
Srinivasa Kalyan Chadamala
Yogi Vemana University, Kadapa, Andhra Pradesh, India
Anu Prasanna Vankara
Yogi Vemana University, Kadapa, Andhra Pradesh, India
LiveDNA: 91.16872
International Journal of Zoological Research
Year: 2019 | Volume: 15 | Issue: 2 | Page No.: 43-53







ABSTRACT
Background and Objectives: Parasitic communities of Rhizoprionodon acutus were investigated from Nellore Coast (14.43°N 79.97°E), Bay of Bengal during January, 2014 and December, 2015. Special focus was given to helminth infracommunities and effect of host size and sex on its structure. Material and Methods: The parasites obtained from the host samples were enumerated and preserved for morphological analysis and ecological study using standard protocols. Results: A total of 11 taxa including 3 ectoparasites and 8 endoparasites were identified. Only, 59.2% of the fish were parasitized by one or more than one parasitic species, with an average of 2 parasites/fish. Endoparasites represented 86% of the total parasites collected with cestodes overwhelming the other parasites within the parasitic community. The number of ecto and endoparasites species per individual host ranged from 1-15 and 1-17, respectively while the mean numbers of parasite specimens per individual host were 0.38 and 1.8, respectively. There were no core and secondary species in the community and all the parasitic species occupied the rank of satellite species. Relationship between host size and parasitization revealed that medium sized (Group 2) fishes were more infected than other classes of fishes. The aggregated or over-dispersed nature of parasitic infection was exhibited by only P. floraformis (5.12), K. minuta (4.50), L. sturionis (15.1) and L. secundus (6.41). Considerable interspecific associations were shown by only those parasites occupying the same niche i.e., copepod and monogenean in gills and different species of cestodes in spiral intestine of the host. Though, there is no significant relation between host sex and overall parasitization in the host but individually, only, L. secundus, H. peridareus, N. lingualis and larva of Hysterothylacium sp. showed a significant relation between host sex and parasitization. Conclusion: The communities of metazoan parasites in R. acutus are depauperate and are close to the isolationist extremity of the continuum with only 11 species.
PDF Abstract XML References Citation
Received: June 03, 2019;
Accepted: August 19, 2019;
Published: September 03, 2019
How to cite this article
Srinivasa Kalyan Chadamala and Anu Prasanna Vankara, 2019. Metazoan Parasite Infracommunities of Rhizoprionodon acutus from Nellore Coast off Bay of Bengal Andhra Pradesh, India. International Journal of Zoological Research, 15: 43-53.
DOI: 10.3923/ijzr.2019.43.53
URL: https://scialert.net/abstract/?doi=ijzr.2019.43.53
DOI: 10.3923/ijzr.2019.43.53
URL: https://scialert.net/abstract/?doi=ijzr.2019.43.53
INTRODUCTION
Rhizoprionodon acutus Rüppell 1837 most commonly known as Fish-shark, Longman's Shark, White-eyed Shark and vernacularly known as ‘Kukkasorrah’ and ‘Palasorrah’ in Southern India specially Andhra Pradesh is the most abundant shark of the Bay of Bengal. It is a familiar continental shelf species with typical coloring of grey on top and white underneath and born with 25-40 cm in length, maturing at 75 cm and reaching a maximum size of 100-178 cm 1. It is a coastal species, which is generally observed in a broad range of artisanal, survival and profitable fisheries and recurrently spotted in fish markets. Despite its common occurrence in the Bay of Bengal, quantitative helminthological studies of R. acutus have been focused so far on particular parasite taxa and limited to sporadic examination of incomplete individual hosts2-9. The parasitic communities in the fish may be subjective to both host related factors (i.e., size, sex, diet, vagility and migratory habits) and habitat related factors10-15. There are only a very few studies which were focused on the parasite assemblages in the spiral intestines of sharks16-18. Very few scientists all over the world had focused on the studies on the parasitic community structure in sharks14,15,19. But, still there is a wide lacuna on the studies on the entire metazoan parasites of the sharks from India. Kalyan et al.20 carried out pioneering work on the population and seasonal dynamics of the metazoan parasites from R. acutus off Nellore coast Bay of Bengal as the first aspect of community ecology and the second aspect of community ecology i.e., study of parasite community structure at both the component and infracommunity levels and their relationship with the size and sex of the host with parasitic infection and their interactions is highlighted in the present study.
MATERIAL AND METHODS
Study area: The present study was performed in Nellore Coast (14.43°N 79.97°E) Andhra Pradesh, Bay of Bengal (Fig. 1) during January, 2014-December, 2015 to investigate the metazoan parasite infracommunities of the Milk shark, R. acutus.
