
Anu Prasanna Vankara
Department of Animal Sciences, Yogi Vemana University, Kadapa-516 003, India
G. Mani
Department of Zoology, Maharajah�s College (Autonomous), Vizianagaram, India
C. Vijayalakshmi
Department of Zoology, Andhra University, Visakhapatnam-530 003, India
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 19-33







ABSTRACT
Mastacembelus armatus is considered to be the delicacy of Southern India. Four hundred ninety four specimens of M. armatus collected from river Godavari from August 2005 to September 2007 were analyzed in order to study their metazoan parasite infracommunities. Twelve species of parasites were collected, 6 digenea, 2 cestodes, 1 monogenea, 1 copepoda, 1 nematoda and 1 acanthocephala; 78% of the fishes were parasitized by one or more than one metazoan, with a mean of 67 parasites/fish. The endoparasites represents 98.3% of the total parasites collected. The digenean Tetracotyle sp. and Circumonchobothrium shindei occupy the position of secondary species and the remaining were satellite species. Relationships between total body length of fish and both total parasite abundance and mean parasite species richness were observed. A new copepod species, Neoergasilus indicus is also encountered in the present investigation. The metazoan parasite infracommunities of M. armatus presented dominance of larval endoparasites; correlation of parasite burden, diversity and species richness with host total length; and no influence of host sex on parasitisation.
PDF Abstract XML References Citation
Received: May 12, 2010;
Accepted: July 25, 2010;
Published: August 21, 2010
How to cite this article
Anu Prasanna Vankara, G. Mani and C. Vijayalakshmi, 2011. Metazoan Parasite Infracommunities of the Freshwater Eel, Mastacembelus armatus Lacèpéde, 1800 from River Godavari, India. International Journal of Zoological Research, 7: 19-33.
DOI: 10.3923/ijzr.2011.19.33
URL: https://scialert.net/abstract/?doi=ijzr.2011.19.33
DOI: 10.3923/ijzr.2011.19.33
URL: https://scialert.net/abstract/?doi=ijzr.2011.19.33
INTRODUCTION
Parasites are small players with crucial role in ecological theatres is well said by Marcogliese (2004) as he signified parasites as imperative tools for the effectual management of our natural resources by having an ecosystem based approach. Parasites can provide information on population structure, evolutionary hypotheses, environmental stressors, trophic interactions, biodiversity and climatic conditions. Parasites are metabolically dependent on their hosts mainly for their nutritional requirements (Marcogliese, 2004). However, it is an established fact that many species of parasites provoke pronounced or subtle effects on hosts affecting their behaviour, growth, fecundity and mortality (Marcogliese, 2004). Moreover, parasites may also regulate host population dynamics and influence community structure (Malan et al., 1997; Marcogliese, 2004; Hatcher et al., 2006; Vignon and Sasal, 2010). It is observed that the parasites act as indicators of changes in the ecosystem structure and function environment and thus, eventually reflecting the position of host in the food web. The study of ecological concepts of parasites provides an overall view of interrelationships between parasites, the host and the environment (Marcogliese, 2004). Parasitism is very important biotic factor as in this association the host is under the influence of the parasite. The structure of metazoan parasite community of fish is receiving major attention of ecologists. Parasitic communities are a complex of individuals belonging to the different parasite species inhabiting a host (Cloutman, 1975). The study on seasonal dynamics of parasitism levels serves as a tool to understand broad aspects that determine the population biology of the host-parasite system (Chubb, 1982). Parasite communities generally lie somewhere along the interactive-to-isolationist continuum, i.e., from rich assemblages of species with high colonisation rates in which interspecific interactions play an important structuring role, to species-poor assemblages where interactions are unlikely. This framework has become one of the paradigms of parasite community ecology. Holmes and Price (1986) gave a theoretical consideration which shows that helminth communities span a continuum between interactive to isolationist. Vast majority of work has been focused on the metazoan parasitic communities in fishes including both ectoparasites and endoparasites (Thoney, 1991; Aho and Bush, 1993; Campos and Carbonell, 1994; Molloy et al., 1995; Luque et al., 1996a, b; Alves and Luque, 2001a, b, Aloo, 2002; Alves et al., 2002; Guidelli et al., 2003; Aguilar-Aguilar and Salgado-Maldonado, 2006; Munoz et al., 2007; Hernandez et al., 2007). The present study aims to develop an analysis of community structure in the Mastacembelid fish, Mastacembelus armatus. Mastacembelus armatus Lacépède, 1800 is the largest spiny eel of the genus Mastacembelus. It is a popular indigenous aquarium fish and also an economically important food fish (Sugunan et al., 2002; Tripathi, 2004). It is esteemed as highly proteinaceous food and attributed as tasty and medicinally important fish. They are commonly known as zigzag eel, spiny eel, leopard spiny eel and white-spotted spiny eel. It is also locally called as Pedda papera or papera or freshwater Baam or Bommidai and harbours a variety of metazoan parasitic fauna which includes monogeneans, digeneans, cestodes, nematodes, acanthocephalans, copepods and isopods. Taxonomic studies on metazoan parasites of M. armatus from various parts of the world includes those of Harshey (1937), Karve and Naik (1951), Gupta (1958), Jain (1958, 1959), Tripathi (1959), Kulkarni (1969), Verma (1973), Shinde and Jadhav (1980), Venkatanarsaiah (1981), Agarwal and kumar (1981), Agrawal and Singh (1982), Agrawal and Agarwal (1988), Agrawal and Sharma (1989), Desmukh and Shinde (1989), Hasnain (1992), Jadhav et al. (1990), Dubey et al. (1992), De and Dey (1992), Noopur et al. (1994), Wongsawad and Jadhav (1998), Kritsky et al. (2004), Jalali and Barzegar (2006) and Jalali et al. (2008). However, there are no studies on the populational and ecological features of metazoan parasites in these fishes. Hence, in this report, we analyse at the component and infracommunity levels a metazoan parasite community of M. armatus from the river Godavari, India.
