
A.M. Al- Attar
Department of Biological Sciences, Faculty of Sciences,King Abdul Aziz University, P.O. Box 139109, Jeddah 21323, Saudi Arabia
International Journal of Zoological Research
Year: 2010 | Volume: 6 | Issue: 3 | Page No.: 199-213







ABSTRACT
The present study was aimed to evaluate the impact of Waltonella duboisi naturally infection in the marsh frog, Rana ridibunda. Healthy and infected frogs of both sexes were collected from Al-Qatif and Al-Hassa farms, Eastern Province, Saudi Arabia. The hematological, biochemical and histopathological changes were estimated in infected male and female frogs compared with healthy frogs. The values of red blood cell count, hemoglobin concentration, hematocrit, mean cell volume, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration and white blood cell count were statistically decreased in infected frogs. Infection with Waltonella duboisi induced significant reduction in the levels of serum glucose and total proteins, while the values of triglycerides cholesterol, creatinine, glutamic pyruvic acid transaminase and glutamic oxaloacetic acid transaminase were significantly elevated. Histopathological examination of stomach, small intestine, liver showed the larval developmental stages of Waltonella duboisi. A partially abnormal of testis and ovary structures with pronounced disturbance in quantity and quality of spermatogenesis and oogenesis processes were noted in infected of both sexes of frogs. From the present study, it is obviously that Waltonella duboisi caused many severe physiological and histopathological alterations in both sexes of the marsh frogs. Thus, more sincere ecological and scientific efforts are required to rescue the marsh frog population from parasitic infection, pathogenic factors and increases of mortality rate.
PDF Abstract XML References Citation
Received: November 17, 2009;
Accepted: March 29, 2010;
Published: May 24, 2010
How to cite this article
A.M. Al- Attar, 2010. Hematological, Biochemical and Histopathological Studies on Marsh Frog, Rana ridibunda, Naturally Infected with Waltonella duboisi. International Journal of Zoological Research, 6: 199-213.
DOI: 10.3923/ijzr.2010.199.213
URL: https://scialert.net/abstract/?doi=ijzr.2010.199.213
DOI: 10.3923/ijzr.2010.199.213
URL: https://scialert.net/abstract/?doi=ijzr.2010.199.213
INTRODUCTION
Pathogens and parasites have a variety of deleterious effects on individual hosts, including decreased growth, reduced development and behavioral modifications that may ultimately result in increased mortality (Dobson and Hudson, 1986; Minchella and Scott, 1991; Dobson and Crawley, 1994; Poulin, 1994; Seppala et al., 2004; Ponton et al., 2009). Parasites may also affect the size of host populations by reducing rates of host reproduction and by increasing rates of host mortality (Anderson, 1978; Anderson and May, 1978; Hudson et al., 1998; Tompkins and Begon, 1999; Huber, 2008; Seppala et al., 2008). Parasites can also alter host morphology (Despommier, 1990; Krist, 2000; Moreels et al., 2004). The level of modification ranges from intracellular to the whole organism. Nevertheless, it has been hypothesized that the impact of parasitic infection on host populations maybe strongly context dependent (Gulland, 1995). The impact of infection on the host will depend upon the virulence of the parasite, the rate of infection and the resistance of the host to infection.
These parameters can be modified by a number of factors, such as overcrowding and nutritional stress, that also can interact with each other. In fact, previous studies suggest that the degree of differential mortality suffered by infected hosts depends not only on the particular host–parasite association under study, but also on the kind and level of a biotic stress experienced by the host (Esch et al., 1975; Camp et al., 1982; Sousa and Gleason, 1989). Thus, predicting the impact of a particular pathogen on host population dynamics will depend on the context of the host-pathogen interaction (Kiesecker and Skelly, 2001).
Nematodes or roundworms inhabit virtually all ecosystems including marine, freshwater and terrestrial environments. They encompass the phylum Nematoda (Nemata) that includes plant, insect, animal and human parasites and predatory forms that consume microorganisms including bacteria, fungi, algae and other nematodes (Platt, 1994). Waltonella duboisi is the member of Onchocercidae family (Phylum: Nematoda, Class: Chromadorea, Order: Rhabdititda). The family of Onchocercidae includes some of the most devastating human parasitic diseases, such as lymphatic filariasis, onchocerciasis, loiassis and other filariasis. Onchocercidae is expanded to include Setariinae and the Waltonllinae subfamilies. The subfamily of Waltonllinae, proposed by Bain and Prod’hon (1974), is restricted to amphibians and consists of Waltonella, Madochoteva and Ochoterenella.
