
Abdullah Adil Ansari
Department of Biological Sciences, Kebbi State University of Science and Technology, Aliero, Kebbi State, Nigeria
Preeta Saywack
Department of Biology, Faculty of Natural Sciences, University of Guyana, Georgetown, Guyana
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 93-99







ABSTRACT
Earthworms are very important organisms, they are both environmentally and economically beneficial and hence their correct identification and classification is very vital. Taxonomy aims to classify organisms based on their similarities and differences. The present study was carried out during the year 2006-2007 at University of Guyana, Georgetown focusing on identification and classification of local earthworm species of Guyana and comparison with a known non-native species (California red). The earthworms were collected (using hand sorting method), cultured and then carefully examined (worms were washed with water, preserved in 10% formalin solution). The two species studied were identified based on their external morphology and internal anatomy as well as their ecological features. The California red earthworm was grouped under the family Lumbricidae and identified as Eisenia foetida, while the local species was grouped under the family Eudrilidae and identified as Eudrilus eugenia.
PDF Abstract XML References Citation
How to cite this article
Abdullah Adil Ansari and Preeta Saywack, 2011. Identification and Classification of Earthworm Species in Guyana. International Journal of Zoological Research, 7: 93-99.
DOI: 10.3923/ijzr.2011.93.99
URL: https://scialert.net/abstract/?doi=ijzr.2011.93.99
DOI: 10.3923/ijzr.2011.93.99
URL: https://scialert.net/abstract/?doi=ijzr.2011.93.99
INTRODUCTION
Earthworm is the common name for the larger members of the Oligochaeta; they belong to the phylum Annelida and are mostly terrestrial organisms found living in the soil. They are described as segmented bristle-bearing worms contributing at large to the biomass of soil invertebrates, particularly in the temperate and tropical regions of the world, (Edwards and Lofty, 1972; Julka, 1993; Ismail, 1997, 2005; Kooch et al., 2008). The principal features of the earthworm is that they are bilaterally symmetrical, externally segmented bearing setae on all segments except the first two, hermaphrodites and lacks a skeleton (Edwards and Bohlen, 1996; Kotpal, 2004). Earthworms can be classified and identified based on many features. These include their morphological, physiological, phylogenetic features as well as their ecological and behavioral characteristics.
Earthworms are scientifically classified under the phylum Annelida, they are over 1800 species of earthworms grouped under this phylum (Sims and Gerard, 1985; Julka, 1988; Martin et al., 2000; Malek, 2007; Kooch et al., 2008; Tripathi and Bhardwaj, 2003). Earthworms belong to the class Oligochaeta. However there is much controversy with the classification of these organisms. Many scientists have developed their own classification schemes and theses have been further revised and developed over the years. Some have placed them in to the class Clitellata making Oligochaeta the subclass. It was Michaelsen 1900 that produced the system that is the basis of the modern taxonomy on earthworms. He divided them into 11 families; he later reorganized his classification into two families (Edwards and Bohlen, 1996). Stephenson later simplified this arrangement into 14 families. There have been four new systems of classification: proposed by Lee (1959) and Jamieson (1971). Some of the main families include: Acanthodrilidae, Ailoscolecidae, Alluroididae, Almidae, Biwadrilidae, Eudrilidae, Exxidae, Glossoscolecidae, Lumbricidae, Lutodrilidae, Megascolecidae, Microchaetidae, Ocnerodrilidae, Octochaetidae, Sparganophilidae.
They have also been grouped into five main families: Lumbricidae, Moniligastridae, Megascolecidae, Eudrilidae, Glossoscolecidae.
Earthworms are distributed all over the world. However they rarely occur in desserts and areas under constant snow and ice, as well as areas lacking soil and vegetation and also on mountain ranges. Some species are found particularly in certain areas (Edwards and Bohlen, 1996).
| • | Lumbricidae | : | Temperate areas of Northern Hemisphere, mostly Eurasia |
| • | Hormogastridae | : | Europe |
| • | Sparganophilidae | : | North America |
| • | Almidae | : | Africa, South America |
| • | Megascolecidae | : | South East Asia, Australia and Oceania, western North America |
| • | Acanthodrilidae | : | Africa, Southeastern North America, Central and South America, Australia and Oceania |
| • | Ocnerodrilidae | : | Central and South America, Africa |
| • | Octochaetidae | : | Central America, India, New Zealand, Australia |
| • | Exxidae | : | Central America |
| • | Glossoscolecidae | : | Central and Northern South America |
| • | Eudrilidae | : | Africa and South Africa |
| • | Moniligastridae | : | Inhabiting India, Sri Lanka, Malaysia and the eastern part of Africa |
The objective of the study was collection of local earthworm species and Californian red earthworm, breeding and culturing of each of the species separately, examination of morphology and anatomy, classification and comparison of both the species.