Fish sampling and Parasitological examination: A total of 152 hosts of various sizes i.e., small, medium and large collected from local fishermen were transported to the laboratory for further parasitological examination. Corporeal parameters such as length, weight, age and gender of each specimen were recorded for ecological studies according to standard procedure. R. acutus measured 18-65 cm (Mean = 32.69±7.18) in total length. The average total length of male (33.31±7.54, n = 81) and female (31.99±6.74, n = 71) fish were not significantly different (t = -1.13, p = 0.256 at p<0.05). In the necropsy, stomach, spiral intestine and rectum were isolated, dissected and placed in Petri dishes containing 0.7% saline solution and examined under stereo zoom microscope (LM-52-3621 Elegant). Similarly, gills and skin were cautiously scraped out for the presence of ectoparasites such as monogeneans, copepods and isopods. Specific characters were observed under the Lynx Trinocular microscope (N-800M). Conventional techniques were employed to prepare permanent slides of the collected parasites21,22.
Data analysis: Structure of the component and infracommunities of Rhizoprionodon acutus was explained by the terminologies of Margolis et al.23 and Bush et al.24. Parasite infracommunities were described in terms of prevalence, abundance and intensity however component communities were described in terms of species richness, mean abundance, mean intensity and community similarity such as dominance, diversity and evenness indices. Species were classified as central/core species (if prevalence >66.6%), Secondary species (prevalence between 33.3-66.6%) and satellite species (prevalence <33.3%) of total number of fish analyzed25. Similarly, based on the percentage of prevalence, the nature of infection by parasite species was classified into dominant (>70%), sub-dominant (50-70%), common (30-50%), frequent (10-30%), rare (4-10%) and sporadic (<4%)26. Dispersion index (DI) was used to evaluate the dispersion pattern of parasite species27,28. The distribution of parasites were classified as aggregated (DI>1.96), regular (DI<-1.96) and random (DI<1.96). Parasite diversity of the sample was calculated using Shannon’s (Shannon and Weaver, 1949) index of diversity (H') was used29. For evenness, Shannon-based evenness was calculated. Host size is one of the decisive factors in determining the parasitic communities in any host. Pearson linear correlation coefficient (r) and spearman rank correlation (rs) was computed to determine possible correlation between host standard length with prevalence and abundance of each parasite species, respectively30. The existence of association between species for measuring degree of association is done by Jaccard’s Index (JI) whose value ranges between 0-1 and as the value approaches to 1, indicates the association between species is high.
 | |
| Fig. 1: | Andhra Pradesh map showing the study area, Nellore Coast, Bay of Bengal |
Mann-whitney U-test was used as an indication to scrutinize the influence of host sex on the parasitic abundance (Vassarstat.net/utest.html and http://www.socscistatistics.com). Community structure of parasites has been determined as a function of host habitats, sizes and sexes. The mean and standard deviation type of statistical tests were conducted using MS excel 2010, SPSS IBM 21.0 to evaluate whether significant statistical differences exist in the structure of parasite infracommunities and statistical significance level adopted was p<0.05.
RESULTS
The community structure of Rhizoprionodon acutus studied at both component level and infracommunity level. An infracommunity comprise all parasite species in particular individual host whereas the assemblage of species associated with a particular microenvironment or all infracommunities in a population of hosts make up the component community.
Community ecology of Rhizoprionodon acutus
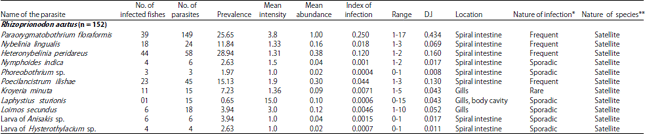
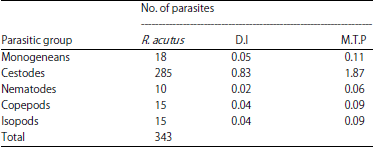
Component community: A total of 11 parasite taxa including 3 ectoparasites (1 monogenean, 1 copepod and 1 isopod) and 8 endoparasites (6 cestodes and 2 larval nematodes) were documented (Table 1). Endohelminthic infection (86%) overwhelmed the ectoparasitic fauna. Cestodes are the most abundant with six species accounting for 83.1% of the total parasites collected followed by monogeneans (5.24%), copepods (4.37%), isopods (4.37%) and larval nematodes (2.91%) which is evidenced by Berger-parker dominance index and mean total parasites (Table 2).