MATERIALS AND METHODS
494 Mastacembelus armatus were procured from the river Godavari, Rajahmundry and local fish markets in and around the river and were brought to the laboratory for the examination of parasites from August 2005 to September 2007. Fishes were identified according to Jayaram (1981), Munro (1982) and Day (1994). Length, weight and sex of each fish individually were noted carefully and all organs were examined separately for the collection of parasites. Parasites were collected, enumerated and permanent slides were prepared by preserving the parasites in AFA (Alcohol, Formaldehyde and Acetic acid in 85:10:5) and stained with Alum carmine (Hiware et al., 2003; Madhavi et al., 2007). M. armatus measured 18-52 cm (mean = 36.42±6.69 cm) in total length. The average total length of male (35.4±6.59 cm, n = 221) and average length of female (37.32±6.64 cm, n = 273) fishes in the sample were not significantly different (t = -1.0, p = 1.00). The dominance frequencies of each parasite infracommunity were calculated by Berger-Parker index of dominance (Magurran, 1988). Pearson’s correlation coefficient r was used as an indication of the relationship between the host’s length and prevalence of parasites. The effect of host sex on abundance and prevalence of parasites was tested by applying chi-square test. Parasite species diversity was calculated using Shannon-Weiner index (H'). For each infracommunity, the evenness (Shannon-based evenness index, E) was calculated. Diversity H' and evenness (E) were calculated only for those fish with more than one parasitic species (diversity not defined for K=1).
RESULTS
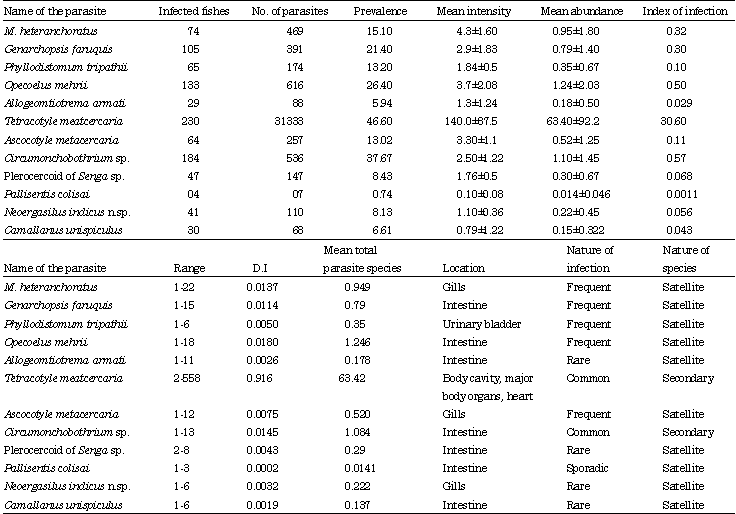
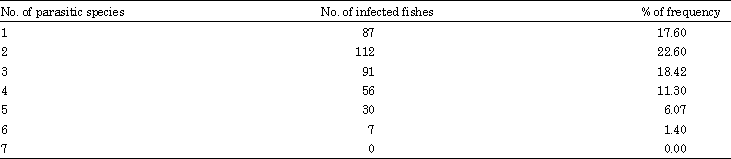
Community ecology of Mastacembelus armatus: Twelve species of metazoan parasites (6 species of digenea, 2 cestodes, 1 monogenea, 1 copepoda, 1 nematoda and 1 acanthocephala) were collected from the host during the study period (Table 1). In M. armatus, the parasitic fauna is predominated by digeneans followed by cestodes, monogeneans, copepods and the remaining groups at negligible level. This is evidenced by the Berger-parker’s dominance indices and mean total parasites (Table 2). 187 hosts (38%) of M. armatus showed infection with single parasitic group, 158 (32%) with two, 36 (7%) with three and a negligible number i.e., 2 (less than 1%) with four parasitic groups respectively (Table 3). No single fish has found infected with all the parasitic groups simultaneously. Of the six species of Digenea, two are larval stages, Tetracotyle sp. and Ascocotyle sp. and other four are adults, Genarchopsis faruquis, Phyllodistomum tripathii,Allogomtiotrema armati and Opecoelus mehrii.
