
Arturo Salame-Méndez
Department of Biology of Reproduction, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
LiveDNA: 52.32753
Alondra Castro-Campillo
Department of Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Jonathan Escobar-Flores
Graduated from the Master in Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Jesús Vergara-Huerta
Graduated from the Master in Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Héctor Serrano
Department of Health Sciences, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Dolores García-Suárez
Department of Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Jorge Haro-Castellanos
Department of Biology of Reproduction, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
Noé González-Ruiz
Department of Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
José Ramírez-Pulido
Department of Biology, Division of Biological and Health Sciences, Metropolitan Autonomous University, Iztapalapa Unit, Mexico
International Journal of Zoological Research
Year: 2020 | Volume: 16 | Issue: 2 | Page No.: 69-78







ABSTRACT
Background and Objective: The ecophysiological relationship between reproductive parameters and environmental variables is mostly unknown in wild temperate species of small rodents, inhabiting mid-latitude, temperate forests. Here the goal was the relationship between testicular function, testes and body weight to seasonal variations of daylight length and temperature in adult males of the Rock Mouse (Peromyscus difficilis felipensis), in a two-year study. Materials and Methods: Spermatogenesis, spermatogenic function, content and production of testosterone and body and testicular weight were monthly evaluated in free-living P. d. felipensis, using standard, histological and biochemical (ELISA) techniques. Results: Although the presence of sperm in both testes and epididymis was found all year round, there was a seasonal pattern with a spring peak in testis volume and testosterone content and production; though, the testicular function was maintained until fall and winter. Conversely, bodyweight decreased during spring-summer, while it increased in fall-winter. There was an ecophysiological positive correlation of reproductive parameters with environmental gathered data, especially with daylight length (i.e., increased correlation during spring, p<0.001) and there was a slight or no participation of environmental temperature. Conclusion: In addition to daylight length, the seasonal pattern of testicular function in these temperate populations of mice, indicates that other environmental factors, waiting to be elucidated (e.g., phytochemicals intake through food), also contribute to its regulation.
PDF Abstract XML References Citation
How to cite this article
Arturo Salame-Méndez, Alondra Castro-Campillo, Jonathan Escobar-Flores, Jesús Vergara-Huerta, Héctor Serrano, Dolores García-Suárez, Jorge Haro-Castellanos, Noé González-Ruiz and José Ramírez-Pulido, 2020. Ecophysiological Relationships of Environment and Reproduction in Males of Rock Mouse (Peromyscus difficilis felipensis) at a Temperate Forest. International Journal of Zoological Research, 16: 69-78.
DOI: 10.3923/ijzr.2020.69.78
URL: https://scialert.net/abstract/?doi=ijzr.2020.69.78
DOI: 10.3923/ijzr.2020.69.78
URL: https://scialert.net/abstract/?doi=ijzr.2020.69.78
INTRODUCTION
Free-living mammals and especially rodents usually have one or two annual reproductive seasons, showing hormonal periodical cyclic variation that correlates to gamete production and displays of specific copulatory behavior patterns1,2. Androgens, such as testosterone, promote several reproductive aspects, including male mating behavior3. In turn, specific environmental conditions of the habitat, such as temperature, daily light hours, food availability, pluvial regime and climate shifts affect these aspects4-8.
Testis volume, usually considered an indicator of sperm production in micro-mammals, is credited for both the individual sex performance and its potential reproductive success9. Testosterone plays a relevant role in testes growth, regulation of spermatogenetic division in meiosis and maintenance of spermatogenesis10,11. Testosterone production with its intra-gonadal and circulating concentration influences more general reproductive events; e.g., sex differentiation, mate selection and male behavior, together with reproductive success12-14.
Data on rodent testosterone contents, testis morphology, behavior and reproductive outcome correlations usually come from high latitudes species, especially from laboratory rodents, or from wild species raised under controlled environmental conditions6,15-19. Moreover, documentation of external somatic characteristics in wild free-living individuals is often associated with the time of year and used as an indicator of the population's reproductive status. In captured micromammals, these characteristics usually are i) amount of gestating or lactating females and ii) testis size and position in males20-22. While this information may describe the reproductive biology of the species at an ecological level, it is also necessary to understand fundamental phenomena that regulate physiological and endocrine processes in free-living rodents.
To contribute to ecophysiological knowledge from free-living rodents inhabiting midlatitude, temperate forests, we have been studying the reproductive biology and ecology (e.g., population ecology, diet, use of space) of two species of deer mice. One of them, a subspecies of the Rock Mouse (Peromyscus difficilis felipensis), is widely distributed on the wooded areas around Mexico City and those nearby in the State of Mexico, including coniferous and mixed forests23,24. In our study locations at the mountain ranges in western Mexico City, the reproductive activity of P. d. felipensis happens all year round but with two distinctive peaks, occurring during the spring and trough fall-winter, respectively25. The study aimed to examine whether there is a relationship between seasonal variations of two environmental conditions (daylight length, temperature) with testicular function (testis volume, spermatogenesis, production of testosterone and its intra-gonadal content) in male adults of P. d. felipensis inhabiting on two locations of these mid-latitude, temperate forests, along two years.
MATERIALS AND METHODS
Study areas: Cumbres del Ajusco National Park (CANP) is located in the southwestern part of Mexico Valley, within the Ajusco-Chichinautzin Mountain Range. Its climate belongs to the subhumid temperate type C(w1)(w)(b’)i’26. Higher temperatures range from April-September (mean±standard deviation, 15±2°C) and lower temperatures through November-February (11±1°C). Rainfalls occur along May-October (152±22 mm), while the dry season follows in November-April (16±11 mm). Vegetation corresponds to a stratified temperate mixed-forest with seasonal plants and tiller grasses. Oyamel trees (Abies religiosa) and oaks (Quercus sp.) dominate the canopy of mixed plots of other conifers and broad-leaved trees. The understory includes abundant bushes, mainly Senecio and Salix, together with seasonal, herbaceous asteraceae, rosaceae and solanaceae, among others and tillers of hard tall grasses (Muhlenbergia sp., Festuca sp.) or zacatonales. The ground includes mosses, lichens and several fungi. Sampling location was at 19°13'37" N, 99°15'37" W and 3180 m.
