
A. Lodhi
Department of Biology, Teachers College, P.O. Box 15758, Jeddah 21454, Saudi Arabia
F. Azam
Department of Biological Sciences, Faculty of Science, King Abdul Aziz University,
P.O. Box 80203, Jeddah 21589, Saudi Arabia







ABSTRACT
The effect of Baythroid (an insecticide) on dry matter yield and N yield of maize (Zea mays L.) was observed with particular emphasis on the uptake of N from soil and applied (15NH4)2SO4. Baythroid was applied @ of 0, 0.4, 0.8, 1.6, 3.2 and 6.4 mg kg-1 and N @ of 25 mg kg-1. The plants were harvested at two growth intervals i.e., 22 days and 47 days after sowing. At the two lower rates, Baythroid had no significant effect on dry matter yield of plants, while at the highest rate studied i.e., 6.4 mg kg-1, it had a negative effect. Maximum beneficial effect of Baythroid on dry matter yield and N uptake was observed at 3.2 mg kg-1. Trends in total plant N were similar to that for dry matter yield and a close correlation was observed between the two parameters. Baythroid application resulted in a significant increase in the uptake of both applied N and native soil N and the losses of applied N were substantially reduced.
PDF Abstract XML References
How to cite this article
A. Lodhi and F. Azam, 2002. Effect of Baythroid on Growth and Nitrogen Nutrition of Maize. Journal of Agronomy, 1: 38-40.
DOI: 10.3923/ja.2002.38.40
URL: https://scialert.net/abstract/?doi=ja.2002.38.40
DOI: 10.3923/ja.2002.38.40
URL: https://scialert.net/abstract/?doi=ja.2002.38.40
INTRODUCTION
In recent years, a large number of agrochemicals, including pesticides have been studied as nitrification inhibitors (Feng and Barker, 1990, Martinez-Toledo et al., 1992). By inhibiting nitrification, these chemicals may influence the internal cycling of nitrogen (N) in soil and its availability to plants. Many studies suggest that N use efficiency of crop plants is improved through the use of nitrification inhibitors. This improvement results mainly from a reduction in fertilizer N losses through denitrification and NO3¯ leaching (Aulakh et al., 1984). Due to nitrification inhibition, the plants will have NH4+ as the dominant N source for uptake and NH4+ fed plants are reported to exhibit higher rate of photosynthesis and dry matter accumulation (Mihailovic et al., 1992). In soil, prolonged availability of NH4+ will lead to higher microbial activity because microorganisms prefer NH4+ to NO3¯ (Recous et al., 1990). As a result, application/availability of NH4+ in soil will cause an increase in the mineralization and availability to plants of native soil N (Azam et al., 1991; 1993, Hart et al., 1986; Woods et al., 1987) through the so-called priming effect (Bingeman et al., 1953) or added N interaction (Jenkinson et al., 1985). However, the reports on the role of nitrification inhibitors in affecting the mineralization of organic N differ, both positive and no effect being observed (Chalk et al., 1990; Crawford and Chalk, 1993; Guiraud et al., 1989).
Previously, extensive studies were reported on the effect of Baythroid on soil microbial processes (Lodhi et al., 1994, 1996b, 2000) and plant growth (Lodhi et al., 1996a, Lodhi and Azam, 1998). These studies revealed a significant positive effect of Baythroid on N transformation processes as well as availability of N to crop plants. In this study, maize (Zea mays L.) was used as a test plant and employed 15N tracer methodology to study the fate of applied N in the soil-plant system as affected by Baythroid.
Materials and Methods
Soil: The soil was a silty clay-loam from the surface (0-15 cm) of an experimental field at the Nuclear Institute for Agriculture and Biology (NIAB), Faisalabad. The soil (< 2mm) contained 0.4% organic C, 0.05% total N, 18.5 mg kg-1 soil of inorganic N (NH4+ + NO3¯ + NO2¯), 19% sand, 40% silt and 41% clay. Soil pH (saturation extract) was 7.8. Analytical methods used have been described previously (Azam et al., 1994).
Baythroid: Baythroid is a highly effective and fast-acting insecticide manufactured by Bayer, Germany and contains Cyfluthrin (a synthetic pyrethroid) as the active ingredient. Baythroid used in the present studies is water-soluble. Uniformity of Baythroid in the solution was checked by determining optical density (230 nm) of aliquots taken from different depths of a column of aqueous solution.