| Table 1: | Basic parameters of parasitic infection in R. acutus (n = 152) from Nellore coast, Bay of Bengal Andhra Pradesh |
 | |
*Common: 30-50%, Frequent: 10-30%, Rare: 4-10%, Sporadic: <4%, **Core sp.: >66%, Secondary sp.: Between 66-33%, Satellite sp.: <33, D.I: Dominance index | |
| Table 2: | Parasitic groups, number of parasites, dominance index (D.I) values and mean total parasites (M.T.P) for parasitic groups from the R. acutus |
 | |
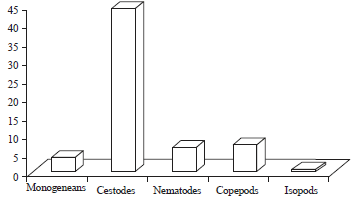 | |
| Fig. 2: | Frequency distribution of number of parasitic groups per individual in R. acutus |
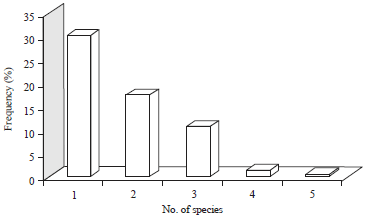
Only 67 (44.07%) hosts were infected with cestodes, 11 (7.23%) hosts with copepods, 10 (6.57%) with nematodes and 1 (0.65%) with isopods (Fig. 2). It was also observed that 67 hosts (44.7%) were single parasitic groups, 18 (11.8%) hosts were infected by 2 parasitic groups and 4 hosts (2.63%) were parasitized by 3 parasitic groups.
Infracommunity: About 90 (59.2%) host fish were parasitized by at least one or more parasitic species. A total of 343 individual parasites were obtained with a mean of 3 parasites/fish. Forty five hosts (29.6%) showed infection with one parasite species and 26 (17.1%), 16 (10.5%), 2 (1.31%) and 1(0.65%) had multiple infections with 2, 3, 4 and 5 parasitic species, respectively (Fig. 3). The cestode, Paraorygmatobothrium floraformis showed high parasitization with 149 parasites (Range = 1-17) collected (43.4% of all parasites) with a mean 1 parasite/fish and the high value of mean relative dominance (0.434) (Table 1). The cestodes, Phoreobothrium sp. (0.87% of all parasites), N. indica and Anisakis larva (1.74% of all parasites), Hysterthylacium larva (1.16% of all parasites), Loimos secundus (5.24% of all parasites) and Lafystius sturionis (4.34%) showed the sporadic nature of infection in the parasitic community.
| Table 3: | Diversity indices of metazoan parasite communities of R. acutus |
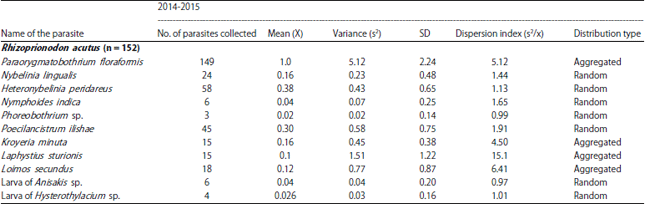
| Table 4: | Mean (X), variance (s2) and dispersion index (s2/x) of parasite species in R. acutus |
 | |
 | |
| Fig. 3: | Frequency distribution of number of parasitic species per individual in R. acutus |
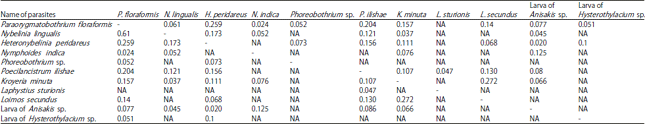
However, P. floramformis, N. lingualis, H. peridareus and P. ilishae showed frequent occurrence in the fishes. Only the copepod, Kroyeria minuta showed rare occurrence in the host (Table 1). None of the hosts showed a core species and secondary species. All the recovered parasites occupied the position of satellite species with less than 33% prevalence (Table 1). The mean parasite diversity (Shannon’s H' index) is 0.39±0.28 and Shannon-based evenness index (E) has a mean of 0.445±0.31 (Table 3). The ratio of variance to mean values gives the index of dispersion. In the present study, P. floraformis, K. minuta, L. sturionis and L. secundus showed greater variance value than the mean which supports the aggregated or over-dispersed nature of parasite infection whereas the other species showed random or under-dispersion or unaggregated distribution (Table 4). Co-occurrence of two species within the same host is termed as an interspecific association. Only parasites sharing common niche such as copepod, K. minuta-monogenean, L. secundus (0.272) within the gills and cestodes, P. floraformis-H. peridareus (0.254), P. floraformis- P. ilishae (0.204), N. lingualis-H. peridareus (0.173) and P. ilishae-H. peridareus (0.156) within the spiral intestine of the host exemplified considerable interspecific associations while the other pairs showed very less to no association (Table 5).