Desierto de los Leones National Park (DLNP), it is located at the western slope of Mexico Valley at Las Cruces Mountain Range, a prolongation of the Ajusco Mountain Range. Its climate belongs to the most subhumid temperate type C(w2)(w’)(b’)ig26. Higher temperatures occur in April-July (12.6±6°C), while lower temperatures are December-February (8.1±2°C). The rainy season is on May-August (235±30 mm) and the dry season is December-February (12±4 mm). Vegetation at DLNP is mostly as in CANP, but with a predominance of oyamel forests at higher elevations. Sampling location was at 19°18'17"N, 99°19'14"W and 2180-3200 m.
Capture and preparation of mice: We collected 43 adult males of P. difficilis felipensis, along two years (January, 2013 to December 2014), in aforementioned sampling locations. Mice were monthly captured, using Sherman traps (H.B. Sherman Traps Co, Tallahassee, FL, USA), baited with oat flakes, then transferred to the Laboratory of Mammals at the Division of Biological and Health Sciences (DBHS) and killed by cervical dislocation. All animal manipulations were made according to international standards27 and were approved by the Ethics Commission at DCBS, UAMI. We took standard measures (mm) and weight (g), before preparing corpses as skeleton voucher specimens20 to house them at UAMI Mammal Collection. Testes were surgically exposed and excised to measure its width and length conventionally (to the nearest 0.01 mm) and to calculate their volume using the geometric formula for a prolate spheroid28. Recognition of adult males followed Hoffmeister29, together with a typical pattern of pelage color for the species30.
Testosterone production: This and the next two following procedures were developed at the Laboratories of Animal Ecophysiology and Organic Synthesis, DBHS. We followed Salame-Méndez et al.31,32 with some modifications. Right testicle gonadal tissue was introduced in Eppendorf tubes containing Krebs-Ringer buffer with glucose (KRBG, pH 7) without metabolic cofactors and 10,000 cpm of tritiated cholesterol (1,2-3H specific activity-sa-47.7 Ci/mmol) (C273H) (Nuclear New England, Boston, MA). Experimental tubes (with tissues) and control tubes (without tissues), were incubated 1 h at 36°C. Metabolic reactions were stopped by freezing; the tubes were then defrosted and tissues in the incubation dissolution were homogenized. Two aliquots were taken from each homogenate, one to evaluate the precursor’s biotransformation (C273H) into testosterone and the other to determine protein concentrations33. Testosterone extraction was made from diethyl ether by duplicate. The organic phase was separated from the aqueous phase in a dry-ice:acetone media and the organic phases were evaporated to dryness. Extraction efficiency was made by adding to five tubes, at random, ≈1,000 cpm tritiated testosterone (1,2,6,7,16,17-3H) (sa 139 Ci/mmol) (Nuclear New England, Boston, MA); average extraction efficiency was 96.4±1.2% and results of each testosterone production were corrected, starting from percentage recovery. Each tube containing testosterone extracts was added diethyl ether:methanol (3:1, v/v) and samples were transferred to chromatoplates (20×20 cm, UV 250 nm radiation indicator, Merck). Testosterone extracts were separated by TLC, using (i) benzene:ethyl acetate (5:3, v/v) and (ii) benzene:methanol (90:10, v/v), as mobile phases. Ratio-to-front (Rf) reference testosterone corresponding areas, were visualized with a UV lamp. The silica was scraped and the androgen adsorbed to the silica was eluted with a mixture of diethyl ether:methanol (1:1, v/v); collected in glass vials and counted, after the addition of Instagel (Packard), in a liquid-scintillation spectrometer (Beckman, LS-7000) with maximum efficiency for tritium of 53%. Testosterone productions were expressed as a percentage of C273H biotransformation, related to 100 mg of gonad protein after 1 h incubation period.
Intra-gonadal testosterone content: Left testicle gonadal tissue was homogenized by sonication. The extraction of total sexual steroid contents in each homogenized was made by duplicated with diethyl ether. Evaluation of testosterone contents in gonadal tissue was made by enzyme-linked immunosorbent assay (ELISA), using commercial kits (DRG Instruments Inc.® Frauenberg, Marburg. Germany) and following instructions provided in manufacturer’s manual. Testosterone concentration was determined from a spectrophotometer (Microplate Reader, MR 600, Dynatech Product® Dynatech, Chantilly, VA. USA). The specificity of testosterone antiserum and linearity described by the kit manufacturer was validated by testing the solutions of the hormone provided in the kit, as well as from solutions of testosterone (T) and rostenedione (A) and progesterone (P4) (Sigma Labs, Santa Fe, NM, USA), previously purified. Percentage of cross-reactivity (specificity) was 100 for T; 0.8±1.23 for A and <0.1 for P4 and some of those reported by the manufacturer: 100 for T; 6.6 for 5α-dihydrotestosterone; <0.1 for dehydroepiandrosterone; 0.9 for A and <0.1 for P4. The sensitivity (minimum detectable concentration) of the assay for testosterone was 83 pg.