| Table 1: | Diversity parameters and distribution patterns of parasitic species of Mastacembelus armatus |
 | |
| Table 2: | Parasitic groups, number of parasites obtained, dominance index and mean total parasites in M. armatus |
 | |
| Table 3: | Frequency distribution of number of parasitic species per individual in Mastacembelus armatus |
 | |
| Table 4: | Frequency distribution of number of parasitic species per individual in M. armatus |
 | |
| Table 5: | Diversity parameters of metazoan parasite communities of M. armatus |
 | |
The parasite fauna of the host is exclusively predominated by the larval stages of the strigeid trematode, Tetracotyle sp. with 31333 parasites and constitutes about 96% of the total parasites collected. Two species of cestodes, Circumonchobothrium shindei together with its plerocercoid stages and plerocercoid stage of Senga sp. dominates the parasite fauna next to Tetracotyle sp. Of the 494 fishes examined, 383 (78%) fish were frequently parasitized with one or more than one parasite species. A total of 34,196 (Range = 2-558) individual parasites was collected with a mean of 67 parasites/fish. Eighty seven (18%) hosts showed infection with single parasite species, 112 (23%), 9 (18%), 56 (11%), 30 (6%) and only 7 (1%) hosts showed multiple infections with 2, 3, 4, 5 and 6 parasite species respectively (Table 4). Mean parasite species richness is 2.20±0.42. The mean parasite diversity (Shannon’s H' index) is 0.714±0.47 and the maximum diversity is 1.75. Shannon-based evenness index (E) has a mean of 0.927±0.597 (Table 5). Tetracotyle larvae showed the highest value of index of dispersion, followed by metacercariae of Ascocotyle nana and Mastacembelocleidus heteranchoratus for both the cycles (Table 6).
| Table 6: | Comparison of the mean (X), Variance (s2) and Dispersion index (s2/X) of parasite species in M. armatus |
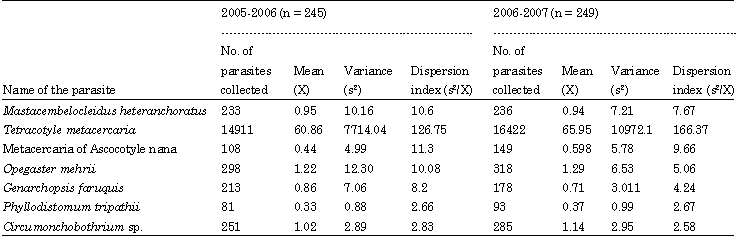 | |
The calculated values of chi-square for both the years are much lower than the table values suggesting the fitting of Poisson distribution pattern for the parasitic species. Grabda-Kazubski et al. (1987) classified the nature of infection by parasite species into dominant (>70%), sub-dominant (50-70%), common (30-50%), frequent (10-30%), rare (4-10%) and sporadic (<4%) based on the percentage of prevalence (Table 1). In the present study, Tetracotyle sp. and Circumonchobothrium shindei show common infection with a prevalence ranging between 30-50%. Plerocercoid of Senga, Neoergasilus indicus and Camallanus unispiculus show rare infections (4-10%). Pallisentis colisai shows sporadic infection (<4%) whereas M. heteranchoratus and metacercaria Ascocotyle nana shows frequent infection. The abundance of parasitic species, range, prevalence, mean intensity, mean abundance and index of infection were calculated (Table 1). In the present investigation, based on the studies of Bush et al. (1997) it is observed that there is no core species in the host. Only secondary and satellite species were encountered. Two species, metacercaria Tetracotyle sp. and C. shindei occur in large numbers and occupy the position of Secondary species while the rest of the parasite species, monogenean-M. heteranchoratus, digeneans-metacercaria A. nana, Genarchopsis faruquis, Phyllodistomum tripathii, Opecoelus mehrii, Allogomtiotrema armati, copepod-N. indicus, nematode-C. unispiculus., cestode- Plerocercoid of Senga sp. and Acanthocephalan-P. colisai are satellite species (Table 1). Mean intensity and mean abundance are exceptionally very high for Tetracotyle sp., moderate for rest for the species and negligible in P. colisai. Berger-Parker’s dominance index and mean of total parasite species was calculated for each species of parasite in the hosts (Table 1). Tetracotyle sp. presented the highest dominance value of 0.916 followed by the digenean, O. mehrii (0.018) and cestode, C. shindei (0.0145). Mean of total parasitic species of Tetracotyle sp. is 63.42, 1.246 for O. mehrii and 1.084 for C. shindei. The pairwise associations between the larval helminths were calculated exclusively since the parasitic fauna of the fish is predominated by larval helminths. The pairwise assosciations were calculated for A. nana and Tetracotyle sp. in M. armatus by using Jaccard’s coefficient (rj) and modified Sørenson’s coefficient. Jaccard’s coefficient of association was low for one pairwise combination in M. armatus suggests that the two species in a pair do not co-occur very often in the same fish individual (Table 7). However, Sørensen’s coefficient for the pairwise combinations in the fishes showed negative coefficient values suggesting that the two species in a pair do not co-occur recurrently in the same fish (Table 8). Host size also serves as a factor in determining the burden of parasitic communities in a host. In the present study, M. armatus from 18 cm to 53 cm were collected and were categorized into 9 classes. The possible interrelationship between the host size and parasite burden were carried out by Pearson’s correlation tests. Pearson’s coefficient was 0.411 for M. armatus which suggests that the total body size and total parasite burden is positively correlated.
