
M. A. Ghobashy
Department of Zoology, Faculty of Science, Suez Canal University, Ismailia, Egypt
M. F.M. Soliman
Department of Zoology, Faculty of Science, Suez Canal University, Ismailia, Egypt
E. A. Hassan
Department of Zoology, Faculty of Science, Suez Canal University, Ismailia, Egypt
International Journal of Zoological Research
Year: 2010 | Volume: 6 | Issue: 1 | Page No.: 13-23







ABSTRACT
In this study, the occurrence of heterophyid infection in two well-known hosts of heterophyd in Egyptian lake (Manzala); the mullet, Liza auratus and the cichlid, Oreochromis niloticus was investigated. Furthermore, the potential factors that possibly affect the occurrence of the infection including host sex, length, weight and seasonal variation were considered. The pathological response of the two fish host to the infection was studied. Results showed that the prevalence, abundance and intensity of infection in the two fish host greatly affected by the factor considered in contradictory way. The responses to infection and the possible effect of the interaction between all the considered factors are discussed in details. In addition, metacercarial infection caused alterations in the histological architecture of the infected tissues and in the composition of the muscle proteins as well which was more pronounced in O. niloticus and L. auratus, respectively. In conclusion, many biological and environmental factors do affect the occurrence of heterophyid infection in addition to the anthropogenic activity. L. auratus was more susceptible to the infection as compared to O. niloticus from the same habitat.
PDF Abstract XML References Citation
How to cite this article
M. A. Ghobashy, M. F.M. Soliman and E. A. Hassan, 2010. Responses of the Mullet, Liza auratus and the Cichlid, Oreochromis
niloticus from Lake Manzala (Egypt) to Heterophyd Infection. International Journal of Zoological Research, 6: 13-23.
DOI: 10.3923/ijzr.2010.13.23
URL: https://scialert.net/abstract/?doi=ijzr.2010.13.23
DOI: 10.3923/ijzr.2010.13.23
URL: https://scialert.net/abstract/?doi=ijzr.2010.13.23
INTRODUCTION
Fish may harbor many pathogens especially parasites which could interfere with the aquaculture industry and constitute a zoonotic threat, especially in developing countries (Elsheikha and Elshazly, 2008). Digenean parasites (Family: Heterophyidae), are among the most abundant pathogenic parasites affecting fish-eating birds and mammals worldwide (Sohn et al., 2005). There are indications that the infection has increased in the last few years because of an increasing production of fish and shellfish in unhygienic fish ponds, anthropogenic activity and the popularity of insufficiently processed food in many parts of the world (Keiser and Utzinger, 2004; Guk et al., 2007).
In Egypt, heterophyids, agents of human heterophyosis, are transmitted in fresh, saline inland and marine waters, where the molluscs and fish intermediate hosts are abundant (Elsheikha and Elshazly, 2008). It was claimed that heterophyosis infection rate was high in some areas such as in Port Said governorate, where 88% of school children were infected (Watson, 1960). More recently the prevalence of heterophyid metcercariae had been estimated allover the Egyptian lakes (Gharieb and Hamdy, 1969; Rifaat et al., 1980; Taraschewski, 1985; Mansour et al., 1987; Raef, 1994).
Host species, sex and condition and the environmental factors were reported by many researchers to influence the intensity and/or prevalence of fish parasites generally. Unfortunately, factors affecting the occurrence of heterophyid infection, the seasonal variation and the possible interaction between these factors are poorly studied (Sithithaworn et al., 1997; Ghobashy et al., 2006; Elsheikha and Elshazly, 2008). In addition, the simultaneous investigations conducted on the infection status of heterophyid metacercariae in a range of fish species, in a particular area are rare.
Therefore, in this study, occurrence of heterophyid infection in well-known hosts of heterophyids in Egyptian lakes; the mullet, Liza auratus and the cichlid, Oreochromis niloticus and the pathological response of the two fish host were investigated. Furthermore, the potential factors that possibly affect the occurrence of the infection including host condition and some environmental factors were considered.
MATERIALS AND METHODS
Area of the Study
This study was carried out to estimate the infection by heterophyid metacercariae in two fish species of Lake Manzala, Egypt. Lake Manzala is the largest of Nile Delta lakes; it has an area of 1275 km2. It is surrounded by the Mediterranean sea to the North, the Suez canal to the East and Damietta Branch of the Nile to the West (Northern latitude 30.6°’, Western longitude 32 25°’).