Spermatogenesis evaluation: Assessment of spermatogenesis was developed in the Laboratory of Animal Ecophysiology. A biopsy taken from each left testicle and both epididymis was then placed within a formalin-buffered solution and included in paraffin. Serial sections (3-5 μm) were processed for staining with hematoxylin-eosin. Spermatogenic and spermiogenic processes were determined following Fawcet34: Phase 1, or spermatocytogenesis, involves the development of spermatogonia until its differentiation into preleptotene spermatocytes; Phase 2 involves the meiotic process, taking place within spermatocytes, which differentiate into spermatids and Phase 3 or spermiogenesis, corresponds to the differentiation of spermatids into spermatozoids.
Environmental conditions: This and the next procedures were carried out at the Laboratory of Mammals, DBHS. Monthly average data for daylight length and temperature were obtained from the nearest meteorological station (Ecoguardas Ajusco (EAJ) 19.27°N, -99.20°W, 2584 m) and TUTIEMPO Network (https://www.tutiempo.net) during the study period.
Statistical analysis: Analysis of variance (ANOVA), followed by Tukey's multiple comparisons, were used to examine average behavior of testicular function (testicular volume, testosterone production, intra-gonadal testosterone content) and individual weight by age, location, month and season of the year (data pooled in trimesters; e.g., March-May into spring and so forth). To establish monthly and seasonal behavior of individual weight, testicular volume variation was eliminated through a covariance analysis (ANCOVA). Linear correlations between individual weight, testis volume, testosterone production and testosterone intragonadal content were analyzed through simple linear regressions. Whereas dependence of testicular function (testis volume, testosterone production and testosterone intra-gonadal content) to environmental conditions (daylight length and temperature) were obtained through a Pearson’s multiple comparison matrix. All analysis were performed with the statistical packages GraphPrisma®35 and NCSS36, under a significance level of α<0.05.
RESULTS
Figure 1a shows monthly testicular function in adult males of Peromyscus difficilis felipensis.
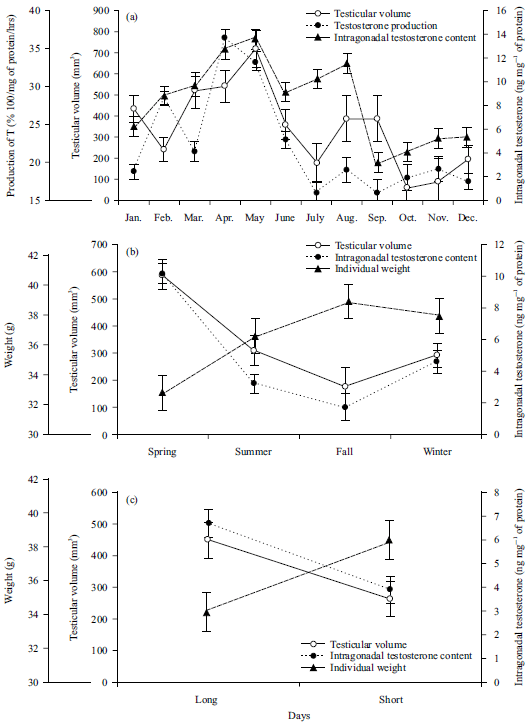 | |
| Fig. 1(a-c): | Temporal variations of testicular function (testicular volume, testosterone production and intragonadal testosterone content) and individual weight in adult males of Peromyscus difficilis felipensis, together with their dependence on daylight length: (a) Monthly variation of testicular function, (b) Seasonal variation of testicular function and (c) Dependent patterns of testicular function, according to daylight length |
| Table 1: | Seasonal means±standard deviations for somatic and physiological variables in adult males of Peromyscus difficilis felipensis and for environmental variables |
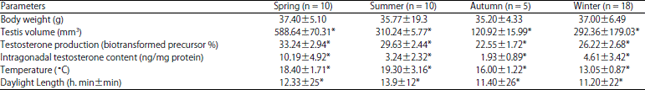 | |
| *: Significant statistical differences, α<0.05 | |
Testosterone production (Fig. 1a) was higher in April, May and August (F = 24.54, df = 11, p<0.0001), whereas its lowest levels were found from September through November (F = 24.54, df = 11, p<0.0001). Intra-gonadal testosterone content (Fig. 1a) starts at a basal low level in January (2.425±0.614), increases through February-March to reach a peak by April-May (13.653±0.752, F = 4.93, df = 11, p = 0.00019) and decreases from June to December, where basal levels are maintained at almost constant values (3.25±1.37) for all three seasons. Testis volume (Fig. 1a) was statistically higher during May (F = 30.82, df = 11, p<0.0001) than in every other month, even when slight variations were found all year long (F = 1.51, df = 11, p = 0.1772); then, from June through February, testis volume was statistically lower (F = 30.82, df = 11, p<0.0001).
Table 1 and Fig. 1b show seasonal changes of testicular function and weight, the former also shows seasonal changes in daylight length and temperature. Testosterone production (intra-gonadal testosterone contents, F = 10.42, df = 3, p<0.0001) and testis volume (F = 8.89, df = 3, p = 0.0001) were significantly higher during spring. A significant decrease for testosterone production was noted from summer through fall (F = 24.54, df = 11, p<0.0001) but with an increase by winter (Fig. 1b). Intra-gonadal testosterone contents was significantly lower in fall and mostly variable during summer or winter (F = 4.93, df = 11, p = 0.0002). Although no significant variations in body weight were recorded along the year (F = 2.64, df = 3, p = 0.0631, Table 1), it was higher during the shorter day seasons (fall-winter, Fig. 1b, c).