| Table 7: | Matrix of pairwise assosciations between larval helminths using Jaccard’s coefficient, rj; each fish sample treated separately |
 | |
| Table 8: | Matrix of pairwise assosciations between larval helminths using modified Sorensen’s coefficient, rs; each fish sample treated separately |
 | |
| Table9: | Correlation of parasite species richness, parasite species diversity and evenness in different size classes of M. armatus |
 | |
Overall parasite richness and parasite diversity are poorly correlated with the host size. However, ‘r’ values have also been calculated for individual species in relation to host body length (Table 9). Of the seven individual species most prevalent species (>10%) the cestode, C. shindei (r = 0.461), copepod, N. indicus (r = 0.514) and digeneans, A. nana (r = 0.706), Tetracotyle sp. (r = 0.402) and O. mehrii (r = 0.334) and the monogenean, M. heteranchoratus (r = 0.638) presented significant correlations between host total length and parasite abundance. Host sex is one of the biotic factors which play an important role in determining the parasitization in a host. Of the total sample of 494 M. armatus, 273 were females (55%) and 221 were males (45%). Among these fish, 225 females (46%) and 157 males (32%) were parasitized by one or more speices of parasites. Based on a benchmark of 0.05 alpha, the estimated p-value of 0.00 suggests that there is no statistically significant association between the parasite abundance of males and females and that the host sex has no influence on parasitization. But the impact of host sex on individual parasite species were studied of which only the metacercaria of Tetracotyle sp. and digenean, G. faruquis, showed relatively higher values of prevalence and intensity of infection in females than in males (Table 10). To find the significant differences in parasite diversity of males and females, t-test was conducted. The estimated values of t = -0.405, p<0.686 suggests that there is no statistically significant difference in the parasite diversity between males (H' = 0.73±0.47) and females H' = (0.702±0.474).
| Table 10: | Diversity parameters of parasitic species in males and females of M. armatus |
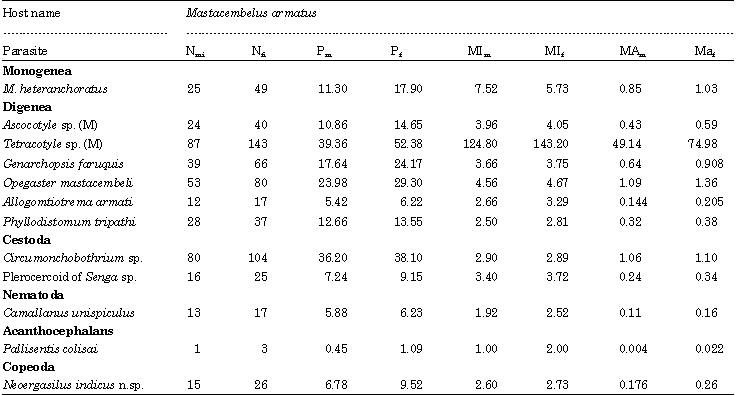 | |
| Nm : No. of males examined; Nf : No. of females examined; Nmi : No. of males infected; Nfi : No. of females infected; Pm and Pf : Prevalence of males and females respectively; MIm and MIf : Mean intensity of males and females; MAm and MAf : Mean abundance of males and females respectively | |
DISCUSSION
Some patterns in the structure and composition of the community of metazoan parasites of M. armatus were detected: (1) dominance of endoparasitic larval stage; (2) positive correlation of parasitic burden and richness and (3) No influence of host sex on parasitization. Majority of freshwater fishes act as second intermediate hosts for many adult avian trematodes (Sukontason et al., 1999; Wongsawad et al., 2004; Kue-A-Pai and Wiwanitkit, 2005) and hence the metacercarial infection is found to be heavy in these fishes. The density of larval parasitic fauna is found to be higher than diversity of adult parasitic spectrum. M. armatus is also considered as good intermediate or paratenic hosts since the parasitic fauna of these fishes is predominated by larval helminths. Larval helminths use fish as intermediate hosts for the completion of their life-cycles in the definitive vertebrate hosts (Luque and Poulin, 2004). Many scientists focussed their studies on the adult helminths in the gastrointestinal tract of fish; few on ectoparasites since these endohelminths and ectoparasites form two distinct guilds of parasites inhabiting the same host and compete for the same resources. But larval helminths does not form an interactive guild as they appear less likely to compete for resources since they seek shelter in various organs and because many of them are in the form of cysts and do not actively feed on host nutrients or tissues. The statistical association of larval helminths among fish hosts have received little attention. In the present study community ecology of larval helminths along with their associations and adults is considered. However, M. armatus depicts a different scenario with high parasitization throughout the year which can be attributed to the non-random assemblages of larval parasites, which is an adaptive strategy of larval helminths to increase their probability of reaching their next host successively. Diet, feeding habits, vagility of host species are the main factors affecting the parasitic community structure especially for the parasites transmitted to their final host (Sasal et al., 1997). Also the classical studies on the impact of diet and feeding habits on the parasitic fauna (community structure) of the hosts were carried out by Pearson (1968), Kuperman (1973), Cannon (1977), Williams and Jones (1994), Luque et al. (1996a) and Johnson et al. (2004). In the present study impact of food and feeding habits of the host can be considered as one of the prime cause of larval parasitic abundance as M. armatus are carnivorous and their diet constitutes insects, crustaceans, molluscs, snails and worms which act as primary intermediate hosts for the most of the digeneans and cestodes. Many parasitic communities are known to occupy precise, predictable and limited locations (sites) within their hosts (Crompton, 1973; Holmes, 1973; Crompton and Nesheim, 1976). Holmes and Price (1986) suggested that the number of species of parasites that a host species supports widely varies from host to host which might be due to the biotic and abiotic factors. Various factors responsible for the parasite composition within a host is host size, phylogeny, composition of diet, complexity of gut and vagility. It is true in case of M. armatus as these fishes occur in shallow waters where the movement is limited. Kennedy and Bush (1994) are of the opinion that the examination of helminth community structure at the level of host’s family and genus over a particular geographical area will improve our understanding of unpredictable helminth communities and their determinant factors. This statement is in total agreement with the present study. It is interesting to note that the 12 species of both ecto and endoparasites reported in the present study. Some of them have been recorded previously from other localities from these hosts. However, a few parasitic species are reported for the first time from this locality. Only copepod, Neoergasilus indicus is the new species report encountered from M. armatus in the present study (Vankara and Chikkam, 2010).