The decline of world populations of amphibians is a major environmental issue (Vertucci and Corn, 1996). Amphibians are an integral part of their ecosystems; affecting nutrient cycling and also serving as high quality prey for many species (De Maynadier and Hunter, 1995). In the last 15 years, scientists have accumulated evidence supporting a global decline in amphibians. As the quantitative evidence grows, it is difficult to deny the validity of this global trend (Houlahan et al., 2000; Stuart et al., 2004). Amphibians are especially at risk from agricultural contamination because they have permeable skin and eggs that readily absorb chemicals from the environment. Unable to be ignored, amphibians are undergoing a global decline. The magnitude of this phenomenon first became apparent in 1990 when overall amphibian population numbers began experiencing significant decreases (Collins and Storfer, 2003; Stuart et al., 2004; Pounds et al., 2006; Lips et al., 2006). Since, then, much research has been done on the subject. Amphibians are often seen as indicator species for habitat degradation and declines in abundance can be regarded as indicative of the future health of their ecosystem. Several causes for these declines have been proposed-including such things as deforestation, invasive species and pollution-although, in many cases, the cause is unclear. Other studies of overall declines around the globe have eliminated natural population fluctuations and metapopulation activities as the causes of the increased mortality rates and loss of populations (Daszak et al., 1999). While the causes of many declines remain a mystery, there are many explanations known to negatively impact species' numbers and many more hypotheses to help explain this trend. Collins and Storfer (2003) divided the potential causes for decline into two categories which include the long standing hypotheses with which we have a fairly strong understanding (e.g., habitat destruction) and the less understood or enigmatic hypotheses (e.g., disease) which include influences more relevant in today's world than before. Common and obvious causes that have long been known to have a hostile affect on any species include such things as habitat fragmentation, overexploitation, habitat destruction and introduction of invasive species. Other factors might be just as important, but currently their role is unclear. These proposed causes include global warming and increased UV radiation, the presence of emerging infectious diseases and the use and spread of environmental contaminants (Collins and Storfer, 2003). The latter two conditions, disease and contaminants, are presently ranked as two of the leading hypotheses for the global amphibian decline (Collins and Storfer, 2003; Storfer, 2003; Stuart et al., 2004).
Rana ridibunda (marsh frog) is a highly opportunistic amphibian. It lives in mixed and deciduous forests, forest steppe, steppe, semidesert and desert zones. Being a semiaquatic species, the frog inhabits a wide variety of flowing and stagnant water habitats, from shallow puddles and ponds to large lakes and rivers, as well as mountain streams. The marsh frog is the only one true frog reported from the oasis of Al-Hassa and Al-Qatif in Eastern Province of Saudi Arabia (Schmidt, 1953; Hass, 1957, 1961; Briggs, 1980). The frog has been subject of study to determine the seasonal population structure, breeding, behavior, tadpole development and individual growth in Saudi climate (Briggs, 1981). The objective of the present study was to evaluate the hematological, biochemical and histopathological changes in the marsh frog, Rana ridibunda, naturally infected with a nematode, Waltonella duboisi.
MATERIALS AND METHODS
Animals
Fifty healthy and sixty four infected adult frogs, Rana ridibunda, of both sexes (27.4-31.6 g) were separately collected from Al-Qatif and Al-Hassa farms, Eastern Province, Saudi Arabia. The collected frogs were placed in plastic containers and transported to the laboratory (temperature: 24±1°C). Healthy and infected frogs were separately kept in glass aquaria and fasted for one day before experimental processes.
Hematological and Biochemical Determinations
After one day, blood specimens were withdrawn from the frogs by cardiac puncture. The body cavity was then dissected and the sex was determined from the presence of testes or ovaries. Blood were collected in tubes with and without anticoagulant. Lithium heparin was used as anticoagulant. The blood in lithium heparin tubes were immediately used for hematological measurements including Red Blood Cell (RBC) count, hemoglobin (Hb) concentration, hematocrit (Hct) value, Mean Cell Volume (MCV), Mean Corpuscular Hemoglobin (MCH), Mean Corpuscular Hemoglobin Concentration (MCHC) and White Blood Cell (WBC) count. The RBC count was estimated according to the classical method using Neubauer hemocytometer. The Hb concentration was measured by blood hemoglobin photometer system (HemoCue AB, Angelholm, Sweden). A microcapillary reader was used to determine the value of Hct. The values of MCV, MCH and MCHC were calculated using standard equations (Platt, 1969). Total WBC were counted according the method of Shaw (1930). For serum biochemical measurements, blood in tubes without anticoagulant were left for clotting and serum separation. These specimens were centrifuged at 1500 rpm for 10 min and blood sera were then collected and stored at 4°C till the determination time of glucose, total protein, triglycerides, cholesterol, creatinine, Glutamic Pyruvic acid Transaminase (GPT) and Glutamic Oxaloacetic acid Transaminase (GOT). All of these biochemical parameters were analyzed using an automatic analyzer (BM/Hitachi System 717).
Histopathological Examination
Stomach, small intestine, liver, testis and ovary tissues from each groups were fixed with 10% buffered formalin. Fixed tissues were dehydrated through ascending grades of ethanol to absolute ethanol. They were cleaned in xylene, impregnated and embedded in paraffin wax (melting point 56°C). Sections were cut at 4 μm on a rotary microtome. They were flattened on warm water and mounted onto albumerised slides and dried overnight. The sections were dewaxed in xylene and hydrated through descending grades of ethanol to water. They were initially stained in Harris haematoxylin and differentiated in acid alcohol and thereafter stained with methylene blue. They were then dehydrated in 95% alcohol, stained in 10% alcohol eosin, dehydrated in absolute alcohol, cleaned in xylene and mounted in Canada balsam. The resulting slides are then viewed under the light microscope.
Statistical Analysis
Hematological and biochemical results were expressed as means of eight determinations±standard deviation (SD). Statistical analysis was performed as healthy group versus infected group using the Student’s t-test. The chosen level of significance is p<0.05.