Pearson’s correlation analysis showed no correlation between daylight length and testis volume (r = 0.28, p = 0.06), whereas daylight length and environmental temperature (Fig. 1c) had a significant positive correlation with the synthesis of testosterone (r = 0.57, p<0.0001) and with intra-gonadal content of the androgen (r = 0.32, p = 0.03). In fact, dependence was higher during larger daylight length through spring-summer, than during shorter daylight length in fall-winter (testosterone production: F = 41.21, df = 1, p<0.0001; intragonadal testosterone content: F = 9.40, df = 1, p = 0.0037; testis volume: F = 4.85, df = 1, p = 0.0331). Conversely, individual weight was significantly lower during longer daylight in spring-summer (F = 41.21, df = 1, p<0.0001; Fig. 1c).
Spermatogenesis in Peromyscus difficilis felipensis was as follows. Spermiogenic processes (Phase 3) were recorded from testicular tubules in adults captured in January, whereas spermatocyte to preleptotenic spermatid differentiation (Phase 2) was the predominant phase in the germinative epithelia in testes from individuals captured during February, with an alternating predominant pattern. We also observed abundant mature spermatozoa in epididymis from animals captured all year long. Therefore, when taken together, these data indicate that the spermatogenic cycle lasts 34±4 days in P. difficilis felipensis.
DISCUSSION
Although spermatogenesis occurred all year round in males of Peromyscus difficilis felipensis, evidences showed a seasonal pattern with a spring peak for other testicular functions and body weight. Adult males of Peromyscus difficilis felipensis had sperm in both testicles and epididymis throughout the year, indicating a continuous active and enduring gamete production (34±4 days), which is consistent with the continuous production and intra-gonadal content of testosterone, the androgen that regulates meiosis during spermatogenesis and maintains spermiogenesis11. On the other hand, together with higher spring values for testis volume, testosterone production and intra-gonadal testosterone content indicate that this is the preferred reproductive season for P. d. felipensis, especially from April to May (Table 1). Supporting this fact are the largest numbers of pregnant and/or lactating females also recorded during late spring and in summer25. This supports that favoring of reproductive activity by P. difficilis in spring is related to days with longer light hours that precede and occur in the summer rainy season and, thus, promote a more diverse understory (i.e., more food availability) in both National Parks.
On the other hand, one verified fact was that position of testes was not always associated to reproductive activity of males of P. d. felipensis. In some mice with abdominal testes, both production and intra-gonadal content of testosterone were similar to those in mice with scrotal testes; the former also presented spermatogenesis in phases 2 and 3, as well as gametes in the epididymis all year round. Other cricetids studied under either natural or controlled conditions show similar evidence. We found the same pattern in free-living populations of the congeneric syntopic species Peromyscus melanotis25,28,37 in our study areas, while Boiani et al.38 reports this fact in Oligoryzomys flavescens from a temperate boreal habitat. Moreover, in a laboratory colony of Neotomodon alstoni, Olivera et al.39 found reproductively active males, even though they had no scrotal testicles. Therefore, inferring reproductive activity just from the sole observation of scrotal testes might prove to be inaccurate for some species of wild micromammals.
There was a positive dependence of the reproductive physiological variables to daylight length. Even though the correlations between daylight length with the production of testosterone and its intra-gonadal content were both low in P. d. felipensis, there is a positive dependence of the physiological variables to this abiotic one as noted elsewhere40. This is reinforced by the fact that testes of mice from the seasons with longer light hours per day (spring-summer), showed significantly higher synthesis and contents of testosterone, as compared to mice from shorter daylights in fall-winter. Therefore, we can suggest that there is a positive dependence between both environmental conditions, but especially with daylight length (Fig. 1c) and testicular function (testosterone production and intra-gonadal testosterone content). Taken together, our results agree with some studies of wild rodents under laboratory conditions, where circulating testosterone concentration increased or decreased in response to light time7,41-43.
On the other hand, the correlation between testicular volume and both testosterone production and intra-testicular content, suggests that testicular function could be rather stimulated by both abiotic (e.g., light hours) and biotic environmental factors (e.g., type of food), which interact with the species genetic reproductive background. That is, although the highest reproductive peak for this species occurred during spring, reproductive activity was recorded until summer, as well as in fall-winter, even though testosterone production, intra-testicular testosterone content and testis size decreased. Moreover, our records of spermatozoa in the epididymis, agree with our results from a previous study25: these indicators for testicular function also went down but did not cease through apparently less favorable conditions for the reproductive activity, such as the colder and dryer seasons of autumn and winter. Also, seasonal dependence between spermatogenesis and testicular volume with testosterone production and intra-testicular testosterone content, allowed us to verify that these indicators increase during the seasons with longer light hours (spring-summer), while they diminish, though without ceasing, during those days with shorter light hours (fall-winter).
The mild hardening of environmental conditions of the studied temperate, mid-latitude forests, are not the main regulating factor of the reproductive activity in males of P. d. felipensis. Young et al.5 have shown under laboratory conditions that a combination of shorter daylight length, lower temperature and food restriction, reduced testis volume by apoptosis in P. maniculatus. Our results with free-living P. difficilis do not support the above since even though the temperature is lowered and daylight is shorter in fall-winter, both plant and insect food items are still available at these mid-latitude, temperate forests44,45. In such conditions, even though testicular size decreases, both testosterone production and spermatogenesis do not cease25; in fact, here we observed no apoptosis processes in the testicles and epididymis analyzed. Therefore, one alternative option is that both production of testosterone and of spermatozoa will decrease rather due to the diminishment of plant secondary compounds (phytoestrogens, FTE) or fungi-derived compounds (mycoestrogens, MCE), during food intake46-48, when these food resources become scarcer o disapear.