Kennedy et al. (1986) hypothesized freshwater fish parasitic communities to be less diverse when compared to their counterparts’ marine fishes due to poor diversity of intermediate hosts in freshwater environment. The richly nutritive habitat and the healthy nature of fish might have declined the diversity and density of infection when compared to the marine fishes. In the present study, M. armatus also exhibited poor parasitic diversity and holds good to the hypothesis of Kennedy et al. (1986). Pairwise interspecific associations among larval helminth species in M. armatus were carried out using two different coefficients of association (Jaccard’s and Sorensen’s coefficients of association). Values of Jr and Js coefficients of association for Tetracotyle-Ascocotyle pairwise associations in M. armatus suggests that two species in a pair do not co-occur very often in the same fish individuals. The present study is in accordance with the views of Poulin and Valtonen (2001) which suggests that assemblages of larval helminth parasites in fish are not random selections of locally available species, but rather non-random packets of larval parasites that travel together along common transmission routes. Few parasitic species are narrowly specific to one species of hosts which are called as ‘monoxenic’ or ‘stenoadaptive’ parasites whereas the parasites with wide specificity constitute ‘polyxenic’ or ‘euryadaptive’ species (Margolis et al., 1982). In the present study, none of the parasitic species of M. armatus are monoxenic. At the level of parasite component community, the concept of core and satellite species concept plays a crucial role in explaining the species interactions (Holmes and Price, 1986; Esch et al., 1990; Sousa, 1994). In the present study, metacercaria Tetracotyle sp. and adult Circumonchobothrium shindei occupy the position of secondary species but these parasites inhabit different niches. C. shindei in the intestine and Tetracotyle sp. inhabiting each and every organ, heart, liver, coelom cavity etc. Helminth communites may be characterized as being isolationists or interactive and they are recognized only when the species regularly co-occur (Holmes and Price, 1986). Tetracotyle sp. and C. shindei share with other species of trematodes and cestodes, so can be called as interactive species. Shotter (1973), Bell and Burt (1991), Machado et al. (1994), Poulin (1995), Fiorillo and Font (1996) and Luque et al. (1996a, b) stated that helminth diversity and parasite community richness are more consistently correlated with host size. Cotgreave (1993), Blackburn and Lawton (1994) and Blackburn and Gatson (1997) documented relationship between parasite abundance and body size. Takemoto and Pavanelli (1994) and Machado et al. (1994) found that increase in size and age of fish resulted a significant increase in the levels of parasitism. Llewellyn (1962), Pennycuick (1971a, b), Shotter (1973), McVicar (1977), Hanek and Fernando (1978a, b), Fernandez (1985), Valtonen et al. (1990) and Roubal (1990) found higher levels of parasitism in hosts with intermediate lengths. Fish acquire the parasites in their youth phase which are then eliminated in the fish’s adult phase which might be due to ageing of parasites or immunological resistance of fish. The parasitization in M. armatus is in harmony with the above studies as the prevalence of infection is low in small fish, abundant in medium size and then decreased in larger fishes. Host sex is one of the biotic factors which are important in studying host-parasite relationships. Effects of host sex on the levels of parasitism was studied by Thomas (1964) which might be due to the physiological and biological factors and behavioural difference between males and females which produce a small albeit consistent sexual trend in infection levels. Females are more likely to be heavily infected when compared to males which might be due to the stress caused during reproductive periods and lead into behavioural changes, thus making them more vulnerable to heavy parasitzation. In contrast, Folstad and Karter (1992) and Poulin (1996) found more parasitization in males which might be due to high levels of testosterone which causes immunosuppresion making them more susceptible to parasites than females. But there are also studies by Lawrence (1970), Pennycuick (1971a, b), Kennedy (1975), Muzzall (1980), Belghyti et al. (1994), Takemoto and Pavanelli (1994, 2000), Machado et al. (1994), Luque et al. (1996a) and Lizama et al. (2005) who found host sex not to be a significant factor in determining the infection rate of helminth parasites in host fishes and the present study is in harmony with the above studies. However, individual parasites, C. shindei and G. faruquis presented a significant impact in relation to sex as females showed relatively higher parasitization for these parasites. The present ecological studies suggest that the parasite communities of M. armatus are predictable, depauperate and non-interactive. Parasite diversity and richness can be attributed to its phylogeny, presence of intermediate hosts in the area. Abundance of parasite population is attributed to feeding capacity of the host which is governed by biotic and abiotic factors like temperature, water currents etc. The present study also holds good with the views of Holmes (1990) which suggests that fish as intermediate hosts have rich parasite fauna since they harbour both adults and larval helminths. The freshwater fish parasitic communities are less diverse than their marine counterparts (Holmes, 1990).