RESULTS
The effects of Waltonella duboisi infection on hematological parameters in male and female frogs are shown in Fig. 1a-g. In comparison with healthy male and female frogs, the infection with Waltonella duboisi induced significant reduction in RBC count of male (-20.88%) and female (-30.07%) frogs. Hb (-30.83% and -42.04%), Hct (-30.06% and -36.91%), MCV (-9.40% and -8.45%) and MCH (-9.57% and -17.66%) levels and WBC (-21.98% and -24.38%) count were statistically declined in infected male and female frogs. The level of MCHC (-8.04%) was decreased only in infected female. Serum biochemical values are shown in Fig. 2a-g.
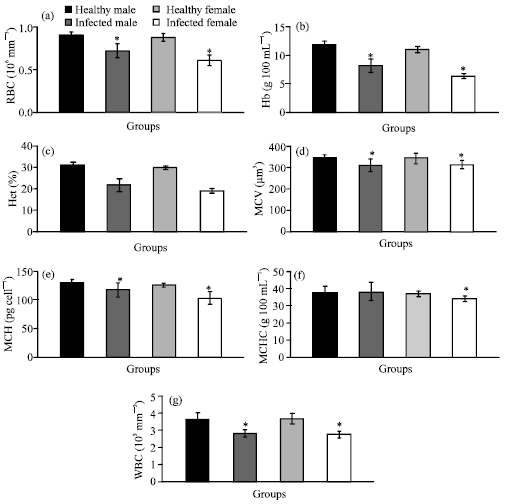 | |
| Fig. 1: | (a-g) The values of hematological parameters of healthy male, infected male, healthy female and infected female Rana ridibunda. *p<0.05 significant vs. healthy group |
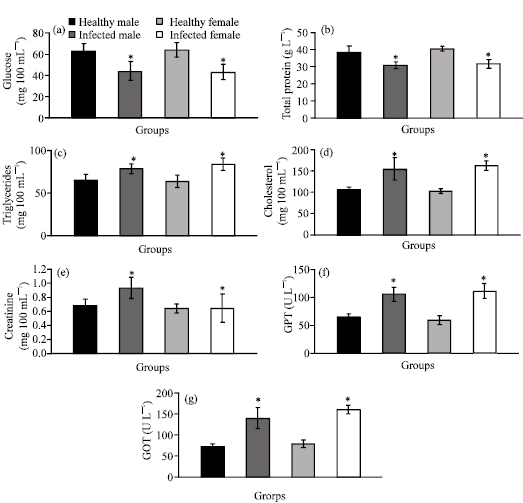 | |
| Fig. 2: | (a-g) The values of serum glucose, total protein, triglycerides, cholesterol, creatinine, GPT and GOT of healthy male, infected male, healthy female and infected female Rana ridibunda. *p<0.05 significant vs. healthy group |
The values of serum glucose (-29.32% and -32.61%) and total protein (-19.54% and -21.37%) were markedly decreased, while the values of serum triglycerides (+21.05% and +31.23%), cholesterol (+45.79% and +58.56%), creatinine (+36.77% and +50.00%), GPT (+62.42% and +88.09) and GOT (+91.72% and +103.01) were significantly increased in male and female frogs infected with Waltonella duboisi. Moreover, it is pronounced from these results that the infected female frogs were more affected than male frogs.
Normal histology of stomach in healthy frogs was shown in Fig. 3a and b. These figures represented a typical structure of stomach tissues including serosa, muscularis, submucosa and mucosa layers. Figure 3c showed the primary of developmental stage of Waltonella duboisi larva in submucosa layer with pronounced cyst formation. Advanced developmental stages of larvae were noted in muscularis layer (Fig. 3d-f). Figure 3g and h showed the final stages of larval development in muscularis layer of stomach. Figure 3i showed leucocytic infiltrations in the area of larval development. Also, necrotic areas were observed in muscularis layer (Fig. 3j). Figure 4a demonstrated the normal structure of small intestine of healthy frogs including normal serosa, muscularis, submucosa and mucosa layers. The developmental stages of Waltonella duboisi larvae were located in muscularis and submucosa layers of small intestine (Fig. 4b-f). Figure 5a shows a typical structure of liver.
