable environments and so had the chance to accumulate the genes for height. It was also later to mature. Selection for high yielding genotypes in sole cropping lead to the selection of tall and later maturing plants because yield was positively correlated with plant height and maturity (Table 4). This is consistent with the reports of Jan-Orn (1976), Ekebil et al. (1977) and Ross et al. (1981).
| Table 1: | Mean squares of individual cropping system analysis |
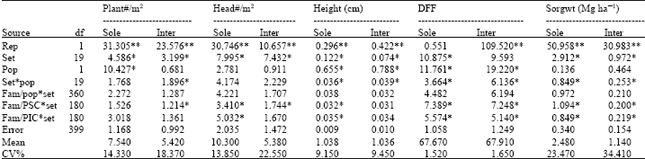 | |
| ns = not significant, ***p<0.001, *p<0.05, PSC = population selected under sole cropping, **p<0.01, PIC = population selected under intercropping | |
| Table 2: | Mean squares of traits analyzed across cropping systems |
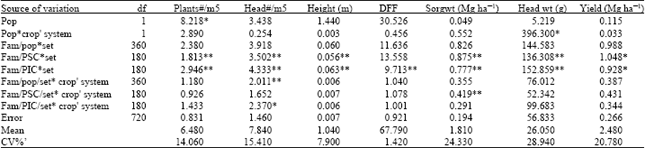 | |
| *p<0.05, **p<0.01, farm = families = progenies Crop’system = cropping system, Yield = combined yield of sorghum grain and soybeans converted to sorghum equivalent yield | |
It was surprising to observe that the yield of population one was lower (not significant at 0.05) than that of population two (PIC) in sole and the reverse was true in intercrop. The reason for this result is unclear, but could be due to chance or due to the fact that population two which was subjected to both inter- and intra-specific competition during selection was able to grow and yield better in sole cropping system in the absence inter-specific competition. The slight difference in days to 50% flowering of both populations (Table 3) in intercropping could be due to the limited resources that accounted for the reduction of the rate of growth and maturity in intercropping. This delay of maturity has been reported by Galwey et al. (1986) for grain sorghum intercropped with cowpeas. There was advantage to intercropping due to the combined yields of the component crops. The maximum yields were always obtained in intercrop and the minimum yields were not as low as those obtained in sole cropping system. The characters of the two populations had similar genotypic and phenotypic correlations in each of the systems (Table 4). This could be due to the fact that both were derived from the same source and selection was not practiced long enough to bring diversion. The correlations of the characters measured in sole and those measured in intercrop were high and similar to those reported by Galwey et al. (1986). This indicates that each genotype performed similarly relative in each other in each cropping systems. The negative genetic correlation coefficients obtained between head weight, and plant height and days to 50% flowering and plant height indicated an inherent association of these traits. These associations show that selection for both shortness and earliness will be difficult but selection for shortness and large head sizes is likely to be successful. In this case selection for earliness and large heads will be more difficult in population one than in population two. Sorghum grain yield, which was positively correlated with all the traits indicate that to breed for high grain yield there would be a need to modify all of them. The closeness of the genetic variances of the traits (Table 5) suggests that two cycles are not enough to bring significant genetic variance differences between the populations derived from the same source. Except for grain yield and head weight of population one there were no significant interactions of the genotypes with cropping systems. The interaction was probably due to the slight superiority in grain yield PSC had over PIC in the intercropping system or due to random errors. If the cause is due to the superiority in yield then the results are consistent with the remark of Francis et al. (1978) that initial selection and screening is more efficient in sole cropping system than intercropping system. Due to the inconsistency of genotype x cropping system interactions, Francis et al. (1978) have suggested that early generation evaluations be done in sole cropping system while advanced selections in specific generations such as F5 and F8 in association with other crops.
The heritability estimates (Table 6) were biased as the numerators included both additive and non-additive genetic sources of variation, hence the high values obtained. However, they were similar in magnitude with those calculated by Jan-Orn et al (1976), Ekebil et al. (1977) and Knapp et al. (1987) for all the traits except head weight. This was probably due to the growth habit of the population.