Both FTE and MCE can mimic mammal estrogens and, thus depending on their amounts, they could deplete or enhance reproductive physiology in wild mammals49,50. At high quantities, they can disrupt normal on-off controls of steroid hormones in herbivorous mammals, thus inhibiting their non-genomic and genomic effects51-53. On the other hand, at lower amounts, FTE and MCE could trigger reproductive physiological processes for mating and reproduction54. Therefore, FTE and MCE could regulate reproductive activity in wild herbivores55,56. Accordingly, there is a relation between the reproductive activity of free-living, rodent species and the time of year when food is abundant and thus of FTE and MCE contents. For instance, Jameson Jr. and Peeters57 reported an increase of reproductive activity in heteromyid mice from a desert: during the summer rains, coinciding with the rise of plants, mice consumed more phytochemicals (e.g., phytoestrogen), while with the reduction of such food resources, during the dry season, mice underwent a delayed and decreased reproductive activity. Kaneko58 found that litter size decreased (36.7%) in a population of Japanese voles (Microtus montebelli) inhabiting a plantation of cypresses, thereof containing a high concentration of phytoestrogens, as compared to another population inhabiting a fallow rice field. For its part, Berger et al.59 found delayed oogenesis and decreased uterine size in females of Microtus montanus, due to compounds contained in plants eaten during late summer, especially during the dry years; therefore the reproductive activity of this species may be also affected by phytochemicals present in plants they consume60. In our study area, it is suggestive that P. d. felipensis and its syntopic congeneric, P. melanotis, eat a high amount of fungi together with a larger diversity of plants during the rainy season44,45, which might be stores of MCE and FTE that might be acting as triggers for breeding. Nevertheless, further studies are needed to elucidate whether FTE and MCE in food have any roll on reproduction in our model Peromyscus, as well as in another species of wild micromammals61,62.
Also interestingly is the fact that bodyweight, deprived of testicular volume, was significantly reduced in adult males of P. d. felipensis during spring, the season with higher reproductive activity (Fig. 1b). Conversely, body weight increased during seasons with shorter days (fall-winter) and lower temperatures (Fig. 1b,c). Low weight may be due to high energy expenditures, together with physiological distress, caused by the increase of allostatic overload during reproductive events63; on the other hand, its subsequent increase, during fall-winter, might be due to an increase of metabolic rate during the cold seasons, to withstand the cold. We have found, indeed, that triglyceride content increases during colder seasons (autumn-winter) in free-living Peromyscus melanotis and P. difficilis64. Furthermore, the diet composition of adult males of P. d. felipensis, the intake of a lower amount of plant species is completed by arthropod consumption during the “harsher” environmental conditions44,45. The above could allow this species to gain weight (e.g., muscle mass and fat tissue) during the autumn-winter to prepare it for the allostatic overload derived from the search and copulation with mates65, as well as for the defense of the mating territory, as has been reported for other wild mice8,66-68.
Finally, we have shown here that male reproductive activity of free-living P. difficilis felipensis is affected by coincident internal and external conditions that then determine individual reproductive success. Constant sperm in testis tubules and the epididymis, together with adult male testosterone production and intra-gonadal testosterone content, associated with daylight and temperature oscillations, are only part of the population maintenance and perpetuation. However, other involved environmental factors also wait to be evaluated, to establish their role in the reproductive dynamics of free-living, herbivore mice such as P. d. felipensis. One is the obvious need for documentation on the basic female reproductive physiology. Another aspect of particularly important interest is the study of potential sex hormone-like acting compounds (e.g., FTE and MCE) within their diet, for these food compounds could be affecting the whole population's physiological reproductive cycle and might ultimately regulate perpetuation of P. d. felipensis, as well as that of other rodents sharing this mid-latitude, temperate forest habitat.
CONCLUSION
In adult males of Peromyscus difficilis felipensis from mid-latitude, temperate forests, spermatogenesis occurred all year round, but their testis function (testis volume, testosterone production and intra-gonadal testosterone content) and reproductive peak were enhanced by longer day length and higher temperatures. Fluctuations of body weight (decrease in spring-summer; increase in fall-winter) were also related to its reproductive pattern. Rather than physical variables, in habitats ruled by the rainy seasons, the more food availability and diversity (e.g., more plants and fungi food items) associated with contents and amount of plant and fungi chemicals, might promote such patterns.
SIGNIFICANCE STATEMENT
To avoid underestimations of the reproductive activity in the population of wild, adult male mice with a retractile testis, it is necessary to verify that their size and position agree with other physiological parameters such as the production and contents of sex hormones. In addition to abiotic factors such as daylight length and temperature, the testicular function of mid-latitude, temperate mice may be being triggered and sustained by bioactive compounds consumed in plant and fungi food items.
ACKNOWLEDGMENTS
Our appreciation to the late Juan Patiño Rodríguez, technical collector and taxidermist at UAMI Mammal Collection, for his invaluable fieldwork and cabinet assistance. M. S. Joaquín Herrera Muñoz gave technical aid for evaluating testosterone production. We also thank the several undergraduate and graduate students that participated during fieldwork. This study was partially supported by the Division of Biological and Health Sciences in UAMI (DCBS-144.03.07 to ASM and DCBS-143.02.46 to ACC), by the National Council of Science and Technology (CONACyT-1253-9203 y 400200-5R29117N to JRP) and by the Public Education Secretariat (SEP 94-01-00-002-247 to JRP).
REFERENCES
- Van Tienhoven, A., 1983. Reproductive Physiology of Vertebrates. 2nd Edn., Cornell University Press, Ithaca, NY, USA, Pages: 491.
Direct Link - Bronson, F.H., 1989. Mammalian Reproductive Biology. University of Chicago Press, Chicago, IL, USA, Pages: 325.
Direct Link - Nelson, R.J., 2011. An Introduction to Behavioral Endocrinology. 4th Edn., Sinauer Associates, Inc., Sunderland, MA, USA, Pages: 712.