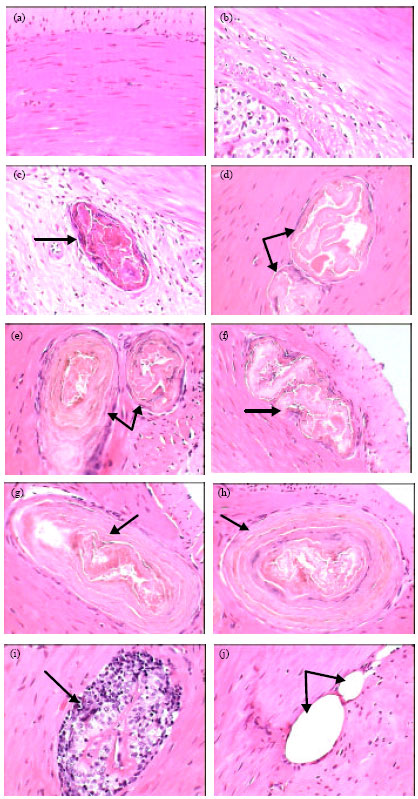 | |
| Fig. 3: | Photomicrographs of stomach of healthy frogs (a and b) showing normal structure and infected frogs (c-h) showing the larval developmental stages of Waltonella duboisi in muscularis and submucosa layers (arrows). Leucocytic infiltration (i) and necrotic areas (j) are also visible (arrows). Original magnification X400 |
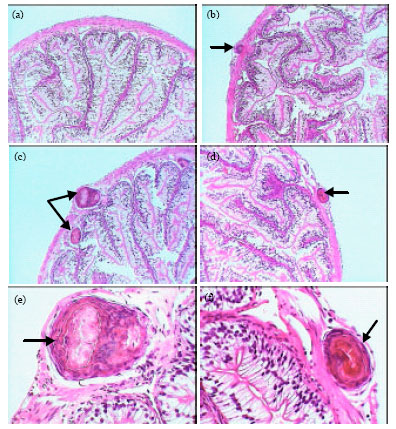 | |
| Fig. 4: | Photomicrographs of small intestine of healthy frogs (a, X100) showing normal structure and infected frogs (b, c and d X100; e and f X400) showing the larval developmental stages of Waltonella duboisi in muscularis and submucosa layers (arrows) |
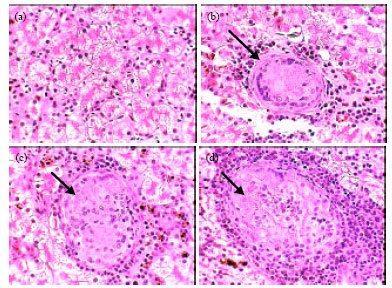 | |
| Fig. 5: | Photomicrographs of liver of healthy frogs (a) showing normal structure and infected frogs (b, c and d) showing the larval developmental stages of Waltonella duboisi in liver tissue (arrows) with appearance of leucocytic infiltrations. Original magnification X400 |
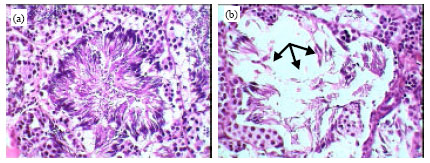 | |
| Fig. 6: | Photomicrographs of testis seminiferous tubules of healthy frogs (a) showing normal structure and infected frogs, (b) showing a partially abnormal of seminiferous tubules structure with pronounced disturbance in quantity and quality of spermatogenesis process (arrows). Original magnification X400 |
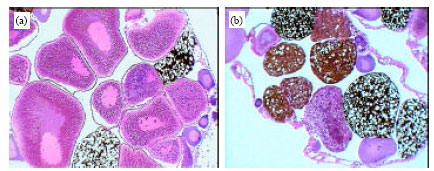 | |
| Fig. 7: | Photomicrographs of ovary of healthy frogs (a) showing a typical structure with normal oogenesis process and infected frogs, (b) showing a partially abnormal of structure with pronounced disturbance in quantity and quality of oogenesis process. Original magnification X100 |
The liver consists of a large number of acini which appear in cross-sections lined with granular columnar hepatocyets. The hepatocytes are surround a bile ductule in the center. Some of the hepatocytes contain pigment granules. Among the acini are found the blood sinusoids and capillaries. Figure 5b-d showed the developmental stages of Waltonella duboisi larvae in liver tissue with appearance of leucocytic infiltrations. The normal structures of testis seminiferous tubules and spermatogenesis process in healthy frogs are shown in Fig. 6a. Figure 6b showed a partially abnormal of seminiferous tubules structure in infected male with pronounced disturbance in quantity and quality of spermatogenesis process. Figure 7a showed a typical structure of ovary in healthy frogs including the normal oogenesis process. Figure 7b shows the structure of ovary in infected frogs. A partially abnormal structure and increases of adipose tissue were observed in ovary of infected frogs (Fig. 7b).
DISCUSSION
It is a first report to study the hematological, biochemical and histopathological alterations in the marsh frog of Eastern Province, Saudi Arabia which naturally infected with a nematode, Waltonella duboisi. Al Saqabi (2006) surveyed the intestinal and blood parasites from Rana ridibunda. He found seven species and genera of parasites belonging to Protozoa, Nematoda and Digenea. Protozoa were found in 91.4% of the frogs, Nematoda in 23.7% and Digenea in 1.7% of the population. The parasitic infection was seen more in females than males. The following parasites were recovered and described; from the Protozoa Balantidium duodeni, Nyctotherus cordiformis, Oplina sp. From the Nematoda (Waltonella duboisi) and from the Digenea the genus Pleurogenes was described. In addition, Trypanosoma and Microfilaria were found in the blood of some frogs 2.8 and 1.1%, respectively.
The present histopathological examination showed that larva of Waltonella duboisi infects many organs in frogs including stomach, small intestine and liver. The developmental stages of larva caused several alterations in tissues structure of infected organs. These histopathlogical alterations induced sever changes in hematological and biochemical parameters. Waltonella duboisi caused significant decreases in hematological parameters of frogs such as RBC, Hb, Hct, MCV, MCH and MCHC values. These results are in accordance with the results indicating a disruption of erythropoiesis or an increase in destruction of blood cells. Also, The hematological values in this study concurs with those of Kozat et al. (2006), Gadre et al. (2008) and Mbaya et al. (2009), where they reported significant decrease in RBC, Hb, Hct, MCV, MCH and MCHC in affected animal with gastrointestinal helminthes. It has been established that the measurement of anaemia gives a reliable indication of the disease status and productive performance of parasites infected human and animals (Haroun et al., 1996; Tsuyuoka et al., 1999; Adogwa et al., 2005; Raji et al., 2006; Mir et al., 2007). Contrary to the present finding of WBC reduction, several studies showed higher WBC values (leukocytosis) in human and animals infected with different gastrointestinal nematodes (Haroun et al., 1996; Intapan et al., 2008; Al-Bayati and Arsalan, 2009; Bernard et al., 2009). Also, the present reduction of WBC count indicates immunodepression induction in Waltonella duboisi-infected frogs.