ACKNOWLEDGMENTS
One of the authors, APV is grateful to Council of Scientific and Industrial research, New Delhi for providing financial assistance to carry out the research.
REFERENCES
- Alves, D.R. and J.L. Luque, 2001. Community ecology of the metazoan parasites of the white-croaker Micropogonias furnieri (Osteichthyes: Sciaenidae) from the coastal zone of State of Rio de Janeiro, Brazil. Mem Inst Oswaldo Cruz., 96: 145-153.
Direct Link - Alves, D.R. and J.L. Luque, 2001. Quantitative aspects of metazoan parasites infrapopulations of Micropogonias furnieri (Osteichthyes: Sciaenidae) from the coastal zone of Rio de Janeiro, Brazil. Parasitological Dia., 25: 30-35.
CrossRefDirect Link - Alves, D.R., J.L. Luque and A.R. Paraguassu, 2002. Community ecology of the metazoan parasites of Pink Cusk-eel, Genypterus brasiliensis (Osteichthyes: Ophidiidae), from the coastal zone of the State of Rio de Janeiro, Brazil. Memorias Instituto Oswaldo Cruz., 97: 683-689.
PubMedDirect Link - Vankara, A.P. and V. Chikkam, 2010. A new ergasilid copepod parasitic on the freshwater spiny eel, Mastacembelus armatus Lacepede, 1800 from river Godavari, India. Crustaceana, 83: 425-434.
CrossRefDirect Link - Belghyti, D., O. Berrada-Rkhami, V. Boy, P. Aguesse and C. Gabrian, 1994. Population biology of two helminth parasites of flatfishes from the Atlantic coast of Morocco. J. Fish Biol., 44: 1005-1021.
CrossRefDirect Link - Bell, G. and A. Burt, 1991. The comparative biology of parasite species diversity: Internal helminths of freshwater fish. J. Anim. Ecol., 60: 1047-1064.
CrossRefDirect Link - Blackburn, T.M. and K.J. Gatson, 1997. A critical assessment of the form of the interspecific relationship between abundance and body size in animals. J. Anim. Ecol., 66: 233-249.
Direct Link - Blackburn, T.M. and J.H. Lawton, 1994. Population abundance and body size in animal assemblages. Philosophical Trans. Biol. Sci., 343: 33-39.
Direct Link - Bush, A.O., K.D. Lafferty, J.M. Lotz and W. Shostak, 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol., 83: 575-583.
PubMed - Chubb, J.C., 1982. Seasonal occurrence of helminths in freshwater fishes: Part IV. Adult Cestoda, Nematoda and Acanthocephala. Adv. Parsitol., 20: 1-292.
PubMedDirect Link - Cloutman, D.G., 1975. Parasite community structure of largemouth bass, warmouth and bluegill in Lake FortSmith, Arkansas. Trans. Am. Fish. Soc., 104: 277-283.
CrossRefDirect Link - Cotgreave, P., 1993. The relationship between body size and population abundance in animals. Trends Ecol. Evol., 8: 244-248.
CrossRef - Crompton, D.W., 1973. The sites occupied by same parasitic helminths in the alimentary tract of vertebrates. Biol. Rev., 48: 27-83.
PubMed - Dubey, A., A.K. Gupta and S.M. Agarwal, 1992. Studies on monogenean parasites of freshwater fishes at Raipur X. Taxonomic discussion on the validity of the genus Urocleidus (Muller, 1934) Muller, 1936 (Ancyrocephalinae) and description of a new species from Mastacembelus armatus and Macrognathus aculeatus at Raipur. Indian J. Helminthol., 44: 116-122.
- Esch, G.W., A.W. Shostak, D.J. Marcogliese and T.M. Goater, 1990. Patterns and Processes in Helminth Parasite Communities: An Overview. In: Parasite Communities: Patterns and Processes, Esch, G.W., A.O. Bush and J.M. Aho (Eds.). Chapman and Hall, London, UK., pp: 1-20.