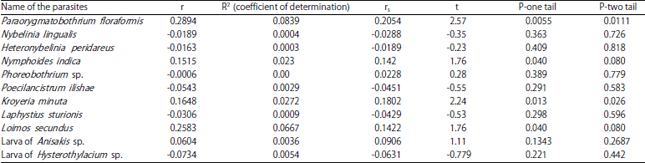
Effect of host length on the metazoan parasites of R. acutus: Pearson’s correlation coefficient ‘r’ was preferred to analyze the probable relationship between host size and parasitization. The positive computed values of r in R. acutus (r = 0.2634, R2 = 0.0694) showed that there is a weak correlation between host size and the parasitization (Table 6). Small and medium sized fish (Group 1 and 2) showed slightly higher infection rate compared to Group 3 and 4 in R. acutus. To ascertain the feasible interrelationship between the host size and individual parasite burden, Pearson’s correlation and Spearman’s rank correlation tests were carried out. Among the eleven individual species in R. acutus, only 5 species i.e., P. floraformis (r = 0.2894, R2 = 0.0839, rs = 0.2054) followed by L. secundus (r = 0.2583, R2 = 0.0667, rs = 0.1422), K. minuta (r = 0.1648, R2 = 0.0272, rs = 0.1802), N. indica (r = 0.1515, R2 = 0.023, rs = 0.142) and Anisakis larva sp. (r = 0.0604, R2 = 0.0036, rs = 0.0906) showed weaker positive correlation with host size while the remaining species showed negative correlation (Table 7).
| Table 5: | Values of Jaccard’s Index (JI) to estimate interspecific association between each pair of parasite species of R. acutus |
 | |
| NA: No association | |
Effect of host sex on metazoan parasites of R. acutus: Of the total sample of 152 R. acutus, 71 are females (46.7%) and 81 are males (53.2%). Among these fish, 45 females (63.3%) and 45 males (55.5%) were parasitized by one or more parasite species. Chi square test was applied for each host species sex-wise individually to scrutinize the effect of host sex on the overall prevalence of infection. Based on a benchmark of 0.05 alpha, the estimated chi-value 0.01365 and p-value 0.711 suggests that there is no statistically significant association occurs between the parasite abundance of males and females of R. acutus. The host sex influence on the parasitic prevalence and abundance of parasites was evaluated through Mann-whitney U-test (Vassarstat.net/utest.html). According to Mann-Whitney Z (U) test, there is a considerable relation between host sex and parasite abundance of each individual parasites in both the hosts (Table 8). Only, L. secundus (Z = 0.784, p = 0.438), H. peridareus (Z = 0.611, p = 0.542) N. lingualis (Z = 0.46, p = 0.645) and larva of Hysterothylacium sp. (Z = 0.52, p = 0.603) in the host, R. acutus showed significant relation between host sex and parasitization while the remaining parasites showed negligible or no relation between sex and the parasitization. Females were slightly more infected than males in R. acutus and the significant values with respect to sex from U-test (z) may be due to the biological, physiological and behavioral variations between the male and female hosts.
DISCUSSION
The interactive-to-isolationist range, i.e., from rich assemblages of species with high interactions, to species-poor assemblages where interactions are unlikely is considered to be one of the illustrious theoretical models in the parasite community ecology research25,31,32. Helminth parasites play a crucial role in modifying elasmobranch population’s demography. Classically, helminth communities of elasmobranchs are considered as species-poor assemblages with unlikely interactions33,34. The parasitic community of R. acutus from Nellore Coast, Bay of Bengal andhra Pradesh also revealed the species-poor assemblages/isolationist range with a total of only 11 parasite taxa including 3 ectoparasites and 8 endoparasites.