It is well known that parasites influences, alter and interfere with normal physiology of the host, to adjust their establishment and living in the body of the host. Serum glucose and total protein levels were significantly decreased in frogs infected with Waltonella duboisi. These results obviously indicate disturbances in carbohydrate and protein metabolism. In general, nematodes infections have been shown to reduce the ability of intestine mucosa to transport a wide range of metbolites (Sukhdeo and Mettrick, 1984). Several studies showed a decrease in the level of serum protein in animals infected with gastrointestinal nematodes (Konx et al., 1993; Haroun et al., 1996; Mir et al., 2007). On the other hand, significant increases of the triglyceride and cholesterol levels were recorded in Waltonella duboisi infested frogs. This is may be attributed to the increased rate of lipolysis as an alternative energy source due to the inadequate carbohydrate absorption in parasitized frogs and the shortage of glycogen storage. The increased lipolysis results in liberation of triglyceride and non-esterified fatty acids which is processed by the liver into acetylCoA in the tricarboxylic acid (TCA) cycle for energy production. Mukerjee et al. (1990) reported that an infection of with Ancylostoma ceylanicum induced profound hyperlipidemia in golden hamsters, particularly hypertriglyceridemia and the effect was directly related to the degree of infection. A significant increase was also noticed in serum cholesterol and phospholipid levels. The hyperlipidemia was further characterized by an increase in Low Density Lipoproteins (LDL) and Very Low Density Lipoproteins (VLDL) with a concomitant decline in High Density Lipoproteins (HDL). Decreased lipolytic activities, especially triglyceride lipase, in hepatic tissue and induction of lipolytic activities in intestine and adipose tissues indicated mobilization of fats from adipose and jejunum with a defective removal of triglyceride-rich lipoproteins in hepatic tissues. Also, they demonstrated that the accumulation of lipids in liver and depletion in adipose tissue supported their results. The derangement may have a significant effect on host parasite interaction and is an important pathophysiological feature occurring during experimental ancylostomiasis.
The obtained results revealed significant increases in serum creatinine, GPT and GOT levels in frogs infected with Waltonella duboisi. Creatine is synthesized in the liver, passes into circulation and is taken up almost entirely by skeletal muscle for conversion to creatine phosphate, which acts as an energy reservoir. Creatine and its phosphate are converted spontaneously into creatinine (McLauchlan, 1988). The two substances are handled differently by the kidney. Both are filtered at glomerulus. Although there may be some additional secretion of creatinine by renal tubules, creatine is reabsorbed by the tubules at low plasma concentration. This ensures that there is little, or no creatine in urine (McLauchlan, 1988). The heightened appearance of creatinine in the serum of Waltonella duboisi-infected frogs indicates the increased transformation of phosphocreatine to creatinine in muscle which might be due to lesser utilization of phosphocreatine in muscular contraction. Thus significant increase in creatinine concentration in serum could be due to increased release from muscles and/or decrease excretion from the kidney. This suggests that infection with Waltonella duboisi causes adverse changes in skeletal muscle and kidney. It was reported that hepatocellular damage could be correlated with the disturbed enzyme activities. Martin et al. (1983) announced that liver tissues, which are known for their high content of transaminases (GPT and GOT) lose their enzymes in case of liver cell damage. This ultimately leads to their raised levels in the sera. Hence, they suggested that the higher values of these enzymes, whenever they are detected in the blood sera, should be taken as indicator of various causes of liver damage.
Unlike microparasites, such as viruses, bacteria and protozoa and unlike intracellular parasites, sexual reproduction of gastrointestinal nematodes leads to release of eggs through urine or feces into the environment. These eggs or resulting larval transmission stages reinfect the host by oral ingestion or by skin penetration. The most striking features of many gastrointestinal nematodes are their long-term persistence (1-10 years) within the host despite immune recognition, their ability to evade host defenses and their ability to continually reinfect the host, all of which contribute to their classification as long-lasting, chronic infections that more often cause morbidity rather than mortality. In children, long-term infection, if associated with poor nutrition, impairs physical and cognitive development (Nokes et al., 1992; Hadju et al., 1995; Savioli et al., 2000). In human communities where a minority of people harbors the majority of worms, it is believed that the few individuals predisposed to heavy infections provide a continual source of infection in the community (Maizels et al., 1993; Peng et al., 1998). It is suspected that these are the most malnourished individuals, although genetic and behavioral factors are also believed to play a role in predisposing individuals to heavy infection (Holland et al., 1992; Maizels et al., 1993). The first challenge for any parasite is to invade its host and to migrate to its final destination, a process that involves traversing host tissue, extracellular matrices, basement membranes and blood or lymph vessel walls. Many parasites, including nematodes, rely on their wide array of specific proteinases (Tort et al., 1999), released as excretory/secretory (E/S) products, to accomplish this task. These proteinases, which can be released by penetrating larval stages or by adult worms, hydrolyze fibrinogen, hemoglobin and collagen. Although, peptidases of the four major groups (serine, aspartic, cysteine and metalloproteinases) have been described for gastrointestinal nematodes (Hill et al., 1994; Gamble et al., 1996; Rehman and Jasmer, 1998; Tort et al., 1999), a large number belong to the papain superfamily of cysteine proteinases (i.e., cathepsin B) and the zinc metalloproteinases (Gamble et al., 1989, 1996) and there is some evidence, as least for Ascaris that if release of larval proteinases is inhibited, nematode migration may be prevented (Knox and Kennedy, 1988).
