
S. A. Sadat Noori
Department of Agronomy and Plant Breeding, Abouraihan Campus, University of Tehran, P.O. Box 4117, Tehran, Islamic Republic of Iran
A. Roustaei
Department of Agronomy and Plant Breeding, Abouraihan Campus, University of Tehran, P.O. Box 4117, Tehran, Islamic Republic of Iran
B. Foghi
Department of Agronomy and Plant Breeding, Abouraihan Campus, University of Tehran, P.O. Box 4117, Tehran, Islamic Republic of Iran
Journal of Agronomy
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 131-136







ABSTRACT
Whole plant responses to salinity and their interaction to different conditions were evaluated for vegetative and productive traits of wheat. For this purpose salinity tolerance was examined for 75 families produced from crossing 25 F2 plants from crosses between Siete Cerros (salt tolerant) and Axona (salt sensitive) spring bread wheats, to their parents and their F1 progeny. The 75 families and their parents were grown under 0 (control) and 50 mM NaCl. Data for eleven metric traits, days to heading, days to maturity, final plant height, spike length, ear weight, straw weight, number of grains per ear, grain yield per plant, 000 grain weight, whole plant weight and harvest index was measured. Salinity reduced performance with regard to all the above traits. The genotype x concentration interaction was highly significant for yield components, indicating that plants responded differently in yield components when exposed to the different levels of salinity. In contrast, the genotype x concentration interactions for the vegetative traits, heading date, maturity date, final plant height, weight of straw and whole plant weight were not significant and or slightly significant indicating a lower differentiation response to salinity. This may be because a limited number of genes control the expression of vegetative more than productive traits in wheat when exposed to salinity.
PDF Abstract XML References
How to cite this article
S. A. Sadat Noori, A. Roustaei and B. Foghi, 2006. Variability of Salt Tolerance for Eleven Traits in Bread Wheat Grown in Different Saline Conditions. Journal of Agronomy, 5: 131-136.
DOI: 10.3923/ja.2006.131.136
URL: https://scialert.net/abstract/?doi=ja.2006.131.136
DOI: 10.3923/ja.2006.131.136
URL: https://scialert.net/abstract/?doi=ja.2006.131.136
INTRODUCTION
Salinity has been and will continue to be a major factor limiting agricultural and crop production in many areas of the world due to both natural and man-made circumstances. In some soils salinity arises from parent materials containing salt, but in agricultural soils, especially in arid and semi-arid regions, it is caused by man due to the effects of irrigation with low quality water, poor water management and inadequate drainage systems. The problem is compounded by the relatively low salt tolerance of most crop plants. The high yielding crop varieties of such as wheat, rice and maize, which are available today and those responsible for much of the production increases of the past decades, are generally adapted to and yield only in, favourable physical, economic and biological condition's. Genetic adaptation of crop species to saline soils is currently receiving considerable attention to produce plants displaying resistance to salinity stress.
Breeding crops for salt tolerance would be likely to provide an economic and efficient method of overcoming saline soil problems. Such a crop improvement programme must be based on adequate variability for salinity tolerance and such variation has been observed between and within species including landraces and cultivars. Unusual achievements have been made during the last few decades through selection and breeding for the salinity tolerance in important crop plants. Wheat is a major food crop grown in most of the countries in arid and semi-arid regions and these contain most of the saline soil areas. As a consequence its production is restricted by salinity effects. Kingsbury and Epstein (1984) and Ashraf and McNeilly (1988) have shown that wheat is also a crop in which there is variation for salinity tolerance and this may be used for improving wheat for salinity tolerance. As yet, no single factor or groups of factors have been identified as promoting salinity tolerance; rather it is a complex whole plant phenomenon (Ashraf and McNeilly, 1988; Toenniessen, 1984; Yeo and Flowers, 1994). Greenway and Munns (1980) reported many examples in which the mechanism of salt tolerance varied from cultivar to cultivar within species, although in general mesophytes exclude ions when exposed to saline conditions. This situation also has been reported in millet (Ashraf and McNeilly, 1987), Rice (Akbar and Yabuno, 1993), wheat (Kingsbury and Epstein, 1984) where no single criterion could be used to indicate tolerance, because salt sensitivity varies greatly with growth stage, species, variety and ecotype. This would complicate the screening and selection process if it was based on a single growth stage, but whole plant responses does seem to present a useful indication of tolerance. A second reason to use the concept of yield potential in studies of stress resistance is that it provides to agronomist an estimate of a yield ceiling attainable with present technology. Such an estimate is extremely useful to quantify the extent of yield reduction caused by the stresses in question (Acevedo and Aereres, 1993).