MATERIALS AND METHODS
Present study was carried out during the year 2006-2007 with the objective of taxonomical overview and description of a local earthworm species and its comparison with non-local species. The majority of the laboratory work was conducted at the University of Guyana.
Earthworms were collected by using handsorting method (Stockli, 1928; Lavelle, 1978; Edwards, 2004). The local earthworm was simply obtained by digging up the soil. Samples were obtained from sites that showed earthworm activity, such as their worm castings. They were also taken from moist soils near ponds and trenches and from garden and farm areas that had manure which are all excellent environments for earthworms, as they are high in nutrients and moisture. Care was taken when digging for the earthworms so as to avoid damaging or killing them. They were then placed in the collecting jars along with some of the same soil they were obtained from. Approximately only 10 to 15 earthworms were required. The California red species was obtained from a local farm. The earthworms were then prepared for breeding and culturing. The baskets were filled with a layer of soil, leaf litter and manure. The two species of earthworms were placed in their respective baskets and left in a dark place. The compost was kept moist by frequent watering and additional leaf litter was added when required. The compost was frequently checked to make sure the earthworms were alive and not escaping from the baskets. A few earthworms were obtained from the compost baskets. They were carefully washed with water, preserved in 10% formalin solution for observation. A digital picture was taken of both species. They were placed in Petri dishes for external examination that included:
| • | Description of the body colour |
| • | Measurement of body length |
| • | The number of segments |
| • | Shape of peristomium |
| • | Ahape and length and position of clitellum |
| • | Presence and position of genital pores |
The examination was repeated with at least two more sample species. An earthworm was obtained and washed to clean. It was placed in a Petri dish with cotton wool soaked in chloroform and covered, until there were no more signs of movement. The earthworm was then removed and placed on the dissecting tray with its ventral side facing up and pinned down. The razor blade was then used to carefully slit the earthworm right down the middle. Too much pressure was not applied so as to avoid damaging any of the internal organs. Pins and a tweezers were then used to carefully spread the earthworm open for examination. Water was sprayed around the sides of the earthworm to wash away any dirt and allow a clear view. It was not poured directly on the specimen as this could wash away some of the fragile organs. The specimen was then carefully examined using a magnifying lens and the dissection microscope. The main features observed were:
| • | No. and position of seminal vesicles |
| • | No. and position of spermathecae |
| • | Crop and gizzard |
| • | The intestine |
The earthworm specimen was washed and a small segment on the ventral side was sliced off using the razor blade. The segment was then washed to remove any dirt and tissues. It was then carefully placed on the slide and a few drops of KOH were added. The slide was then placed over the flame of the of the spirits lamp until the integument turned yellow. It was then left to cool for a few seconds and then a few drops of water were then placed on the slide (Verma, 1989). It was then observed under the microscope, to see if the setae were isolated and visible. The shape and arrangement of the setae were noted and the specimen was drawn. If the setae were clearly visible it was then removed from the microscope and a drop of DPX mount was placed on the specimen to make it a permanent mount. A cover slip was then placed over it carefully to avoid air bubbles. The slide was then stored for later use.
Both the seminal vesicle and spermathecae were removed from a dissected worm using tweezers. They were placed on separate slides and the drop of DPX mount was added and the cover slip placed over it. It was then observed under the microscope and drawn. The slide was stored for later use and reference. The micrometer was set up and the scale was deduced. The slide containing the isolated setae was then placed under the microscope and the setae was aligned to the scale and measured. The length of the body setae was then calculated. Earthworm specimens were kept in glass jars in formalin for later use and reference.