Global declines in amphibian populations have caused amphibians to become the most threatened class of vertebrates. Since the late 1980s there has been a growing realization that populations of many amphibian species from a variety of taxa are declining at an alarming rate. Recent reports suggest that the frequency of mass incidences of frog deformities has increased and population declines have been observed within the last two decades (Blaustein and Wake, 1990, 1995; Bishop and Pettit, 1992; Materna et al., 1995; Houlahan et al., 2000; Gardner, 2001; Blaustein et al., 2003). To prevent further decline it is very important to establish what factors are responsible. While evidence of widespread reductions in the numbers of amphibians is rapidly accumulating, the cause remains unclear. Postulated causes of the deformities and population declines include ultraviolet radiation (Blaustein et al., 2003), parasites (Rollins-Smith et al., 2002a, b), agrochemicals, especially pesticides (Ouellet et al., 1997; Sparling et al., 2001) and infectious microbial disease (Bradford, 1991; Carey, 1993; Lips, 1998). From the present study, it is obviously that Waltonella duboisi caused many severe physiological and histopathological alterations in both sexes of marsh frogs. This infection may lead to increase the mortality rate and decline the population rate of marsh frogs. According to the current research, it is highly recommended and also requested to urgently increase our efforts using ecological and scientific strategies to rescue the infected marsh frogs.
REFERENCES
- Adogwa, A., A. Mutani, A. Ramnanan and C. Ezeokoli, 2005. The effect of gastrointestinal parasitism on blood copper and hemoglobin levels in sheep. Can. Vet. J., 46: 1017-1021.
PubMedDirect Link - Al-Bayati, O.A.S. and S.H. Arsalan, 2009. Clinical and hematological study in sheep infected with gastrointestinal parasites in Mosul. Iraqi J. Vet. Sci., 23: 93-100.
Direct Link - Al-Saqabi, S.M., 2006. Some intestinal and blood parasites infection in the frog Rana ridibunda in Saudi Arabia. Arab Gulf J. Sci. Res., 24: 151-159.
Direct Link - Anderson, R.M. and R.M. May, 1978. Regulation and stability of host-parasite population interactions: I. Regulatory processes. J. Anim. Ecol., 47: 219-247.
CrossRefDirect Link - Bernard, G., M. Worku and M. Ahmedna, 2009. The effects of Diatomaceous earth on parasite infected goats. Bull. Georgian Natl. Acad. Sci., 3: 129-135.
Direct Link - Blaustein, A.R. and D.B. Wake, 1995. The puzzle of declining amphibian populations. Sci. Am., 272: 52-63.
Direct Link - Blaustein, A.R. and D.B. Wake, 1990. Declining amphibian populations: A global phenomenon. Trends Ecol. Evol., 5: 203-204.
Direct Link - Blaustein, A.R., J.M. Romansic, J.M. Kiesecker and A.C. Hatch, 2003. Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers. Distrib., 9: 123-140.
Direct Link - Bradford, D.F., 1991. Mass mortality and extinction in a high-elevation population of Rana muscosa. J. Herpetol., 25: 174-177.
Direct Link - Carey, C.L., 1993. Hypothesis concerning the causes of the disappearance of boreal toads from the mountains of Colorado. Conserv. Biol., 7: 355-362.
CrossRefDirect Link - Collins, J.P. and A. Storfer, 2003. Global amphibian declines: Sorting the hypothesis. Diversity Distributions, 9: 89-98.
Direct Link - Daszak, P., L. Berger, A.A. Cunningham, A.D. Hyatt, D.E. Green and R. Speare, 1999. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis., 5: 735-748.
PubMedDirect Link - Despommier, D.D., 1990. Trichinella spiralis-the worm that would be virus. Parasitol. Today, 6: 193-196.
PubMedDirect Link - Dobson, A. and M. Crawley, 1994. Pathogens and the structure of plant communities. Trends Ecol. Evol., 9: 393-398.
CrossRef - Dobson, A.P. and P.J. Hudson, 1986. Parasites, disease and the structure of ecological communities. Trends Ecol. Evol., 1: 11-15.
CrossRef - Esch, G.W., J.W. Gibbons and J.E. Bourque, 1975. An analysis of the relationship between stress and parasitism. Am. Midland Naturalist, 93: 339-353.
Direct Link - Gadre, A.S., D.K. Maske, C.G. Panchbhai, T.R. Gawande, S.W. Kolte and A.R. Sirothia, 2008. Haematological changes in naturally infested dairy animal at central zone of vidarbha. Vet. World, 1: 47-48.