Sampling of the Specimens
Altogether, 229 specimens of the mullet; Liza auratus and 209 specimens of the cichlid, Oreochromis niloticus were collected monthly from Lake Manzala, during a period extended from June 2003 to May 2004 with a help of a fisherman. The collected fish were transported in ice boxes to the laboratory to be examined. Sex was determined according to Guerrero and Shelton (1974). The sizes of L. auratus were determined by measuring the fish length from the tip of the longest jaw to the center of the fork in the caudal fin and were classified into five length classes. The total length of O. niloticus was determined by measuring the distance from the tip of the longest jaw or the end of the snout to the longest caudal lobe pushed together and classified into six length classes (Miller and Lea, 1972). Weights of the collected fish were determined and were classified into five classes for L. auratus and six classes for O. niloticus. Sample sizes of collected fish, L. auratus and O. niloticus according to the factors considered including months, host sex, sizes and weights are shown in Table 1 and 2, respectively.
| Table 1: | Sample size of L .auratus according to the factors considered |
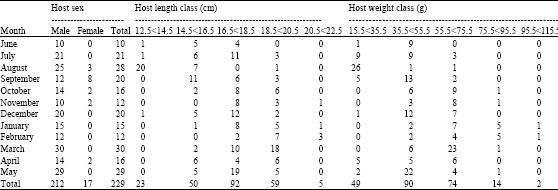 | |
| Table 2: | Sample size of O. niloticus according to the factors considered |
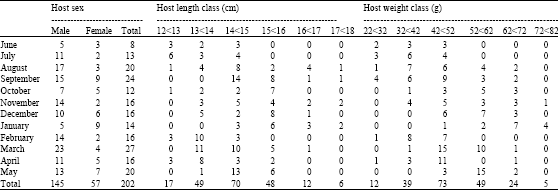 | |
Parasitological Study
The external surface of the host fish, L. auratus and O. niloticus was rinsed in water and the collected water was then examined for metacercarial infection. Body cavity was opened and all the internal viscera including the muscles, liver, intestine, kidney and gonads were examined for the presence of heterophyid metacercariae. These organs were kept in saline solution for few minutes to recover of parasites, then the organs were cut into small pieces, compressed between glass plates and examined under a binocular dissecting microscope (Soliman et al., 2004; Ghobashy et al., 2006). Metacercariae were counted cm-2 of the compressed tissue using binocular dissecting microscope and the intensity was expressed by the mean of 5 readings. The mean prevalence, intensity and abundance of metacercariae infection were estimated according to Margolis et al. (1982).
Pathological Studies
For histopathological examination, the infected muscles were fixed in 10% buffered formalin, embedded in paraffin, cut at 5 μm sections and stained with haematoxylin and eosin. For protein electrophoresis analysis, 0.5 g of the raw fish muscle were homogenized with 4 mL of 2% SDS extraction solution (2% sodium dodecyle sulphate, 5% mercaptoethanol and 60 mM Tris-HCl pH 8.0) (Etienne et al., 1999). The homogenates were centrifuged (20,000 change to g, for 15 min at 20°C) and the supernatants were collected. Solutions of samples and the molecular weight marker proteins were applied to the gel according to the protocol provided by Mackie et al. (2000).
Statistical Analysis
To satisfy the assumption of statistical analysis used, all the data were normalized by log (x+1) transformation to achieve Homoscedasticity or linearity. Factors affecting the metacercarial burdens were analyzed by GLM (General linear models) using a model with the negative binomial distribution error. For studying the differences between groups, analysis of variance was used. In all cases, significant difference was set at p = 0.05. Spearman correlation test was used for study association. The data were implemented by the software pack weights using SPSS 13.
RESULTS AND DISCUSSION
Out of 229 L. auratus examined during a one-year study, 37.9% were found infected with Heterophyidae metacercariae in their muscles. In general, the infection rate recorded in this study was lower than those reported by Wells and Blagg (1956) (100%), Fahmy and Selim (1959) (80%), Rifaat et al. (1980) (100%) and Mahmoud (1989) (87%) in different fresh water bodies. Dealing with Lake Manzala, Raef (1994) reported that 22.76% of the total fish examined were infected with metacercariae in Port-Said city, which was lower than that recorded in the present study. Generally, the low prevalence of infection in L. auratus recorded in this lake compared to most of the pervious studies on Mugil sp., may be attributed to an improvement in lake conditions and reduction in parasitism there. Other environmental and human factors may contribute to the healthy conditions of the lake.
On the other hand, out of 202 examined O. niloticus fish, 7.9% only were found infected with metacercariae in the muscles with a significantly lower total abundance (0.01±0.005 cm-2) and intensity (0.2±0.06) as compared to L. auratus (0.6±0.2 and 1.5±0.5 cm-2, respectively) (p<0.001). The prevalence rate (7.9%) was also very low compared to those recorded in L. auratus (37.9%) (p<0.001) indicating that O. niloticus seems to be more resistant to infection. High infection rates were recorded by Fahmy and Selim (1959) (62%) in Tilapia sp., Gharieb and Hamdy (1969) (67%) in T. zilli and T. nilotica from Lakes El-Manzala and Borollos. This may suggest that the conditions in Lake Manzala have apparently improved.