Pot experiment: Five kg portions of the air-dried and sieved (< 2 mm) soil were filled in 6 kg plastic pots and treated as follows: T1, control (no treatment); T2, 25 mg N kg-1; T3, T4, T5, T6 and T7 received 0.4, 0.8, 1.6, 3.2 and 6.4 mg kg-1 of Baythroid (on an active ingredient basis), respectively in addition to 25 mg N kg-1 as (15NH4)2SO4 (1.0 atom % 15N excess). All pots including control received 25 mg P kg-1 and 6.2 mg K kg-1 as KH2PO4. The potted soil was sown to maize (Zea mays L., var. Akbar) using five seeds pot-1. Upon germination, the stand was thinned to three seedlings pot-1. The plants were grown under greenhouse conditions during the months of April-May-1991, when the temperature varied from 35-40 oC and relative humidity from 30-40%. The pots were irrigated as required with de-ionized water and after 22 days, triplicate pots from each treatment were harvested (harvest I). The remaining pots were harvested 47 days after planting (harvest II). Plant tops were clipped and the roots were carefully removed from the soil to achieve maximum recovery and washed. Dry matter yield of both root and tops was recorded and the material was finely powdered. Soil samples were air-dried and kept for further analysis.
Sub-samples of the plant material and soil were analyzed for Kjeldahl N using a modified digestion procedure to include of NO3¯ and NO2¯ (Bremner and Mulvaney, 1982). The digested samples were distilled following procedures of Mulvaney (1986) to reduce cross contamination of samples. The distillates were concentrated to 2-3 ml on a water bath after acidification with 0.2ml of 0.025M H2SO4 to avoid loss of N during heating. Isotope-ratio analyses of the samples were performed on a double inlet system mass spectrometer.
The data were subjected to analyses of variance followed by Duncan’s Multiple Range Test (DMRT). Co-efficient of correlations between different parameters were calculated by computer software programmes.
RESULTS
Dry matter yield of both root and shoot at the two harvest showed a significant improvement due to applied N, both in the absence and presence of Baythroid (Table 1). Compared to control, Baythroid had no significant effect on dry matter yield at the two lower levels of application (0.4 and 0.8 mg kg-1) and a negative effect at the highest level (6.4. mg kg-1) tested. On the other two levels, Baythroid caused a significant increase in dry matter of both shoot and root portions. At first harvest, net gain in total dry matter yield at 1.6 and 3.2 mg Baythroid kg-1 over N-only treatment was 12 and 28%, respectively, while the benefit increased to 19 and 40%, respectively, at harvest II. However, different treatments affected the dry matter distribution in root and shoot portions similarly at the two harvests as suggested by a close correlation between the two sets of data (r = 0.98).
As could be expected, N yield of plants increased significantly due to applied N at both the harvests (Table 2). The positive effect of applied N on N uptake increased significantly at 3.2 mg kg-1 Baythroid in the two harvests. Baythroid at 6.4 mg kg-1 soil had a negligible effect on N uptake at harvest I but a significantly positive effect at harvest II albeit lower than that observed at 3.2 mg kg-1 Baythroid. However, in general, Baythroid application at different levels along with N had variable effect on N yield of root and shoot portions. Trends in N yield were fairly similar to that observed for dry matter yield and a close relationship (r = 0.99) was observed between the two parameters at harvests I and II.