In present research, whole plant responses to salinity and their interaction to different conditions were evaluated for vegetative and productive traits of wheat. For this purpose we used the data obtained from a Triple Test Cross programme to examine the response to saline and non-saline conditions of eleven metric traits of seventy-seven bread wheat families (Triticum aestivum) derived from cross between Siete Cerros (salt tolerant) and Axona (salt sensitive).
MATERIALS AND METHODS
F1 and F2 generation material from the wheat varieties Siete Cerros (salt tolerant) and Axona (salt sensitive) were produced during the years 2001, 2002 and 2003. From the F2 population, 25 plants were selected randomly and used as female parents for crossing with the respective male testers P1, P2 and F1, designated as L1, L2 and L3 respectively, from which families, L1i, L2i and L3i were produced. The 75 families thus produced and their parents, were raised in a randomized complete block design in two replicates, under two (normal and stress) environmental conditions, each replicate consisting of one hundred and fifty experimental units (pots).
All seed was surface sterilized first in 70% (v/v) ethanol 1 min and then in 5% (w/v) sodium hypochlorite (6 min) and finally washed three times with distilled water. Seeds were germinated on moist filter paper in petri dishes. The experiment was carried out in a glasshouse at day temperature 22±2°C and night temperature 16±2°C with natural daylight supplemented by 400 watt mercury vapour lamps to give a 16 h day length. The plants grown in 18 cm diameter x 19 cm deep plastic pots filled with 4.40 kg washed river sand (oven-dried weight). The sand was thoroughly washed with tap water for one week, followed by three washings with full strength nutrient solution (Hewitt, 1966).
Two NaCI concentrations 0 mM NaCI (control), 1.37 dS m-1 and 150 mM, 17.21 dS m-1 were used, in full strength nutrient solution. Five seedlings (six-day-old) of each family were grown separately and equidistantly from each other in each pot. Salt treatments began 18 d after the start of the experiment and the salt concentration was increased stepwise in aliquots of EC 5.0 dS m-1 (25 mM) every other day until 150 mM NaCI was reached. Twice per week, 200 mL of distilled water was added to each pot to maintain sand moisture and to prevent salt accumulation. EC of the leachates was tested weekly.
The plants were harvested at maturity and measurements recorded in the 1540 samples [(5 plants from 75 progeny + 2 parents) per treatment] x 2 replicates, for the days to heading, days to maturity, final plant height (cm), spike length (cm), weight of ears (g), weight of straw (g), number of grains per spike, 1000 grain weight (g), whole plant dry weight (g), oven dried at 60°C for 72 h and harvest Index, grain yield per plant per whole plant dry weight.
RESULTS
The analyses of variance of all eleven metric traits are given in Table 1 and means and ranges for traits between the parents and among the families in saline and non-saline conditions are presented in Fig. 1.
Days to heading: It was not affected to a great extent by any saline treatment. The salt tolerant parent, Siete Cerros, flowered later in the salinity treatment than in the control. The sensitive parent Axona began flowering on average a few days later growing in the control than in the salinity treatment. The remaining plants were mostly intermediate in flowering date (Fig. 1).
Days to maturity: Differences between families and parents were significant (p<0.001) for maturity date. Genotype x concentration was also significant (p<0.05) reflecting the different responses of the families/parents to 150 mM NaCI. Differences between mean number of days to maturity indicate that plants reached maturity earlier in 150 mM NaCI than in the control (p<0.001), with the exception of the salt tolerant parent Siete Cerros (Fig. 1).
Final plant height: Salinity reduced this character in both parents and families and reduction was more in the sensitive Axona than in Siete Cerros (Fig. 1), the differences being significant (p<0.001).
Spike length: Overall, spike length was reduced significantly (p<0.001) by salinity and its length reduction was more in Axona than in Siete Cerros (Fig. 1).
Weight of ears: 150 mM NaCI reduced ear weight significantly (p<0.001) from that of the control in all 75 families and Axona was more sensitive than Siete Cerros.