| Table 3: | Populations means for characters evaluated in the different cropping systems |
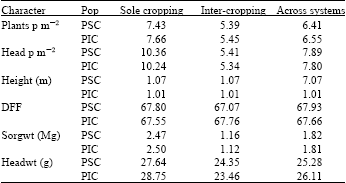 | |
| PSC and PIC are populations selected under sole- and intercropping respectively | |
| Table 4: | Genetic correlation coefficients of sorghum characters |
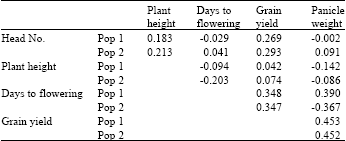 | |
| Table 5: | Estimates of family components of variance and their standard errors calculated |
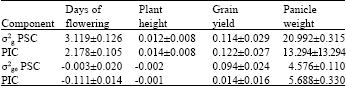 | |
| Table 6: | Comparison of estimated heritabilities of some sorghum traits based on S1-progeny means |
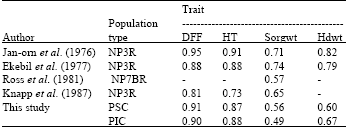 | |
| Table 7: | Means and least significance differences of family bulks in the two cropping systems |
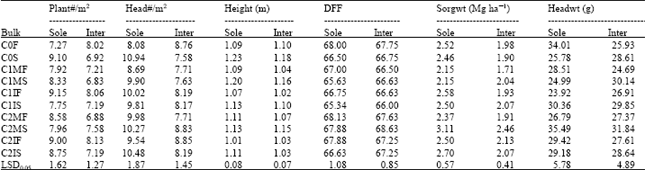 | |
| Table 8: | Percentage reduction in grain yield in S1-progeny bulks compared to HS-progeny bulks in sole and in intercrop |
 | |
| Note: The half-sibs are those bulks obtained from sterile plants while the S1s are those obtained from fertile plants | |
Days to 50% flowering and height which are highly heritable so had the highest values. The result is that it becomes very difficult to select high yielding genotypes without the tendency to select tall and late maturing genotypes due to the high correlation these traits have with high grain yield.
The predicted gains per cycle calculated for each population are 0.072 and 0.066 Mg ha-1, respectively. This amounts to 4.00 and 3.65% yield increases if the selection intensity is 0.2. This rate of increase for the intercropping system would be tremendous.
Evaluation of family: It is obvious from Table 7 that the mean values for plant population and head number per square meter are low in the intercrop because of the presence of soybeans. The mean values for the characters show that the bulks ranked in nearly the same order in each of the cropping systems. This is an indication that there is not enough selection in cycle two to affect any significant differences between the bulks selected on the basis of either cropping system. Nevertheless, bulks of the first cycle had lower means than their cycle two counterparts indicating a gain from selection in one cycle. So it is obvious that with selection for the best genotypes the response will significantly raise the yield of the bulks in advanced generations. The same trend was observed in the standardized yields. It was however, disappointing to note that there was no advantage to intercropping when the sole and intercrop yields of each bulk were compared. This is because the sorghum population was probably far above the optimum in the intercrop but below the optimum for sole cropping, thus the beans which were the sole contributors to intercrop advantage produced yields below the efficiency levels. This explanation is consistent with the reports of many researchers reviewed by Ofori and Stern (1987). The low yield could also be due to the losses during harvesting with the combine and the subsequent separation of the seeds. It can therefore be concluded that yield advantages in intercrop might be obtained when the plant population is optimum and the method of harvest modernized to economically get all the produce from the field through the use of combined harvesters.
The half-sib bulks (C0S, C1MS, C1IS, C2MS and C2IS) were taller than their counterpart S1-family bulks. This was probably due to heterosis. The bulks formed from the genotypes selected on the basis of intercropping systems generally performed better in sole cropping system. This can be attributed to the removal of the inter-specific competition in sole hence the ability for plants to express their yield potential. Usually inbreeding is applied to cross-pollinated crops such as pearl millet and maize but not to a self-pollinated crop like sorghum since inbreeding is the normal system of mating. However, the use of genetic male sterility has converted the plants in this study to behave as cross-pollinated ones. The inbreeding depression expressed as percentage reduction in yield of the S1-progeny bulks compared to the half-sib progeny bulks is listed in Table 8. The populations selected for multiple cropping systems (C1I and C2I) have significantly less inbreeding depression compared to the populations (C1M and C2M) selected for sole cropping, a reflection on the ease with which homozygosity is likely to be achieved in sole than in intercropping systems. It also implies that a stable variety or population selected for intercropping systems is likely to be heterozygous at most loci or be a blend of varieties. This is usually the observation in farmers’ fields.