| Table 1: | Effect of Baythroid on dry matter yield (g pot-1) of maize and its distribution in root and shoot portions |
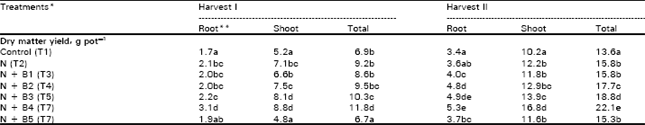 | |
| *, N was added at 25 mg kg-1;B1, B2, B3, B4, and B5 represent 0.4, 0.8, 1.6, 3.2, and 6.4 mg kg-1 Baythroid **, Figures in a column sharing a similar letter are not significantly different from each other at 5% level of probability according to DMRT | |
| Table 2: | Effect of Baythroid on N yield (mg pot-1) of maize and its distribution in root and shoot portions |
 | |
| * and ** see Table 1 for details | |
| Table 3: | Contribution of applied N (Ndff) and native soil N (Ndfs) to the plant N and the added N interaction (ANI) |
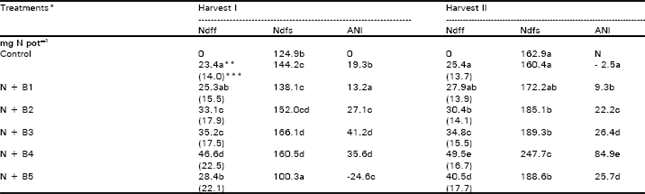 | |
| * and ** see Table 1 for detail***, Figures in parentheses indicate percent contribution of applied N to the total plant N | |
| Table 4: | .Percent of applied N taken up by plants (FNU), remaining in soil (FNR), or lost from the soil-plant system (FNL) |
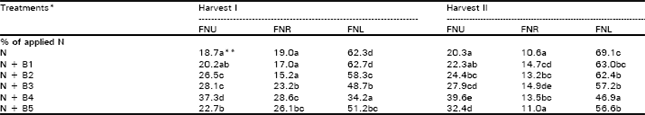 | |
| * and **, see Table 1 for details | |
Maize plants derived a greater portion of their N (78-86%) from the soil organic matter and the applied N made up only 14-22% of the total plant N (Table 3). Application of Baythroid at 8 mg pot-1 soil or above significantly enhanced the uptake of applied N and its contribution to the total plant N. At 6.4 mg kg-1 soil Baythroid, however net uptake of fertilizer N decreased in both the harvests but significantly more at harvest I. In both the harvests, maximum fertilizer N uptake was observed at 3.2 mg kg-1 soil Baythroid. Uptake of unlabeled N (presumably from soil organic matter) was also maximum at this level of Baythroid. Application of N, alone or together with Baythroid, increased the uptake of soil N except at 6.4 mg kg-1 soil Baythroid in harvest I. Increase in soil N uptake was attributable to the added N interaction (ANI) which was significantly greater in the presence of Baythroid. At 6.4 mg kg-1 soil Baythroid in harvest I, negative ANI was observed suggesting a significant inhibition of N mineralization from soil organic matter. However, harvest II showed a positive ANI at the highest level of Baythroid probably because of its degradation and removal of negative effects. Maximum ANI was observed at 3.2 mg kg-1 soil Baythroid.
Balance of applied N at the two harvests (Table 4) showed significant loss of applied N from the soil-plant system. The plants were able to use 18-37% was unaccounted at the time of harvest I. Except at 6.4 mg kg-1 Baythroid, there was no big increase in the uptake of applied N by plants, while decrease was apparent in the FNR. Loss of applied N occurred mainly by the 1st harvest and only a small addition to loss was observed at harvest II.
DISCUSSION
Nitrification inhibition is an established way to decrease fertilizer N losses through denitrification and NO3¯ leaching (Aulakh et al., 1984). Consequently, higher amounts of applied fertilizer N may be available to plants leading to improved biomass yield (Crawford and Chalk, 1993). Baythroid, which was previously found to be highly inhibitory to nitrification (Lodhi et al., 1994), had a significantly positive effect on dry matter yield and N uptake of maize. In wheat, Crawford and Chalk (1993) reported a positive effect of nitrification inhibition on dry matter and N yield. In wheat, lower levels of Baythroid had no significant effect, while at higher levels both straw and grain yield increased (Lodhi and Azam, 1998). In case of rice where Baythroid and N-serve were compared, the former had a negative effect on dry matter yield at higher levels of application (Lodhi et al., 1996a). So, the increase in dry matter could be attributed to a higher uptake of N since a close relationship was observed between these two parameters.
The increase in the uptake of N following nitrification inhibition may partially be attributed to a decrease in the loss of N and to a prolonged availability of NH4+-N. Loss of fertilizer N was indeed reduced in the presence of Baythroid. Persistence of plant-available N in NH4+ form will not only lead to greater microbial activity in the plant rhizosphere because of microbial preference for NH4+ as compared to NO3¯, but also to enhanced availability of soil N. In several studies, NH4+-N caused an increase in the mineralization and availability to plants of native soil N as a result of priming effect of the applied N or an added N interaction (Jenkinson et al., 1985, Hart et al., 1986, Woods et al., 1987, Azam et al., 1994). In this study, Baythroid caused an increase in the plant uptake of unlabeled N that presumably resulted from enhanced mineralization of native organic N in the soil leading to positive ANI. This ANI could be termed real as defined by Jenkinson et al. (1985), since in an incubation study under laboratory conditions, we have observed a significant increase in the mineralization of organic N in the presence of Baythroid (Lodhi et al., 1994). This increase is generally greater than the fertilizer N immobilized in soil, a situation supporting the occurrence of real ANI. In addition, root biomass increased in response to the applied N and Baythroid.