Direct Link - Fiorillo, R.A. and W.F. Font, 1996. Helminth community structure of four species of Lepomis (Osteichthyes: Centrarchidae) from an oligohaline estuary in South Eastern Louisiana. J. Helminthol. Soc. Washington, 63: 24-30.
Direct Link - Folstad, I. and A.J. Karter, 1992. Parasites, bright males and the immunocompetence handicap. Am. Naturalist, 139: 603-622.
Direct Link - Guidelli, G.M., A. Isaac, R.M. Takemoto and G.C. Pavanelli, 2003. Endoparasite infracommunities of Hemisorubim platyrhynchos (Valenciennes, 1840) (Pisces: Pimelodidae) of the Baia river, upper Parana river floodplain, Brazil: Specific composition and ecological aspects. Brazilian J. Biol., 63: 261-268.
PubMed - Hanek, G. and C.H. Fernando, 1978. The role of season, habitat, host age and sex on gill parasites of Lepomis gibbous (L.). Can. J. Zool., 56: 1247-1250.
CrossRefDirect Link - Hanek, G. and C.H. Fernando, 1978. The role of season, habitat, host age and sex on gill parasites of Ambloplites rupestris (Raf.). Can. J. Zool., 56: 1251-1253.
CrossRefDirect Link - Harshey, K.R., 1937. On two new trematodes of the genus Opegaster Ozaki, with a systematic discussion on the families Opecoelidae Ozaki, 1925 and Coitocaecidae Ozaki, 1928. Proc. Ind. Aca. Sci., 5: 64-75.
CrossRef - Hatcher, M.J., J.T.A. Dick and A.M. Dunn, 2006. How parasites affect interactions between competitors and predators. Ecol. Lett., 9: 1253-1271.
PubMed - Hernandez, A.D., J.F. Bunnell and M.V. Sukhdeo, 2007. Composition and diversity patterns in metazoan parasite communities and anthropogenic disturbance in stream ecosystem. Parasitology, 134: 91-102.
PubMedDirect Link - Holmes, J.C., 1973. Site selection by parasitic helminths: Interspecific interactions, site segregation and their importance to the development of helminth communities. Can. J. Zool., 51: 333-347.
PubMedDirect Link - Holmes, J.C., 1990. Competition, contacts and other factors restricting niches of parasitic helminths. Ann. Parasitol. Hum. Comp., 65: 69-72.
CrossRefPubMedDirect Link - Jain, S.L., 1958. Monogenea of freshwater fishes. VII. Bifurcohaptor, a new genus from the gill filaments of two fishes from Lucknow. J. Parasitol., 44: 388-394.
Direct Link - Jain, S.L., 1959. Monogenea of Indian freshwater fishes VIII. Sprostonia, a new genus of freshwater Tetraonchinae with the descriptions of four new species, from the gills filaments of fishes, from Lucknow. Parasitology, 49: 153-168.
PubMed - Jalali, B. and M. Barzegar, 2006. Fish parasites of Zarivar Lake. J. Agric. Sci. Technol., 8: 47-58.
Direct Link - Johnson, M.W., P.A. Nelson and T.A. Dick, 2004. Structuring mechanisms of yellow perch (Perca flavescens) parasite communities: Host age, diet and local factors. Can. J. Zool., 82: 1291-1301.
Direct Link - Kennedy, C.R. and A.O. Bush, 1994. The relationship between pattern and scale in parasite communities: A stranger in a strange land. Parasitology, 109: 187-196.
PubMedDirect Link - Kennedy, C.R., A.O. Bush and J.M. Aho, 1986. Patterns in helminth communities: Why are birds and fish different? Parasitology, 93: 205-215.
PubMed - Kritsky, D.C., K.C. Pandey, N. Agrawal and S.M.A. Abdullah, 2004. Monogenoids from the gills of spiny eels (Teleostei: Mastacembelidae) in India and Iraq, proposal of Mastacembelocleidus gen. n. and status of Indian species of Actinocleidus, Urocleidus and Haplocleidus (Monogeneoidea: Dactylogyridae). Folia Parasitol., 51: 291-298.
- Kue-A-Pai, P. and V. Wiwanikit, 2005. Contamination of the Gnathostoma spp. Infective stage larvae in Fluta alba sold in Bangkok. Internet Trop. Med., 2: 3-3.
Direct Link - Lawrence, J.L., 1970. Effects of season, host age and sex on endohelminths of Catostomi commersoni. J. Parasitol., 56: 567-571.
CrossRefDirect Link - Llewellyn, J., 1962. The life histories and population dynamics of monogenean gill parasites of Trachurus trachurus (L.). J. Mar. Biol. Assoc, U.K., 42: 587-600.
CrossRefDirect Link - Luque, J.L. and R. Poulin, 2004. Use of fish as intermediate hosts by helminth parasites: A comparative analysis. Acta Parasitol., 49: 353-361.