RESULTS AND DISCUSSION
Research has been done over the years to further understand the distribution and taxonomy of earthworms. Much work has been done to document the studies carried out on these organisms. Identification and classification are major topics, which are continually being further developed. The morphological features of the earthworm are of great value in the identification of the species. The size of the earthworm can be used to deduce the species. L. mauritii is 80-210 mm in length and 3.5-5 mm in diameter, while M. posthuma is 115-130 mm in length with a diameter of 5 mm and P. excavatus is 23-120 mm in length with its diameter being 2-5 mm (Ismail, 1997). The number and arrangement of the segments also gives an indication of the species. L. mauritii has about 165-190 segments and is prolobous, while M. posthuma has 140 segemnts and is tanylobous and P. excavatus is epilobous and has 165 segments (Ismail, 1997). The number position of segments forming the clitellum also varies with species. In L. mauritii segments 14 to 17 forms the clitellum and in M. posthuma it is formed by segments 14 to 16 while in P. excavatus it is formed by segments 13 to 17 (Ismail, 1997). Much work has also been done on the geographical distribution of earthworms. This allows us to know which species are found where in the world. Earthworms occur all over the world. Some species are widely distributed and is termed peregrine by Michaelsen, while those that do not spread widely are found in more specific areas are called endemic (Edwards and Bohlen, 1996). Megascolecidae and Lumbricidae are both widely distributed. In the United States they are about 33 lumbricid species (Edwards and Bohlen, 1996). Lee 1985 reviewed the distribution of peregrine species; about 3% of all earthworms are peregrine. 20-30 species of Lumbricidae have been spread from Northern and Western Europe by man and now are dominant temperate and tropical regions of the world (Edwards and Bohlen, 1996). Fifteen to twenty species of several Megascolecid families originated from eastern and southeastern Asia and now are established throughout most of the tropical world (Yousefi et al., 2009). Microscolex dubius is thought to be of South American origin and is now established throughout northern temperate zone and North America (Edwards and Bohlen, 1996). The glossoscolecid Pontoscolex corethrurus was originally from South America and now is found in tropical and warmer temperate regions (Edwards and Bohlen, 1996). Both physical factors and nature of organic matter of soils have been known to influence the abundance and distribution of earthworms. The major physical factors are soil water content and mineral matter. The organic matter influences the abundance and species diversity of earthworms (Julka, 1988, 1993; Kale, 1998; Kooch et al., 2008).
Earthworms are segmented bristle bearing worms (Ismail, 1997, 2005). They belong to the phylum Annelida and are the larger members of the Oligochaeta. Taxonomically they are five large families of earthworms. These include; Moniligastridae, Megascolecidae, Eudrilidae, Glossoscolecidae and Lumbricidae (Julka, 1988; Ismail, 1997, 2005; Yousefi, et al., 2009). Morphologically earthworms have little variation. They differ mainly in length, segmentation, clitellum and external apertures.
The two species studied were identified based on their morphology and anatomy as well as their ecological features. The California red earthworm was grouped under the family Lumbricidae and identified as Eisenia foetida, while the local species was grouped under the family Eudrilidae and identified as Eudrilus eugenia.
Eisenia foetida is more commonly known as manure worm, tiger worm and the red wiggler. Its specific name arises because when handled roughly it exudes a pungent liquid. Its colour was a key identification feature. Its colour can range from purple to red to a dark or brownish red. However, it has an unusual pigmentation. The pigment is not evenly distributed, but appeared as dark segmental bands separated by lighter inter segmental bands (Edwards and Bohlen, 1996).
| Table 1: | Showing the taxonomic characters of both the local and California red earthworm species |
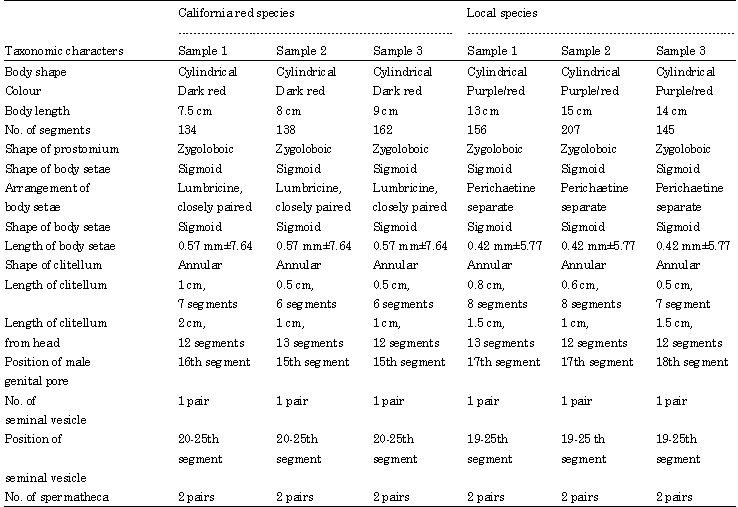 | |
Eudrilus eugenia is a purplish red worm, cultured as the African night crawler. Based on habitat and life cycle both these species were classified as epigeic organisms. Epigeic organisms live in organic horizons, they were found in the natural environment in the upper topsoil layer about one foot (Ismail, 1997, 2005). Here they fed on decaying organic matter and exhibited fast movement. However the Eisenia foetida exhibited more aggressive movement. The Eisenia foetida is found mostly in moist manure and thrives in environments such as composts heaps, forests, gardens and cowpats. They as well as the local species are detritivores as they fed at or near the soil surface on mostly plant litter.