Direct Link - Sadleir, R.M.F.S., 1969. The Ecology of Reproduction in Wild and Domestic Mammals. Methuen & Co., Ltd., London, UK, Pages: 321.
Direct Link - Young, K.A., B.R. Zirkin and R.J. Nelson, 2000. Testicular regression in response to food restriction and short photoperiod in white-footed mice (Peromyscus leucopus) is mediated by apoptosis. Biol. Reprod., 62: 347-354.
CrossRefDirect Link - Trainor, B.C., L.B. Martin, K.M. Greiwe, J.R. Kuhlman and R.J. Nelson, 2006. Social and photoperiod effects on reproduction in five species of Peromyscus. General Comp. Endocrinol., 148: 252-259.
CrossRefDirect Link - Bronson, F.H., 2009. Climate change and seasonal reproduction in mammals. Phil. Trans. R. Soc. B, 364: 3331-3340.
CrossRefDirect Link - Ochiai, K., K. Susaki, T. Mochizuki, Y. Okasaka and Y. Yamada, 2010. Relationships among habitat quality, home range size, reproductive performance and population density: Comparison of three populations of the Japanese serow (Capricornis crispus). Mammal Study, 35: 265-276.
CrossRefDirect Link - Cortés-Calva, P. and S.T. Álvarez-Castañeda, 1999. Gonadal size of males of Chaetodipus arenarius (Rodentia: Heteromyidae) during a reproductive cycle in Baja California Sur, Mexico. Rev. Biol. Trop., 47: 617-622.
Direct Link - Schulte-Hostedde, A.I., J.S. Millar and G.J. Hickling, 2003. Intraspecific variation in testis size of small mammals: implications for muscle mass. Can. J. Zool., 81: 591-595.
CrossRefDirect Link - Walker, W.H., 2011. Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis, 1: 116-120.
CrossRefDirect Link - Mooradian, A.D., J.E. Morley and S.G. Korenman, 1987. Biological actions of androgens. Endocrine Rev., 8: 1-28.
CrossRefDirect Link - Norris, D.O. and K.H. Lopez, 2011. Hormones and Reproduction of Vertebrates. 1st Edn., Academic Press, Bulington, MA, USA, Pages: 400.
Direct Link - Mani, S., P. Mermelstein, M. Tetel and G. Anesetti, 2012. Convergence of multiple mechanisms of steroid hormone action. Horm. Metab. Res., 44: 569-576.
CrossRefDirect Link - Ojeda, S.R. and H.F. Urbanski, 1994. Puberty in the rat. In: The Physiology of Reproduction, Knobil, E. and J.D. Neil, Raven Press, Chicago, IL, USA pp: 363-410.
Direct Link - Meites, J., 1995. Neuro-endocrine control of reproduction in aging rats and humans. In: The Reproductive Neuroendocrinology of Aging and Drug Abuse, Dipak, K.S. and C.D. Barnes, CRC Press, Boca Raton, FL, USA., pp: 109-117.
Direct Link - Perrot-Sinal, T.S., M. Kavaliers and K.P. Ossenkopp, 1998. Spatial learning and hippocampal volume in male deer mice: relations to age, testosterone and adrenal gland weight. Neuroscience, 86: 1089-1099.
CrossRefDirect Link - Trainor, B.C. and C.A. Marler, 2001. Testosterone, paternal behavior, and aggression in the monogamous California mouse (Peromyscus californicus). Horm. Behav., 40: 32-42.
CrossRefDirect Link - Couto, D. and S.A. Talamoni, 2005. Reproductive condition of Akodon montensis Thomas and Bolomys lasiurus (Lund) (Rodentia, Muridae) based on histological and histometric analyses of testes and external characteristics of gonads. Acta Zool. (Stockholm), 86: 111-118.
CrossRefDirect Link - Ramírez-Pulido, J., I. Lira, S. Gaona, C. Müdespacher and A. Castro-Campillo, 1989. Management and Maintenance of Mammal Collections. Metropolitan Autonomous University, Mexico, Pages: 127.
Direct Link - Kunz, T.H., C. Wemmer and V. Hayssen, 1996. Sex, age and reproduction. In: Measuring and Monitoring Biological Diversity: Standard Methods for Mammals, Wilson, D.E., F.R. Cole, J.D. Nichols, R. Pudran and M.S. Foster, Smithsonian Institution Press, Washington D.C., USA, pp 279-290.
Direct Link - Romero-Almaraz, M.L., C. Sánchez-Hernández, C. García-Estrada and R.D. Owen, 2007. Small Mammals: Manual of Techniques for Capture, Preparation, Preservation and Study. 2nd Edn., National Autonomous University of Mexico, Mexico, Pages: 202.
Direct Link - Hoffmeister, D.F., L. De La Torre, 1961. Geographic variation in the mouse Peromyscus difficilis. J. Mammal., 42: 1-13.
CrossRefDirect Link - Chavez-T., J.C. and G. Ceballos, 2014. Peromyscus difficilis (J.A. Allen, 1891), Southern Rock Deermouse. In: Mammals of Mexico, Ceballos, G., Johns Hopkins University Press, Baltimore, MD, USA, pp. 351-352.
Direct Link - Salame-Méndez, A., A. Castro-Campillo, R.M. Vigueras-Villaseñor, J. Herrera-Muñoz, H. Serrano and J. Ramírez-Pulido, 2008. Production of testosterone in the testes of two species of Peromyscus (Rodentia: Muridae) during lowered sexual activity. In: Advances in the Study of the Mammals of Mexico, Lorenzo, C., E. Espinoza and J. Ortega, Mexican Association of Mammalogy (AMMAC), Mexico, pp: 311-321.
Direct Link - García, E., 2004. Modificaction to the Climatic Classification System of Köpen. 5th Edn., National Autonomous University of Mexico, Mexico, Pages 91.