Direct Link - Gamble, H.R., R.H. Fetterer and L.S. Mansfield, 1996. Developmentally regulated zinc metalloproteinases from third- and fourth-stage larvae of the ovine nematode Haemonchus contortus. J. Parasitol., 82: 197-202.
Direct Link - Gamble, H.R., J.P. Purcell and R.H. Fetterer, 1989. Purification of a 44 kilodalton protease which mediates the ecdysis of infective Haemonchus contortus larvae. Mol. Biochem. Parasitol., 33: 49-58.
PubMedDirect Link - Gardner, T., 2001. Declining amphibian populations: A global phenomenon in conservation biology. Anim. Biodivers. Conserv., 24: 25-44.
Direct Link - Hadju, V., K. Abadi, L.S. Stephenson, N.N. Noor, H.O. Mohammed and D.D. Bowman, 1995. Intestinal helminthiasis, nutritional status and their relationship: A cross-sectional study in urban slum school children in Indonesia. Southeast Asian J. Trop. Med. Public Health, 26: 719-729.
PubMedDirect Link - Haroun, E.M., O.M. Mahmoud, M. Magzoub, Y.A. Hamid and O.H. Omer, 1996. The haematological and biochemical effects of the gastrointestinal nematodes prevalent in camels (Camelus dromedarius) in central Saudi Arabia. Vet. Res. Commun., 20: 255-264.
CrossRefDirect Link - Holland, C.V., D.W. Crompton, S.O. Asaolu, W.B. Crichton, S.E. Torimiro and D.E. Walters, 1992. A possible genetic factor influencing protection from infection with Ascaris lumbricoides in Nigerian children. J. Parasitol., 78: 915-916.
PubMedDirect Link - Houlahan, J.E., C.S. Findlay, B.R. Schmidt, A.H. Meyers and S.L. Kuzmin, 2000. Quantitative evidence for global amphibian population declines. Nature, 404: 752-755.
CrossRefDirect Link - Huber, S.K., 2008. Effects of the introduced parasite Philornis downsi on nestling growth and mortality in the medium ground finch (Geospiza fortis). Biol. Conserv., 141: 601-609.
CrossRef - Hudson, P.J., A.P. Dobson and D. Newborn, 1998. Prevention of population cycles by parasite removal. Science, 282: 2256-2258.
Direct Link - Intapan, P.M., N. Morakote, K. Chansung and W. Maleewong, 2008. Hypereosinophilia and abdominopulmonary gnathostomiasis. Southeast Asian J. Trop. Med. Public Health, 39: 804-807.
PubMedDirect Link - Kiesecker, J.M. and. D.K. Skelly, 2001. Effects of disease and pond drying on gray tree frog growth, development and survival. Ecology, 82: 1956-1963.
Direct Link - Knox, D.P. and M.W. Kennedy, 1988. Proteinases released by the parasitic larval stages of Ascaris suum and their inhibition by antibody. Mol. Biochem. Parasitol., 28: 207-216.
PubMedDirect Link - Kozat, S., N. Yuksek, Y. Goz and I. Keles, 2006. Serum iron, total iron-binding capacity, unbound iron-binding capacity, transferrin saturation, serum copper and hematological parameters in pregnant akkaraman ewes infected with gastro-intestinal parasites. Turk. J. Vet. Anim. Sci., 30: 601-604.
Direct Link - Krist, A.C., 2000. Effect of the digenean parasite Proterometra macrostoma on host morphology in the freshwater snail Elimia livescens. J. Parasitol., 86: 262-267.
PubMedDirect Link - Lips, K.R., 1998. Decline of a tropical montane amphibian fauna. Conserv. Biol., 12: 106-117.
Direct Link - Lips, K.R., F. Brem, R. Brenes, J.D. Reeve and R.A. Alford et al., 2006. Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. Proc. Natl Acad. Sci. USA., 103: 3165-3170.
Direct Link - Maizels, R.M., D.A. Bundy, M.E. Selkirk, D.F. Smith and R.M. Anderson, 1993. Immunological modulation and evasion by helminth parasites in human populations. Nature, 365: 797-805.
CrossRefDirect Link - Materna, E.J., C.F. Rabeni and T.W. LaPoint, 1995. Effects of the synthetic pyrethroid insecticide on larval Leopard frogs. Environ. Toxicol. Chem., 14: 613-622.
Direct Link - Minchella, D.J. and M.E. Scott, 1991. Parasitism: A cryptic determinant of animal community structure. Trends Ecol. Evol., 6: 250-254.
CrossRef - Moreels, T.G., R.J. Nieuwendijk, J.G. de Man, B.Y. de Winter and A.G. Herman et al., 2004. Concurrent infection with Schistosoma mansoni attenuates inflammation induced changes in colonic morphology, cytokine levels and smooth muscle contractility of trinitrobenzene sulphonic acid induced colitis in rats. Gut, 53: 99-107.
CrossRefDirect Link - Mukerjee, S., R. Chander, B.L. Tekwani, S. Gupta, J.C. Katiyar, O.P. Shukla and N.K. Kapoor, 1990. Molecular basis of hyperlipidemia in golden hamsters during experimental infection with Ancylostoma ceylanicum (Nematoda: Strongylidae). Int. J. Parasitol., 20: 217-223.