Direct Link - Luque, J.L., J.F.R. Amato and R.M. Takemoto, 1996. Comparative analysis of the communities of metazoan parasites of Orthopristis rubber and Haemulon steindachneri (Osteichthyes: Haemulidae) from the Southeastern Brazilian littoral: I. Structure and influence of size and sex of the hosts. Revista Brasileira Biologia, 56: 279-292.
- Luque, J.L., J.F.R. Amato and R.M. Takemoto, 1996. Comparative analysis of the communities of metazoan parasites of Orthopristis rubber and Haemulon steindachneri (Osteichthyes: Haemulidae) from the Southeastern Brazilian littoral: II. Diversity, interspecific assosciations and distribution of gastrointestinal parasites. Revista Brasileira Biologia, 56: 293-302.
- Malan, F.S., I.G. Horak, V. de Vos and J.A. van Wyk, 1997. Wildlife parasites: Lessons for parasite control in livestock. Vet. Parasitol., 71: 137-153.
CrossRef - Marcogliese, D.J., 2004. Parasites: Small players with crucial roles in the ecological theater. Ecohealth, 1: 151-164.
CrossRefDirect Link - Margolis, L., G.W. Esch, J.C. Holmes, A.M. Kuris and G.A. Shad, 1982. The use of ecological terms in parasitology (Report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol., 68: 131-133.
CrossRefDirect Link - McVicar, A.H., 1977. Intestinal helminth parasites of ray, Raja naevus in British waters. J. Helminthol., 51: 11-21.
PubMedDirect Link - Molloy, S., C. Holland and R. Poole, 1995. Metazoan parasite community structure in brown trout from two lakes in western Ireland. J. Helminthol., 69: 237-242.
PubMedDirect Link - Munoz, G., A.S. Grutter and T.H. Cribb, 2007. Structure of the parasite communities of a coral reef fish assemblage (Labridae): Testing ecological and phylogenetic host factors. Int. J. Parasitol., 93: 17-30.
PubMed - Muzzall, P.M., 1980. Population biology and host-parasite relationships of Triganodistomum attenuatum (Trematoda: Lissorchiidae) infecting the white sucker, Catostomus commersoni (Lacepede). J. Parasitol., 66: 293-298.
Direct Link - Pearson, J.C., 1968. Observations on the morphology and life-cycle of Paucivitellosus fragilis Coil, Reid and Kuntz, 1965 (Trematoda: Bivesiculidae). Parasitology, 58: 769-788.
CrossRef - Pennycuick, L., 1971. Differences in the parasite infections in three-spined sticklebacks (Gasterosteus aculeatus L.) of different sex, age and size. Parasitology, 63: 407-418.
CrossRefDirect Link - Poulin, R., 1995. Phylogeny, ecology and richness of parasite communities in vertebrates. Ecol. Monographs, 65: 283-302.
Direct Link - Poulin, R., 1996. Sexual inequalities in helminth infections: A cost of being male? Am. Nat., 147: 287-295.
Direct Link - Roubal, F.R., 1990. Seasonal changes in ectoparasite infection of juvenile yellowfin bream, Acanthopagrus australis (Gunther) (Pisces: Sparidae), from a small estuary in Northern New South Wales. Aust. J. Mar. Freshwter Res., 41: 411-427.
CrossRefDirect Link - Sasal, P., S. Morand and J.F. Guegan, 1997. Determinants of parasite species richness in Mediterranean marine fish. Mar. Ecol. Prog. Ser., 149: 61-71.
Direct Link - Shotter, R.A., 1973. Changes in the parasite fauna of whiting Odontogadus merlangus L. with age and sex of the host, season and from different areas in the vicinity of the isle of man. J. Fish. Biol., 5: 559-573.
CrossRef - Sousa, W.P., 1994. Patterns and processes in communities of heirninth parasites. Trends Ecol. Evol., 9: 52-57.
CrossRefDirect Link - Sukontason, K., S. Piangjai, Y. Muangyimpong, K. Sukontason, R. Methanitikorn and U. Chaithong, 1999. Prevalence of trematode metacercariae in cyprinoid fish of Ban Pao district, Chiang Mai province northern Thailand. Southeast. Asian J Trop Med Public Health, 30: 365-370.
PubMedDirect Link - Takemoto, R.M. and G.C. Pavanelli, 2000. Aspects of the ecology of proteocephalid cestodes parasites of Sorubim lima (Pimelodidae) of the upper Parana river, Brazil: I. Structure and influence of host's size and sex. Braz. J. Biol., 60: 577-584.
PubMedDirect Link - Thomas, J.D., 1964. A comparison between the helminth burdens of male and female brown trout, Salmo trutta (L.) from a natural population in River Teify, West Wales. Parasitology, 54: 263-272.
CrossRefDirect Link - Vignon, M. and P. Sasal, 2010. Fish introduction and parasites in marine ecosystems: A need for information. Environ. Biol. Fish., 87: 1-8.
CrossRef - Wongsawad, C., A. Rojanapaibul, P. Wongsawad, A. Rojanapaibul and T. Marayong et al., 2004. Helminths of vertebrates in Mae Sa Stream, Chiang Mai, Thailand. Southeast Asian J. Trop. Med. Public Health, 35: 140-146.
Direct Link