The local species (Table 2) belongs to the family Eudrilidae that is restricted to Africa; however, the Eudrilus eugenia species has become widely distributed in the warmer parts of the world (Table 2).
Body length was another diagnostic feature. Ideally Eisenia foetida ranges from 35-130 mm, from measurement of the sample worms they too fell into this range, while the local species fell into the range of 111-300 mm, which is characteristic of Eudrilidae and night crawlers (Table 1).
Shape and arrangement of body setae was another determining characteristic. Both species had a sigmoid shape (Table 1). This is one of the most common shapes in earthworms. However both their setae arrangement varied. Eisenia foetida had a lumbricine closely paired arrangement that is characteristic of the Lumbricidae family. The lumbricine arrangement consists of eight setae per segment in ventral and latero ventral pairs (Edwards and Bohlen, 1996). The local species displayed a perichaetine arrangement, where the setae was arranged in a ring right around the segment, with a large or small break in the mid dorsal and mid ventral regions (Edwards and Bohlen, 1996).
| Table 2: | Identification of earthworm species based on the above characters |
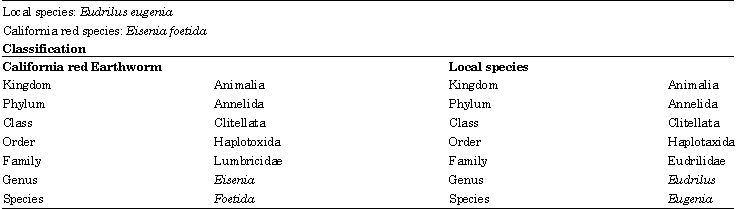 | |
This arrangement is seen particularly in the Megascolecoidea super family (Edwards and Bohlen, 1996) to which the local species belongs (Table 1).
Genital pores were also a significant taxonomic character. The male pores of eurilids lie in segment 17 and this was seen in the local species, in lumbricids the male pores are usually found around segment 15 (Edwards and Bohlen, 1996). The California red species had its male pore around segment 15 and 16, indicating its relation to lumbricidae family (Table 1).
Other key diagnostic features were the clitellum, which is the glandular thickening of the epidermis. In both species (Table 1) they were found to the anterior of the body and ranged from about 4 to 10 segments long. This is characteristic of the lumbricidae family as well as its annular shape (Edwards and Bohlen, 1996). Both species (Table 1) had one pair of seminal vesicles and two pairs of spermathecae. The position of the spermathecae in Eudrilids is a characteristic feature. Unlike most earthworms their spermathecae migrates from the usual anterior position to the vicinity of the ovaries around segment 13. This was observed in the local species.
Both these species (Table 2) were similar in most respects and their vital systems functioned in the same way. However, a major differentiating feature is that Eudrilus eugenia has an unusual mechanism for internal fertilization in a special chamber, unlike most earthworms that exhibit external fertilization (Edwards and Bohlen, 1996).
Eisenia foetida and eudrilus Eugenia (Table 2) are also both ideal worms for vermiculure as they are easy to grow in high-density cultures. Esienia foetida is the most useful in this area as it can tolerate the widest range of environmental conditions and disruption to its environment, unlike the local species, which cannot tolerate this. The local species is however ideal for use as bait worm because of its large size (Ismail, 1997, 2005).
On conclusion, earthworms are very important organisms; their roles not only lie in the environment but also have expanded towards agriculture and even medicine. Earthworms occur all over the world and they are over 1800 species, each having ideal requirements and functions hence, it is important to accurately identify and classify these organisms. The local species identified in this research project was Eudrilus eugenia of the family Eudrilidae which is an ideal earthworm for fish bait and use in vermiculture.
REFERENCES
- Kooch, Y., H. Jalilvand, M.A. Bahmanyar and M.R. Pormajidian, 2008. Abundance, biomass and vertical distribution of earthworms in ecosystem units of hornbeam forest. J. Boil. Sci., 8: 1033-1038.
CrossRefDirect Link - Lee, K.E., 1959. A key for the identification of New Zealand earthworms. Tuatara, 8: 13-60.
Direct Link - Yousefi, Z., M. Ramezani, S.K.A. Mohamadi, R.A. Mohammadpour and A. Nemati, 2009. Identification of earthworm species in Sari Township in Northern Iran, 2007-2008. J. Applied Sci., 9: 3746-3751.
Direct Link
seifollah farmohammadi Reply
Dear Abdullah , I think you have good experience in Identification of earth worm ,do you like to have a mutual cooperation ? It's a very useful paper may I have a copy of it ?