PubMedDirect Link - Nokes, C., S.M. Grantham-McGregor, A.W. Sawyer, E.S. Cooper and D.A. Bundy, 1992. Parasitic helminth infection and cognitive function in school children. Proc. Biol. Sci., 247: 77-81.
Direct Link - Ouellet, M., J. Bonin, J. Rodrigue, J.L. DesGranges and S. Lair, 1997. Hindlimb deformities in free-living anurans from agricultural habitats. J. Wildlife Dis., 33: 95-104.
Direct Link - Peng, W., X. Zhou, X. Cui, D.W. Crompton and R.R. Whitehead et al., 1998. Transmission and natural regulation of infection with Ascaris lumbricoides in a rural community in China. J. Parasitol., 84: 252-258.
Direct Link - Ponton, F., D. Duneau, M.I. Sanchez, A. Courtiol and A.T. Terekhin et al., 2009. Effect of parasite-induced behavioral alterations on juvenile development. Behav. Ecol., 20: 1020-1025.
Direct Link - Poulin, R., 1994. Meta-analysis of parasite-induced behavioural changes. Anim. Behav., 48: 137-146.
CrossRef - Pounds, J.A., M.R. Bustamante, L.A. Coloma, J.A. Consuegra and M.P.L. Fogden et al., 2006. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature, 439: 161-167.
CrossRefDirect Link - Rehman, A. and D.P. Jasmer, 1998. A tissue specific approach for analysis of membrane and secreted protein antigens from Haemonchus contortus gut and its application to diverse nematode species. Mol. Biochem. Parasitol., 97: 55-68.
PubMedDirect Link - Rollins-Smith, L.A., J.K. Doersam, J.E. Longcore, S.K. Taylor, J.C. Shamblin, C. Carey and M.A. Zasloff, 2002. Antimicrobial peptide defenses against pathogens associated with global amphibian declines. Dev. Comp. Immunol., 26: 63-72.
PubMedDirect Link - Rollins-Smith, L.A., C. Carey, J. Longcore, J.K. Doersam, A. Boutte, J.E. Bruzgal and J.M. Conlon, 2002. Activity of antimicrobial skin peptides from ranid frogs against Batrachochytrium dendrobatidis, the chytrid fungus associated with global amphibian declines. Dev. Comp. Immunol., 26: 471-479.
PubMedDirect Link - Savioli, L., M. Neira, M. Albonico, M.J. Beach and H.M. Chwaya et al., 2000. Treatment for intestinal helminth infection. Review needed to take account of all relevant evidence, not only effects on growth and cognitive performance. BMJ, 321: 1226-1227.
PubMedDirect Link - Seppala, O., K. Liljeroos, A. Karvonen and J. Jokela, 2008. Host condition as a constraint for parasite reproduction. Oikos, 117: 749-753.
CrossRefDirect Link - Seppala, O., A. Karvonen and E. Valtonen, 2004. Parasite-induced change in host behaviour and susceptibility to predation in an eye fluke-fish interaction. Anim. Behav., 68: 257-263.
Direct Link - Sousa, W.P. and M. Gleason, 1989. Does parasitic infection compromise host survival under extreme environmental conditions?. The case for Cerithidea californica (Gastropoda: Prosobranchia). Oecologia, 80: 456-464.
Direct Link - Sparling, D.W., G.M. Fellers and L.L. McConnell, 2001. Pesticides and amphibian population declines in California, USA. Environ. Toxicol. Chem., 20: 1591-1595.
PubMedDirect Link - Storfer, A., 2003. Amphibian declines: Future directions. Diversity Distributions, 9: 151-163.
Direct Link - Stuart, S.N., J.S. Chanson, N.A. Cox, B.E. Young, A.S.L. Rodrigues, D.L. Fischman and R.W. Waller, 2004. Status and trends of amphibian declines and extinctions worldwide. Science, 306: 1783-1786.
Direct Link - Sukhdeo, M.V.K. and D.F. Mettrick, 1984. Effect of size of Trichinella spiralis (Nematode) infections on glucose and ion transport in the rat intestine. J. Parasit., 70: 499-506.
Direct Link - Tompkins, D.M. and M. Begon, 1999. Parasites can regulate wildlife populations. Parasitol. Today, 15: 311-313.
PubMedDirect Link - Tort, J., P.J. Brindley, D. Knox, K.H. Wolfe and J.P. Dalton, 1999. Proteinases and associated genes of parasitic helminths. Adv. Parasitol., 43: 161-266.
PubMedDirect Link - Vertucci, F.A. and P.S. Corn, 1996. Evaluation of episodic acidification and amphibian declines in the Rocky Mountains. Ecol. Appl., 61: 449-457.
Direct Link - Raji, Y., O.S. Akinsomisoye and O.M. Azeez, 2006. Impact of malaria parasites on reproductive indices of male mice. Reprod. Med. Biol., 5: 201-209.
CrossRef
DR PRAHLAD DUBE Reply
this is an intresting and use ful article that gives reliable information on the theme.
full article was not sen if possible send a soft copy of this and rlated articles.
Prof. William Reply
The main purpose of this scientific paper is to report new subject and results about the impact of studied parasite,Waltonella duboisi, on marsh frog. The hematological, biochemical and histopathological are very important indicators which reflect the effect of this parsite on marsh frog. The conclusion is also very intresting.