Direct Link - NRC., 2011. Guide for the Care and Use of Laboratory Animals. 8th Edn., National Academies Press, Washington, DC., USA, ISBN-13: 9780309154000, Pages: 246.
Direct Link - Castro-Campillo, A., L. León-Altamirano, J. Herrera-Muñoz, I. Salgado-Ugarte, E. Mendieta-Márquez et al., 2012. Is there a difference between testosterone contents in two populations of the Black Eared Mouse, living under similar conditions but with differences in population patterns? Acta Zool. Mex., 28: 525-539.
CrossRefDirect Link - Castro-Campillo, A., A. Salame-Méndez, J. Vergara-Huerta, A. Castillo and J. Ramírez-Pulido, 2008. Fluctuations of land micro-mammals in temperate forests adjacent México City, Federal District. In: Advances in the Study of the Mammals of Mexico, Lorenzo, C., E. Espinosa and J. Ortega, AMMAC Special Publications II, Mexico, pp: 391-410.
Direct Link - Salame-Méndez, A., A. Castro-Campillo, E. Mendieta-Márquez, I.H. Salgado-Ugarte, J. Herrera-Muñoz and J. Ramírez-Pulido, 2004. Seasonal assessment of sexual steroid production in the testes of the Black-eared Mouse (Peromyscus melanotis Allen and Chapman, 1897) of different age classes. Acta Zool. Mex., 20: 103-114.
Direct Link - Salame-Méndez, A., E. Mendieta-Márquez, J. Herrera-Muñoz, A. Castro-Campillo and J. Ramírez-Pulido, 2005. Production assessment of steroid hormones in testes and ovaries of the Rock Mouse (Peromyscus difficilis felipensis). Rev. Soc. Mex. Hist. Nat., 2: 200-206.
Direct Link - Groves, W.E., F.C. Davis Jr. and B.H. Sells, 1968. Spectrophotometric determination of microgram quantities of protein without nucleic acid interference. Anal. Biochem., 22: 195-210.
CrossRefDirect Link - Fawcett, D.W., 1989. Bloom-Fawcett: Histology Treatise. 11th Edn., New Interamerican Editorial, Mexico, Pages: 1026.
Direct Link - Motulsky, H., 1999. Analyzing Data with GraphPad Prism. Graph Pad Software Inc., San Diego, CA., USA., Pages: 379.
Direct Link - Salame-Méndez, A., J. Herrera-Muñoz, R.M. Vigueras-Villaseñor, A. Castro-Campillo, A., E. Mendieta-Márquez and J. Ramírez-Pulido, 2005. Description of the ontogenetic profile of sexual steroid hormones and intermediaries in testes of the Black-eared Mouse (Peromyscus melanotis, Allen y Chapman, 1897). Rev. Soc. Mex. Hist. Nat., 2: 193-199.
Direct Link - Boiani, L., N. Berois and G. D'Elia, 2008. Annual male reproductive cycle of a Hantavirus reservoir, the long-tailed mouse Oligoryzomys flavescens (Rodentia; Cricetidae, Sigmodontinae) from Uruguay. Mastozool. Neotrop., 15: 23-32.
Direct Link - Olivera, J., J. Ramírez-Pulido and L.S. Williams, 1986. Reproduction of Peromyscus (Neotomodon) alstoni (Mammalia: Muridae) in laboratory conditions. Acta Zool. Mex., 16: 1-27.
CrossRefDirect Link - Mukaka, M.M., 2012. Statistics corner: A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J., 24: 69-71.
PubMedDirect Link - Demas, G.E. and R.J. Nelson, 1998. Photoperiod, ambient temperature, and food availability interact to affect reproductive and immune function in adult male deer mice (Peromyscus maniculatus). J. Biol. Rhythms, 13: 253-262.
CrossRefDirect Link - Pyter, L.M., Z.M. Weil and R.J. Nelson, 2005. Latitude affects photoperiod-induced changes in immune response in meadow voles (Microtus pennsylvanicus). Can. J. Zool., 83: 1271-1278.
CrossRefDirect Link - Pyter, L.M., B. Trainor and R.J. Nelson, 2006. Testosterone and photoperiod interact to affect spatial learning and memory in adult male white-footed mice (Peromyscus leucopus). Eur. J. Neurosci., 23: 3056-3062.
CrossRefDirect Link - Peralta-Juárez C.M., A. Castro-Campillo, M.E. Fraile-Ortega, A. Salame-Méndez, De La I.M. Cruz-Argüello, J.L. Patiño-Ortega, and J. Ramírez-Pulido, 2016. Seasonal diversity of diet contents in Peromyscus difficilis felipensis (Rodentia: Cricetidae), according to its reproductive stage. Proceedings of the 63rd Annual Meeting of The Southwestern Association of Naturalists (SWAN), April 14-17, 2016, SWAN & IBUNAM, pp: 70-71.
Direct Link - Peralta-Juárez, C.M., A. Castro-Campillo, M.E. Fraile-Ortega, A. Salame-Méndez and I.M. DelaCruz-Argüello et al., 2016. Diet of Peromyscus difficilis felipensis (Rodentia: Cricetidae) and its temporal intra-population variation in a mixed temperate forest of Mexico City. Proceedings of the XIII National Congress of Mammalogy, October 24-28, 2016, AMMAC & UNICACH.
Direct Link - Labov, J.B., 1997. Phytoestrogens and mammalian reproduction. Comparative Biochem. Physiol. Part A: Physiol., 57: 3-9.
CrossRefDirect Link - Kaldas, R.S. and C.L. Hughes, 1989. Reproductive and general metabolic effects of phytoestrogens in mammals. Reprod. Toxicol., 3: 81-89.