| Table 6: | Parasitic abundance in the different size classes of R. acutus |
 | |
| Table 7: | Pearson’s correlation coefficients (r) and Spearman’s rank correlation coefficients (rs) to evaluate the relationship between parasite abundance with host length of R. acutus |
 | |
| Table 8: | Diversity parameters of parasitic species in males and females and values of Mann-Whitney U-test to evaluate rate of host sex and parasitic abundance in R. acutus |
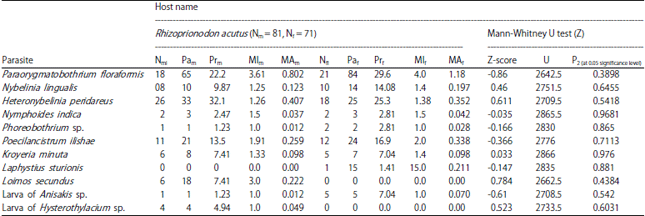 | |
Nmi: Number of males infected, Nfi: Number of females infected, Prm: Prevalence of males, MIm: Mean intensity of males, MAm: Mean abundance of males, Pam: Parasitic of male, MIf: Mean intensity of females, Prf: Prevalence of females, MAf: Mean abundance of females, Paf: Parasitic of female | |
Several factors such as temperature, feeding habits of host, availability of infective intermediate hosts and parasite maturation, complexity of intestinal system, host vagility, host size and phylogeny influence the parasitic communities25,35-40. The analysis of data revealed that the cestode parasites present in the spiral intestine of R. acutus showed a seasonal periodicity and cyclic variability in parasitic burden. Parasite richness increases with increase in the proportion of animal food in the host diet. Diet analysis of R. acutus revealed the presence of intermediate hosts such as molluscs, crustaceans and small fishes which may transmit the larval forms of the parasites to the final host41,42. The study of associated infections traverses with parasite community ecology at the level of infracommunity and much progress in parasite community ecology has been made at this level, because it is the only level at which investigational exploitations of entire infrapopulations are feasible. Interactions between the infrapopulations in individual hosts resolves the coexistence of different parasitic species within single hosts (infracommunity and ultimately in parasite component community). At the level of parasite component community, the concept of core and satellite species concept plays a crucial role in explaining the species interactions25,31,32,43,44. There were no core and secondary species encountered from the host during the present study. Though, H. peridareus and P. floraformis occurred in highest numbers but still they occupied the position of satellite species. According to Bell and Burt45 parasite community diversity is positively correlated with host’s local prevalence. If hosts are sparse, they may lack parasites and he also stated that a certain concentration of hosts and parasites is required to assure the completion of life cycles36. In the present study, the abundance of R. acutus Nellore coast might be the reason for cestode species richness in R. acutus compared to other hosts of the same area13,20. Those parasitic species which are narrowly specific to one species of hosts are called as ‘monoxenic’ or ‘stenadaptive’ parasites whereas those parasites which have wide specificity constitute ‘polyxenic’ or ‘euryadaptive’ species23. All the species of R. acutus were monoxenic species with narrow specificity in the host. Aggregation of parasites is one of the sturdy forces in host population dynamics46. Parasitic helminths display characteristic type of distribution of helminth numbers among the host individuals. They show either over-dispersed pattern of distribution or random or unaggregated pattern of distribution28. The high variance to mean ratio (DI) is an indication of over-dispersed or aggregated pattern of distribution of helminths and only 4 species, L. sturionis, L. secundus, P. floraformis and K. minuta represented aggregated or over-dispersed type of distribution while the rest of the species showed random type of distribution. Jaccard’s coefficient of association (rj) represents the interspecific association among each pair of parasite species. Existence of association among different species might be due to dissimilar habitats47. Parasites sharing the common niche i.e., copepod and monogenean sharing the gill microhabitat and different species of cestodes sharing spiral intestine within the host showed a significant interspecific associations during the present study. Host traits such as length, age, sex, weight and body conditions can extensively control the patterns of infectivity and disease48-52. Length and age of the hosts is one of the prime factors in parasite infra population variations53-56. The present study correlates well with the views of Llewellyn57, Pennycuick58,59 and McVicar60 who suggested low parasitic infection in small sized young fishes, abundant in medium sized adult fishes and much less in large sized old fishes57-59,61. The variation in infection with age group may be because younger small sized and adult medium sized fishes are more susceptible to parasitic infection due to lesser immunological competence while the older fish are highly immunological competent and do not allow extra parasite burdens. At the same time, the parasite life span also plays its role with number of parasites diminishing in the host with increasing age. Host sex is one of the biotic factors which play a vital role in shaping the parasitization in a host. Parasitization in R. acutus agrees with the views of few scientists who supported that females are more susceptible to parasitic infections due to their higher food consumption during spawning periods and immunosuppression due to higher estrogen and progesterone levels60,62-66. Thus, in conclusion, the parasitic communities of R. acutus forms an amorphous assemblage with little resource limitation and competitive influence67-69. There is a severe insufficiency of parasitological information for the majority of shark species, including R. acutus and these types of studies will enable the implementation of new fishery management strategies in shark fishery70 and pave the pathway to use these parasites as biological indicators71.
CONCLUSION
The present findings provide an inclusive data about the structure of the parasite communities in R. acutus at both component community and infracommunity level which can provide a better understanding hosts only 11 parasite taxa including 3 ectoparasites and 8 endoparasites. The communities of metazoan parasites in R. acutus are depauperate and are close to the isolationist extremity of the continuum. The present study would be a great contribution to the knowledge of the metazoan parasite fauna of Elasmobranch fishes to the future helminthologists.