CONCLUSION
Both sorghum grain and soybean yields were low due to losses during harvesting and handling. Genotypes growing in sole cropping produced bigger heads than those growing in intercropping. Sorghum grain yield differed among the genotypes in each of the cropping systems. An advantage to intercropping by using combined yield was observed for both populations but the population selected based on sole cropping performed better in intercropping system. The ranking of the S1-families based on total grain yield in sole showed that the first twenty families were made of equal numbers of families from the two populations, an indication that a short term selection program in sole may produce genotypes suitable for both cropping systems. The maturity of sorghum was delayed by about one day in intercropping than when grown in sole cropping. The genotypes selected on the basis of sole cropping were later to flower than their counterparts selected based on intercropping. On the average cropping systems did not have any effects on height of the families. Selection against tallness was achieved in both systems when yield was not the factor. Otherwise, yield was positively correlated with height. In general the GE interactions for most of the characters evaluated were not significant. Many more cycles are needed to create diversity between the two populations when GE interaction effects might be revealed. Estimates of heritability and the expected responses due to selection confirmed that there is much to gain with further selection. A comparison of the bulks obtained by bulking equal quantities of seed from the heads of all plants harvested in each cycle showed that higher yields were obtained from half-sib bulks than S1-bulks. Inbreeding depression was less in the bulks selected in intercropping systems than their counterparts selected in sole cropping systems. Homozygosity is therefore, likely to be achieved in sole cropping than in intercropping, hence genotypes suitable for intercropping systems are likely to be more variable than those more suitable to sole cropping systems.
ACKNOWLEDGMENTS
I wish to express my sincere gratitude to Dr. Wayne C. Youngquist, Dr. William Compton and Prof. David J. Andrews of the University of Nebraska, Lincoln for the supervision and inspiration given me during the conduction of this study. I thank my colleagues at the Savanna Agricultural Research Institute for their criticism during the preparation of this manuscript.
REFERENCES
- Andrews, D.J., 1974. Responses of sorghum varieties to intercropping. Exp. Agric., 10: 57-63.
Direct Link - Andrews, D.J. and A.H. Kassam, 1976. Importance of multiple cropping systems in increasing world food supplies. In multiple cropping. Am. Soc. Agron., 27: 1-10.
Direct Link - Badu-Apraku, B., F.J. Abamu, A. Menkir, M.A.B. Fakorede, K. Obeng-Antwi and C. The, 2003. Genotype by environment interactions in the regional early maize trials in West and Central Africa. Maydica, 48: 93-104.
Direct Link - Doggett, H. and S.A. Eberhart, 1968. Recurrent selection in sorghum. Crop Sci., 8: 119-121.
Direct Link - Ekebil, J.P., W.M. Ross, C.O. Gardner and J.W. Maranville, 1977. Heritability estimates, genetic correlations, predicted gains from S1 progeny, test in three grain sorghum random-mating populations. Crop Sci., 17: 373-377.
Direct Link - Francis, C.A., M. Prager, D.R. Laing and C.A. Flor, 1978. Genotype H environment interactions in bush bean cultivars in monoculture and in associated with maize. Crop Sci., 18: 237-242.
Direct Link - Francis, C.A., 1985. Variety development for multiple cropping systems. Crit. Rev. Plant Sci., 3: 133-168.
Direct Link - Galwey, N.W., M.A. de Queiroz and R.W. Willey, 1986. Genotypic variation in the response of sorghum to intercropping with cowpeas and in the effect on the associated legume. Field Crops Res., 14: 263-290.
Direct Link - Jan-Orn, Jinda, C.O. Gardner and W.M. Ross, 1976. Quantitative genetic studies of the NP3R random-mating grain sorghum population. Crop Sci., 16: 489-496.
Direct Link - Knapp, S.J., W.M. Ross and W.W. Stroup, 1987. Precision of genetic variance and heritability estimates from sorghum populations. Crop Sci., 25: 265-268.
Direct Link - Litsinger, J.A. and K. Moody, 1976. Integrated pest management in multiple cropping systems. Am. Soc. Agron., 27: 293-316.
Direct Link - Monneveux, P., C. Sanchez, D. Beck and G.O. Edmeades, 2006. Drought tolerance improvement in tropical maize source populations. Crop Sci., 46: 180-191.
CrossRefDirect Link - Ofori, F. and W.R. Stern, 1987. Cereal-legume intercropping systems. Adv. Agron., 41: 41-90.
Direct Link - Reddy, G.S., V. Maruthi and M. Vanaja, 2001. Effect of soil depth on productivity of sorghum (Sorghum bicolor) and pigeonpea (Cajanus cajan) in sole and intercropping systems. Ind. J. Agric. Sci., 71: 510-515.
Direct Link - Ross, W.M., K.D. Kofoid, J.W. Maranville and R.L. Voigt, 1981. Selecting for grain protein and yield in sorghum random-mating populations. Crop Sci., 2: 774-777.
Direct Link - Shafii, B., K.A. Mahler, W.J. Price and D.L. Auld, 1992. Genotype × environment interaction effects on winter rapeseed yield and oil content. Crop Sci., 32: 922-927.
CrossRefDirect Link - Wahua, T.A.T. and D.A. Miller, 1978. Relative yield totals and yield components of intercropped sorghum and soybeans. Agron. J., 70: 287-291.
Direct Link - Willey, R.W., 1979. Intercropping-its importance and research needs. Part 1. Competition and yield advantages. Field Crop Abst., 32: 1-10.
Direct Link