The source of plant available N may also influence dry matter accumulation. It is reported that maize plants supplied with NH4+-N had a higher chlorophyll content that may help plants maintain a higher rate of photosynthesis leading to greater dry matter yield (Mihailovic et al., 1992). Prolonged presence of NH4+ will also leave higher amounts of NO3¯ available for plant uptake since plants are reported to be less competitive with soil microorganisms for NH4+ (Schimel et al., 1989). In a pot experiment (unpublished), NH4+-N has a slight edge over NO3¯ in affecting the dry matter and N yield, with the two sources showing a synergistic effect.
The results indicate that insecticides may have significant potential to increase crop yields. The positive effects may result from changes in different microbial functions like nitrification and N mineralization. However, detailed studies under both laboratory and field conditions will be required to realize such effects. In the present study, Baythroid proved to be a desirable chemical not only as inhibitor of nitrification, but also as promoter of soil N mineralization and thus appeared to have no negative effects on soil micro flora other than nitrifiers (Lodhi et al., 1994).
ACKNOWLEDGMENTS
The authors gratefully acknowledge the mass spectrometric facilities provided by Dr. M. Ishaq Sajjad at RIAD, PINSTECH, Islamabad. Technical assistance of Mr. M. H. Sajjad and equipment donations by Alexander von Humboldt Foundation of Germany are also gratefully acknowledged.
REFERENCES
- Azam, F., R.L. Mulvaney and F.W. Simmons, 1995. Effects of ammonium and nitrate on mineralization of nitrogen from leguminous residues. Biol. Fertil. Soils, 20: 49-52.
CrossRefDirect Link - Bingeman, C. W., J. E. Varner and W.P. Martin, 1953. The effect of the addition of organic materials on the decomposition of an organic. Soil Sci. Soc. Am. Proc., 17: 34-38.
Direct Link - Crawford, D.M. and P.M. Chalk, 1993. Sources of N uptake by wheat (Triticum aestivum L.) and N transformations in soil treated with a nitrification inhibitor (nitrapyrin). Plant Soil, 149: 59-72.
Direct Link - Hart, P.B.S., J.H. Rayner and D.S. Jenkinson, 1986. Influence of pool substitution on the interpretation of fertilizer experiments with 15N. Eur. J. Soil Sci., 37: 389-403.
CrossRefPubMedDirect Link - Lodhi, A. and F. Azam, 1998. Yield and nitrogen uptake of wheat (Triticum aestivum L.) as affected by nitrapyrin and a nitrification inhibiting insecticide. Cereal Res. Commun., 26: 305-312.
Direct Link - Lodhi, A., N.N. Malik and F. Azam, 1996. Growth and nitrogen nutrition of rice (Oryza sativa L.) in soil treated with N-serve and a nitrification inhibiting insecticide. Pak. J. Bot., 28: 75-83.
Direct Link - Lodhi, A., F. Azam and N.N. Malik, 1994. Effect of baythroid on nitrogen transformations in soil. Biol. Fertil. Soils, 17: 173-176.
CrossRefDirect Link - Mihailovic, G., J.R. Filpovic, M. Djurdjevic and Z. Dzeletovic, 1992. Effect of nitrogen form on maize response to drought stress. Plant Soil, 144: 191-197.
CrossRefDirect Link - Mulvaney, R.L., 1986. Comparison of procedures for reducing cross contamination during steam distillations in nitrogen-15 tracer research. Soil Sci. Soc. Am. J., 50: 92-96.
Direct Link - Schimel, J.P., L.E. Jackson and M.K. Firestone, 1989. Spatial and temporal effects on plant-microbial competition for inorganic nitrogen in a california annual grassland. Soil Biol. Biochem., 21: 1059-1066.
CrossRefDirect Link - Woods, L.E., C.V. Cole, L.K. Porter and D.C. Coleman, 1987. Transformations of added and indigenous nitrogen in gnotobiotic soil: A comment on the priming effect. Soil Biol. Biochem., 19: 673-678.
PubMedDirect Link