| Table 1: | Mean squares from analysis of variance for eleven metric traits for families derived from cross between two bread wheat (Siete Cerros x Axona) and their parents grown in saline (150 mM NaCI) and non-saline conditions |
 | |
| *** = p < 0.001, ** = p < 0.01, * = p < 0.05 and NS = Non-significant | |
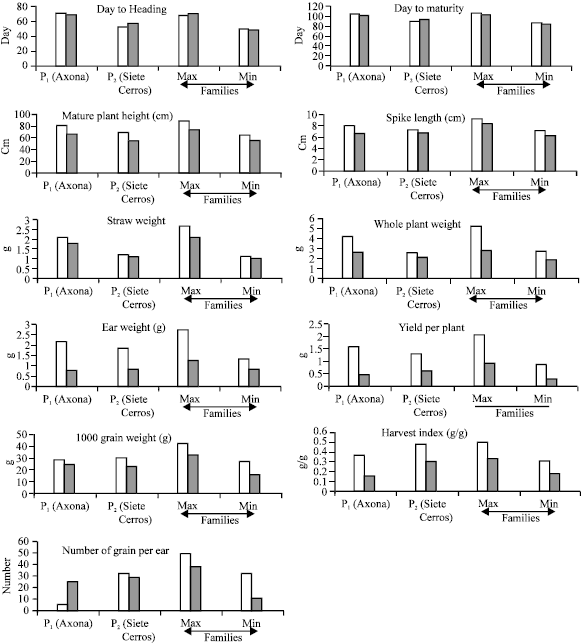 | |
| Fig. 1: | Chart of range for traits between the parents (P1 = NaCl Sensitive and P2 = NaCl Tolerant) and among the families (Maximum and Minimum) in saline (Black colour) and non saline (White colour) conditions |
At 150 mM NaCI Axona produced few seeds, but Siete Cerros produced more seeds. There was considerable variation in ear weight in the 75 families, particularly in the plants grown in the control (Fig. 1).
Weight of straw: Straw weight was, overall, reduced significantly (p<0.001) by salinity in all plants, but not as would be expected in the control. Most of the plants had only one tiller in the NaCI treatment and vigour was clearly reduced. The reduction in straw weight was more in the sensitive parent Axona than in the tolerant Siete Cerros as would be expected (Fig. 1). Some plants in NaCI treatment began tillering after that the main tiller matured (after about three month).
Whole plant weight: Whole plant weight was significantly (p<0.001) less in the plants grown at 150 mM NaCI than those in the control for all families, the reduction being greater in Axona than in Siete Cerros (Fig. 1). The water usage by the plants was less in the saline condition than in the control, most of the saline solution remaining in the watering saucers up to 48 h after treatment.
Yield components: Seeds per ear, grain yield per plant and 1000 grain weight differed significant differently (p<0.001) between families/parents and salinity reduced them significantly (p<0.001) in all families and were reduced more in Axona than in Siete Cerros (Fig. 1). Grain per ear and 1000 grain weight varied between the families, in those plants grown in saline treatment. In all 75 families and parents, grain yield per plant was less variable than in others grown in saline treatment.
Harvest index: 150 mM NaCl significantly (p<0.001) reduced harvest index in families and parents compared with control (Fig. 1). Differences between families/parents were also significant (p<0.001). Harvest index varied considerable across the families, grown in the NaCl treatment.
DISCUSSION
Wheat is categorized as moderately salt tolerant (Maas and Hoffman, 1977) but seed germination and seedlings in their early stages of growth are sensitive to salinity (Francois and Mass, 1993) . There is also further variation among wheat plants grown in saline condition's (Martin et al., 1994) suggesting that opportunities exist to increase salt tolerance in wheat through selection and breeding. From screening more than 5000 bread wheat accessions from the USDA World Wheat Collection (Kingsbury and Epstein, 1984) isolated a number of salt tolerant spring wheat lines. Differences in salt tolerance in wheat varieties in Pakistan and differences in salt tolerance at all growth stages, were found by (Qureshi et al. 1980). From these studies, salt tolerance at the seedling stage was considered to be useful indicator for use as a preliminary screening of wheat varieties. In another attempt, Ashraf and McNeilly (1988). reported that the measure of tolerance obtained from different growth stages varies as previously reported for wheat by Srivastava and Jana (1984). However, assessment of salinity tolerance based upon whole plant growth in saline conditions, is ultimately necessary. Thus this is the main subject investigated by the present work.
Development of crop varieties that combine high yield potential and high salinity tolerance will not be an easy task. Salinity tolerance is a complex whole plant character with physiological and biochemical functions controlled by numerous genes. Moreover, other environmental and soil factors influence its expression (Toenniessen, 1984). It has been argued that greater and more rapid advancement in developing salinity tolerance can be gained by concentrating efforts on generating variability, which would allow reduction in the effort spent on exploitation of variability (Ramage, 1980). Thus in the present work a segregating generation was used.
Ashraf (1994). tested some F2 populations derived from crosses of salt tolerant cultivars for salinity tolerance at 24 dS m-1 NaCI + CaCl2 based upon number of tillers per plant, 100 seed weight, number of grains per plant and seed yield. He concluded that improvement in salinity tolerance in spring wheat is possible through selection and breeding.