CrossRefDirect Link - Bernhoft, A., 2010. Bioactive Compounds in Plants—Benefits and Risks for Man and Animals. The Norwegian Academy of Science and Letters, Oslo, Norway, Pages: 253.
Direct Link - Forbey, J.S., A.L. Harvey, M.A. Huffman, F.D. Provenza, R. Sullivan and D. Tasdemir, 2009. Exploitation of secondary metabolites by animals: A response to homeostatic challenges. Integr. Comp. Biol., 49: 314-328.
CrossRefDirect Link - Wocławek-Potocka, I., C. Mannelli, D. Boruszewska, I. Kowalczyk-Zieba, T. Waśniewski and D.J. Skarżyński, 2013. Diverse effects of phytoestrogens on the reproductive performance: cow as a model. Int. J. Endocrinol., 2013: 1-15.
CrossRefDirect Link - Rosselli, M., K. Reinhart, B. Imthurn, P.J. Keller and R.K. Dubey, 2000. Cellular and biochemical mechanisms by which environmental oestrogens influence reproductive function. Human Reprod. Update, 6: 332-350.
CrossRefDirect Link - Nilsson, S., S. Mäkelä, E. Treuter, M. Tujague and J. Thomsen et al., 2001. Mechanisms of estrogen action. Physiol. Rev., 81: 1535-1565.
CrossRefDirect Link - Glover, A. and S.J. Assinder, 2006. Acute exposure of adult male rats to dietary phytoestrogens reduces fecundity and alters epididymal steroid hormone receptor expression. J. Endocrinol., 189: 565-573.
CrossRefDirect Link - Rochester, J.R. and J.R. Millam, 2009. Phytoestrogens and avian reproduction: Exploring the evolution and function of phytoestrogens and possible role of plant compounds in the breeding ecology of wild birds. Comparative Biochem. Physiol. Part A: Mol. Integ. Physiol., 154: 279-288.
CrossRefDirect Link - Hughes, C.L., Jr., 1988. Phytochemical mimicry of reproductive hormones and modulation of herbivore fertility by phytoestrogens. Environ. Health Perspect., 78: 171-174.
CrossRefPubMedDirect Link - Wynne-Edwards, K.E., 2001. Evolutionary biology of plant defenses against herbivory and their predictive implications for endocrine disruptor susceptibility in vertebrates. Environ. Health Perspect., 109: 443-448.
CrossRefDirect Link - Jameson, E.W. Jr. and H.J. Peeters, 2004. Mammals of California. University of California Press, Berkeley and Los Angeles, CA, USA, Pages: 429.
Direct Link - Kaneko, Y., 1978. A comparison of the number of embryos and measurements of Microtus montebelli in two types of habitats. Acta Theriol., 23: 140-143.
Direct Link - Berger, P.J., E.H. Sanders, P.D. Gardner and N.C. Negus, 1977. Phenolic plant compounds functioning as reproductive inhibitors in Microtus montanus. Science, 195: 575-577.
Direct Link - Negus, N.C., P.J. Berger and L.G. Forslund, 1997. Reproductive Strategy of Microtus montanus. J. Mammal., 58: 347-353.
CrossRefDirect Link - Castro-Campillo A., A. Salame-Méndez, J. Haro-Castellanos, M.A. Briones-Salas and H. Serrano et al., 2015. Presence of ME [mycoestrogens] and PE [phytoestrogens] related to environmental temporality and reproductive activity in two wild micro-mammals. Proceedings of the XIII Meeting Participation of Women in Science, August 17-19, 2015 CIO, UDG,.
Direct Link - Salame-Méndez A., J. Haro-Castellanos, A. Castro-Campillo, M.D. García-Suárez and H. Serrano et al., 2016. Spatio-temporal analysis of phytochemicals in faeces of micro-mammals. Proceedings of the 63rd Annual Meeting of The Southwestern Association of Naturalists (SWAN), April 14-17, 2016 SWAN & IBUNAM, pp: 17-73.
Direct Link - Wingfield, J.C., 2005. The concept of allostasis: coping with a capricious environment. J. Mammal., 86: 248-254.
CrossRefDirect Link - Vergara-Huerta J., J. Ramírez-Pulido, A. Castro-Campillo, F. Méndez-DelaCruz and I.H. Salgado-Ugarte et al., 2011. Implications of metabolism in the abundance of two species of Peromyscus (Rodentia: Muridae). Proceedings of the III Mexican Congress of Ecology, April 3-7, 2011, Scientific Mexican Society of Ecology, pp: 1-304.
Direct Link - Peralta-Juárez, C.M., A. Castro-Campillo, M.E. Fraile-Ortega, A. Salame-Méndez and DelaCruz-Argüello et al., 2016. Differential diet in Peromyscus difficilis felipensis (Rodentia: Cricetidae), according to its reproductive stages in a mixed, temperate forest. Proceedings of the XIII National Congress of Mammalogy, October 24-28, 2016, AMMAC & UNICACH.
- Koivula, M., E. Koskela, T. Mappes and T.A. Oksanen, 2003. Cost of reproduction in the wild: manipulation of reproductive effort in the bank vole. Ecology, 84: 398-405.
CrossRefDirect Link - Waterman, J., 2007. Male mating strategies in rodents. In: Rodent Societies: An Ecological and Evolutionary Perspective, Wolff, J.O. and P.W. Sherman, The University of Chicago Press, Chicago, IL, USA, pp 27-41.
Direct Link - Reid, R.E.B., E.N. Greenwald, Y. Wang and C.C. Wilmers, 2013. Dietary niche partitioning by sympatric Peromyscus boylii and P. californicus in a mixed evergreen forest. J. Mammal, 94: 1248-1257.
CrossRefDirect Link