ACKNOWLEDGMENT
This work was financially supported by Council of Scientific and Industrial Research, Government of India and was carried out under the Major Research project (No. 37 (1592)/13/EMR-II). The corresponding author is grateful to CSIR for sanctioning the project. And the first author is thankful to UGC-NFHE for sanctioning the Fellowship.
SIGNIFICANCE STATEMENT
This study unveiled the parasitic community of the Milk shark and their interactions with the host at both the component and infracommunity levels with respect to size and sex of the host. These types of studies will be beneficial for the future researchers to carry out more advanced studies on the shark parasites in different perspectives such as in fishery management, bioindicators of environmental pollution and host biology.
REFERENCES
- Subhapradha, C.K., 1957. Cestode parasites of fishes of Madras coast. Indian J. Helminthol., 7: 41-132.
Direct Link - Srinivasakalyan, C. and V. Anuprasanna, 2016. Taxonomic reports of the cestodes of the genus Nybelinia Poche, 1926 and Heteronybelinia Palm, 1999 (Cestoda: Trypanorhyncha, Diesing, 1863) from the Shark, Rhizoprionodon acutus Rüppell, 1937, from Nellore Coast, Bay of Bengal, India. Int. J. Recent Scient. Res., 7: 9373-9377.
Direct Link - Kalyan, C.S. and V. Anuprasanna, 2016. A morphological study of Kroyeria minuta Pillai, 1968 (Copepoda: Eudactylinidae) infecting the milk shark, Rhizoprionodon acutus Rüppell, 1937 off Nellore Coast, Bay of Bengal, India. J. Applied Life Sci. Int., 9: 1-10.
CrossRefDirect Link - Kalyan, C.S. and A.P. Vankara, 2018. Morphological and molecular identification of Paraorygmatobothrium floraformis in Rhizoprionodon acutus off Nellore Coast, Bay of Bengal, India. Trends Bioinform., 11: 25-32.
CrossRefDirect Link - Poulin, R., 2000. Variation in the intraspecific relationship between fish length and intensity of parasitic infection: Biological and statistical causes. J. Fish Biol., 56: 123-137.
CrossRefDirect Link - Vales, D.G., N.A. Garcia, E.A. Crespo and J.T. Timi, 2011. Parasites of a marine benthic fish in the Southwestern Atlantic: Searching for geographical recurrent patterns of community structure. Parasitol. Res., 108: 261-272.
CrossRefDirect Link - Gudivada, M., A. Vankara, M. Hemalatha and C. Vijayalakshmi, 2014. Ecology of metazoan parasite community of marine threadfin fish, Polydactylus sextarius (Bloch and Schneider, 1801) from Visakhapatnam Coast, Bay of Bengal. Asian J. Anim. Sci., 8: 47-55.
CrossRefDirect Link - Vankara, A.P., S.K. Chadamala and H. Mannela, 2015. Aspects of the ecology of tetraphyllid cestodes from the slender bamboo shark, Chiloscyllium indicum Gmelin, 1789 (Orectolobiformes: Hemiscylliidae) from Nellore Coast, Bay of Bengal, India. South Asian J. Life Sci., 3: 42-50.
CrossRefDirect Link - Espinola-Novelo, J.F., R. Escribano and M.E. Oliva, 2018. Metazoan parasite communities of two deep-sea elasmobranchs: The Southern lanternshark, Etmopterus granulosus and the largenose catshark, Apristurus nasutus, in the Southeastern Pacific Ocean. Parasite, Vol. 25.
CrossRefDirect Link - Rasmussen, T.K. and H.S. Randhawa, 2018. Host diet influences parasite diversity: A case study looking at tapeworm diversity among sharks. Mar. Ecol. Progr. Ser., 605: 1-16.
CrossRefDirect Link - Kennedy, C.R. and H.H. Williams, 1989. Helminth parasite community diversity in a marine fish, Raja batis L. J. Fish Biol., 34: 971-972.
CrossRefDirect Link - Cislo, P.R. and J.N. Caira, 1993. The parasite assemblage in the spiral intestine of the shark Mustelus canis. J. Parasitol., 79: 886-899.
CrossRefDirect Link - Curran, S. and J.N. Caira, 1995. Attachment site specificity and the tapeworm assemblage in the spiral intestine of the blue shark (Prionace glauca). J. Parasitol., 81: 149-157.
CrossRefDirect Link - Morris, T.C., J. van der Ploeg, S.B. Awa, C.D. van der Lingen and C.C. Reed, 2019. Parasite community structure as a predictor of host population structure: An example using Callorhinchus capensis. Int. J. Parasitol.: Parasites Wildlife, 8: 248-255.