It is clear from the analyses of data presented here (Table 1) that genotypes considerably differed significantly as expected because these families were derived from segregation generations. The characters measured were reduced by saline effect as found by Maas and Grieve (1990). for yield components i.e., spike length, number of spikelets per spike and number of kernels per spike and also for number of tillers per plant. Grieve and Francois (1992) reported also such reduction in yield components and straw yield under salinity treatment in wheat.
The genotype x concentration interaction was highly significant (p<0.001) for yield components namely, number of grains per ear, grain yield per plant, 1000 grain weight and harvest index and was also significant (p<0.01) for weight of ear and spike length. This suggests (Kearsy, 1993). the presence of multigenic factors governing the expression of these economic traits which may switch on or off in responses to salinity.
Since grain yield and its stability are two major goals in plants selected for salt tolerance, it is natural that grain yield is frequently used as a criterion to measure responses of genotypes to salt stress. However, selection for grain yield is inefficient because of its low heritability, especially under stressful field conditions (Jana, 1993) Shannon (1985). emphasizes the role of ontogenetic drift in plant responses to salt stress. Because a plant's salt injury depends on its ontogenetic stage as well as on the nature and severity of previous stresses, it is desirable to measure responses to salt stress at different growth stages.
It is interesting that genotypexconcentration interactions in responses to salinity were highly significant (p<0.001 or 0.01) for yield components such as number of grains per ear, grain yield per plant, 1000 grain weight, harvest index, weight of ear and spike length. On the other hand, genotype x concentration interactions for vegetative traits such as heading date, maturity date, final plant height, weight of straw and whole plant weight were not significant or only slightly so (p<0.05) in responses to salinity. Possibly this indicates a more limited number of genes controlling the expression of these traits or that gene expression is more stable. A greater reduction in grain yield than in vegetative growth under saline conditions has been found in maize, Zea mays L. (Kaddah and Ghowail, 1968). rice, Oryza sativa L. (Pearson, 1959). and sorghum, Sorghum bicolor L. Moench (Francois, et al. 1984). In contrast, however, Francois et al. (1986) reported vegetative growth and straw yield, in both semi-dwarf and durum wheat was decreased more by soil salinity than was grain yield.
Salt tolerance is a complex character and is highly influenced by environmental factors, such as soil, temperature, light and type of salinity. Combined efforts in plant breeding, plant physiology and soil chemistry, may lead to some success. Information on the relationship between salt tolerance and any morphologic character may provide some marker genes for salt tolerance.
ACKNOWLEDGMENTS
Financial support to S.A. Sadat Noori was granted by Ministry of Science, Research and Technology of Iran and University of Tehran, to carry out these studies and is gratefully acknowledged.
REFERENCES
- Ashraf, M. and T. McNeilly, 1987. Salinity effects on five cultivars/lines of pearl millet (Pennisetum americanum L.). Plant Soil, 103: 13-19.
CrossRefDirect Link - Ashraf, M. and T. McNeilly, 1988. Variability in salt tolerance of nine spring wheat cultivars. J. Agron. Crop Sci., 160: 14-21.
CrossRefDirect Link - Ashraf, M., 1994. Genetic variation for salinity tolerance in spring wheat. Hereditas, 120: 99-104.
Direct Link - Francois, L.E., T. Donovan and E.V. Maas, 1984. Salinity effects on seed yield growth and germination of grain sorghum. Agron. J., 76: 741-744.
PubMed - Francois, L.E., E.W. Maas, T.J. Donovan and V.L. Youngs, 1986. Effect of salinity on grain yield and quality, vegetative growth and germination of semi-dwarf and durum wheat. Agron. J., 78: 1053-1058.
CrossRefDirect Link - Grieve, C.M. and L.E. Francois, 1992. The importance of initial seed size in wheat plant response to salinity. Plant Soil, 147: 197-205.
CrossRefDirect Link - Kaddah, M.T. and S.I. Ghowail, 1964. Salinity effects on the growth of corn at different stages of development. Agron. J., 56: 214-217.
CrossRefDirect Link - Kingsbury, R.W. and E. Epstein, 1984. Slection for salt resistant spring wheat. Crop Sci., 24: 310-315.
Direct Link - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Maas, E.V. and C.M. Grieve, 1990. Spike and leaf development in salt stressed wheat. Crop Sci., 30: 1309-1313.
Direct Link - Shannon, M.C., 1985. Principles and strategies in breeding for higher salt tolerance. Plant Soil, 89: 227-241.
CrossRef