CrossRefDirect Link - Kalyan, C.S., M. Hemalatha and A.P. Vankara, 2018. Population dynamics of metazoan parasites of Rhizoprionodon acutus from the Nellore Coast off Bay of Bengal. Annu. Res. Rev. Biol., 28: 1-16.
CrossRefDirect Link - Margolis, L., G.W. Esch, J.C. Holmes, A.M. Kuris and G.A. Shad, 1982. The use of ecological terms in parasitology (Report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol., 68: 131-133.
CrossRefDirect Link - Bush, A.O., K.D. Lafferty, J.M. Lotz and A.W. Shostak, 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol., 83: 575-583.
CrossRefPubMedDirect Link - Poulin, R., 1993. The disparity between observed and uniform distributions: A new look at parasite aggregation. Int. J. Parasitol., 23: 937-944.
CrossRefDirect Link - Shaw, D.J. and A.P. Dobson, 1995. Patterns of macroparasite abundance and aggregation in wildlife populations: A quantitative review. Parasitology, 111: S111-S133.
CrossRefDirect Link - Shannon, C.E. and W. Weaver, 1949. The Mathematical Theory of Communication. 1st Edn., University of Illinois Press, Urbana, IL., ISBN-10: 0252725484, Pages: 117.
Direct Link - Esch, G.W., A.W. Shostak, D.J. Marcogliese and T.M. Goater, 1990. Patterns and Processes in Helminth Parasite Communities: An Overview. In: Parasite Communities: Patterns and Processes, Esch, G.W., A.O. Bush and J.M. Aho (Eds.). Chapman and Hall, London, UK., pp: 1-20.
Direct Link - Sousa, W.P., 1994. Patterns and processes in communities of heirninth parasites. Trends Ecol. Evol., 9: 52-57.
CrossRefDirect Link - Aho, J.M., 1990. Helminth Communities of Amphibians and Reptiles: Comparative Approaches to Understanding Patterns and Processes. In: Parasite Communities: Patterns and Processes, Esch, G.W., A.O. Bush and J.M. Aho (Eds.). Chapter 7, Chapman and Hall, New York, USA., ISBN: 978-94-010-6863-5, pp: 157-195.
- Barton, D.P., 1999. Ecology of helminth communities in tropical Australian amphibians. Int. J. Parasitol., 29: 921-926.
CrossRefDirect Link - Kennedy, C.R., 1971. The effect of temperature upon the establishment and survival of the cestode Caryophyllaeus laticeps in orfe, Leuciscus idus. Parasitology, 63: 59-66.
CrossRefDirect Link - Sen, S., S.K. Chakraborty, E. Vivekanandan, P.U. Zacharia and A.K. Jaiswar et al., 2018. Feeding habits of milk shark, Rhizoprionodon acutus (Ruppell, 1837) in the Gujarat coastal waters of North-Eastern Arabian Sea. Regional Stud. Mar. Sci., 17: 78-86.
CrossRefDirect Link - Caswell, H., 1978. Predator-mediated coexistence: A nonequilibrium model. Am. Naturalist, 112: 127-154.
CrossRefDirect Link - Hanski, I., 1982. Dynamics of regional distribution: The core and satellite species hypothesis. Oikos, 38: 210-221.
CrossRefDirect Link - Bell, G. and A. Burt, 1991. The comparative biology of parasite species diversity: Internal helminths of freshwater fish. J. Anim. Ecol., 60: 1047-1064.
CrossRefDirect Link - Anderson, R.M. and R.M. May, 1978. Regulation and stability of host-parasite population interactions: I. Regulatory processes. J. Anim. Ecol., 47: 219-247.
CrossRefDirect Link - Hubalek, Z., 1982. Coefficients of association and similarity, based on binary (presence-absence) data: An evaluation. Biol. Rev., 57: 669-689.
CrossRefDirect Link - Raffel, T.R., R.P. LeGros, B.C. Love, J.R. Rohr and P.J. Hudson, 2009. Parasite age-intensity relationships in red-spotted newts: Does immune memory influence salamander disease dynamics? Int. J. Parasitol., 39: 231-241.
CrossRefDirect Link - Tinsley, R., L. Stott, J. York, A. Everard and S. Chapple et al., 2012. Acquired immunity protects against helminth infection in a natural host population: Long-term field and laboratory evidence. Int. J. Parasitol., 42: 931-938.
CrossRefDirect Link - Mwita, C.J., 2014. The community of parasites infecting Clarias gariepinus in the Tanzanian waters: A case of Lake Victoria. Open J. Ecol., 4: 873-882.
CrossRefDirect Link - Aydogdu, A., N. Emre and Y. Emre, 2015. Prevalence and intensity of parasitic helminths of thicklip grey mullet Chelon labrosus in hosts in Beymelek Lagoon Lake in Antalya, Turkey, according to season, host size, age and sex of the host. Turk. J. Zool., 39: 643-651.
CrossRefDirect Link - Saha, M., P.K. Bandyopadhyay, A. Roy and S. Ghosh, 2015. Impact of seasons, host age, size and sex on the prevalence of protozoan parasites in ornamental fish. IOSR J. Agric. Vet. Sci., 8: 54-59.
Direct Link - Nyaku, R.E., R.G. Okayi, O.D. Kolndadacha and M. Abdulrahman, 2007. A survey of ectoparasites associated with three species of fish Auchenoglanis ocidentalis, Oreochromis niloticus and Bagrus bayad, in River Benue, Makurdi, Benue State, Nigeria. Proceedings of the 22nd Annual Conference of the Fisheries Society of Nigeria, November 12-16, 2007, Kebbi, Nigeria, pp: 10-14.
- Tasawar, Z., K. Umer and C.S. Hayat, 2007. Observations on lernaeid parasites of Catla catla from a fish hatchery in Muzaffargarh, Pakistan. Pak. Vet. J., 27: 17-19.
Direct Link - Akinsanya, B., A.A. Hassan and A.O. Adeogun, 2008. Gastrointestinal helminth parasites of the fish Synodontis clarias (Siluriformes: Mochokidae) from Lekki lagoon, Lagos, Nigeria. Rev. Biol. Trop., 56: 2021-2026.
PubMedDirect Link - Llewellyn, J., 1962. The life histories and population dynamics of monogenean gill parasites of Trachurus trachurus (L.). J. Mar. Biol. Assoc, U.K., 42: 587-600.
CrossRefDirect Link - Pennycuick, L., 1971. Seasonal variations in the parasite infections in a population of three-spined sticklebacks, Gasterosteus aculeatus L. Parasitology, 63: 373-388.
CrossRefDirect Link - Pennycuick, L., 1971. Differences in the parasite infections in three-spined sticklebacks (Gasterosteus aculeatus L.) of different sex, age and size. Parasitology, 63: 407-418.
CrossRefDirect Link - McVicar, A.H., 1977. Intestinal helminth parasites of the ray Raja naevus in British waters. J. Helminthol., 51: 11-21.
CrossRefDirect Link - Uruku, M.N. and I.A. Adikwu, 2017. Seasonal prevalence of parasites of clarids fishes from the lower Benue river, Nigeria. Niger. J. Fish. Aquacult., 5: 11-19.
Direct Link - Mhaisen, F.T., N.K. Al-Salim and N.R. Khamees, 1988. Occurrence of parasites of the freshwater mugilid fish Liza abu (Heckel) from Basrah, Southern Iraq. J. Fish Biol., 32: 525-532.
CrossRefDirect Link - Balling, T.E. and W. Pfeiffer, 1997. Frequency distributions of fish parasites in the perch Perca fluviatilis L. from Lake Constance. Parasitol. Res., 83: 370-373.
CrossRefDirect Link - Kim, K.H., K.J. Ahn and C.S. Kim, 2001. Seasonal abundances of Prosomicrocotyla gotoi (Monogenea) and Opecoelus sphaericus (Digenea) from greenlings Hexagrammos otakii in a Southern coastal area in Korea. Aquaculture, 192: 147-153.
CrossRefDirect Link - Ozer, A., T. Ozturk and M.O. Ozturk, 2004. Prevalence and intensity of Gyrodactylus arcuatus Bychowsky, 1933 (Monogenea) infestations on the three-spined stickleback, Gasterosteus aculeatus L., 1758. Turk. J. Vet. Anim. Sci., 28: 807-812.
Direct Link - Omeji, S., R.A. Obande and S.T. Member, 2015. Prevalence of endoparasites of Synodontis shcall and Synodontis ocellifer (upside-down cat fish) from Lower River Benue, Nigeria. Int. J. Anim. Biol., 1: 176-181.
Direct Link - Rhode, K., 1991. Intra- and interspecific interactions in low density populations in resource-rich habitats. Oikos, 60: 91-104.
CrossRefDirect Link - Gotelli, N.J. and K. Rohde, 2002. Co-occurrence of ectoparasites of marine fishes: A null model analysis. Ecol. Lett., 5: 86-94.
CrossRefDirect Link - Morris, T., A. Avenant-Oldewage, S. Lamberth and C. Reed, 2016. Shark parasites as bio-indicators of metals in two South African embayments. Mar. Pollut. Bull., 104: 221-228.
CrossRefDirect Link