The low percentage of infection recorded in both L. auratus and O. niloticus in this study compared to the majority of previous studies may be partly due to variation in the nutrient content of the water. However, the epidemiological picture of food borne trematodiasis has changed in the recent years. In some settings, the prevalence of food borne trematode infections increases while in other it decreases significantly, which can be explained by factors such as social and economic development, urbanization, adequate food inspections, health education campaigns, use of chemical fertilizers and water pollution (Keiser and Utzinger, 2004).
Regarding the seasonal and the monthly variation of infection (Table 3), Lee (1968) stated that parasitic infection in freshwater fish might correlate to the season. Chubb (1979) found that freshwater digeneans often show seasonal changes in prevalence and abundance related to cercarial emergence with an increase in water temperature. Sithithaworn et al. (1997) observed that seasonal variation in metacercariae is a common phenomenon in areas with both high and low endemicity of infection. However, in this study, the effect of the seasonal variation on metacercarial infestation was obvious in L. auratus, where highest infection rate (52.5%) was recorded in summer while the lowest rate was recorded in spring (24%). This result agrees with those of Kang et al. (1985) and El-Naffar and El-Shahawi (1986) who reported that heterophyid infection is more abundant in summer. Raef (1994) recorded the highest infestation rate in summer (31.40%), followed by spring (24.66%), then autumn (16.43%) and lowest in winter (12.51%). The high prevalence of infection in summer may coincide with time of emergence of cercaria from snail vectors, which is usually encouraged by the temperature increase of water (Oshima and Nishi, 1963). Moreover, in summer, the water is more liable to contamination with excreta of human beings, as of the reservoir hosts. This coincides with the time of maturation of the snail host, which flourishes better in warm water when the surrounding environmental conditions become favorable (Samman, 1974). The highest abundance and intensity of infection in L. auratus was recorded in spring, while the lowest rates were recorded in autumn (Table 3). Abou-Zakham et al. (1990) found that the highest intensity of the infection with Stictodora tridactyla metacercariae in Mugil sp. and Tilapia sp., collected from Lake Manzala is recorded in summer.
Regarding O. niloticus, the highest infection rate is recorded in spring with the highest intensity while the lowest is in winter (Table 3). Makhalouf et al. (1987) and Mansour et al. (1987) found that in Tilapia sp., infected with H. heterophyes metacercariae in three brackish water lakes in Egypt (Manzalah, Borollos and Idku Lakes), the highest prevalence of infection was during summer months and the highest abundance and intensity of infection in spring and the lowest in winter in agreement to that recorded in this study.
| Table 3: | Factor affecting prevalence, intensity and abundance of Heterophyid metacercarial infection in L. auratus and O. nilotica |
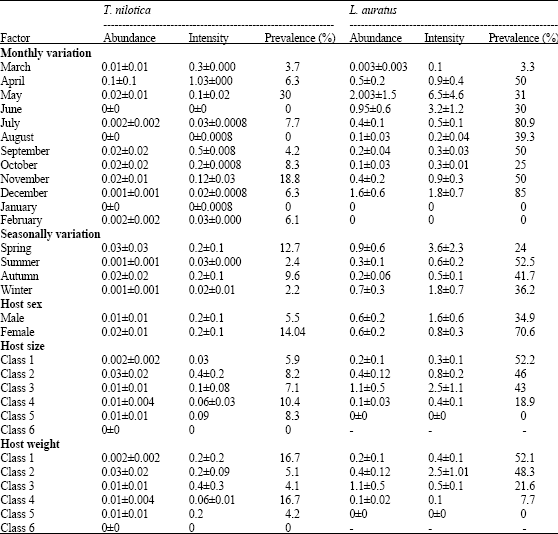 | |
The highest infection rate in the muscles of L. auratus is recorded in December (85%) followed by July (80.9%), while no infection is found in January and February (Table 3). Since, these winter months have nearly similar temperatures, this may indicate the weak effect of temperature on the infection rate. In contrary to L. auratus, highest infection rates in O. niloticus (Table 3) is recorded in May (30%) followed by November (18.8%), while no infection is recorded in June, August and January confirming that temperature has a minor influence on the parasite invasion to the examined fish. Smyth and Halton (1983) recorded that the life span of infective free-living stages of a number of trematode species is negatively correlated with temperature.
Regarding the host sex, the infection rate with heterophyid metacercariae in L. auratus and O. niloticus was higher in female fish (70.6%), (14.04%) compared to male one (34.9%), (5.5%), respectively, indicating that the female host is more susceptible to the infection. In contrast, the present results showed that intensity of the infection was higher in the male hosts compared to the female in L. auratus (p<0.01) and nearly equal in O. niloticus (Table 3). Generally, reports about the susceptible host sex to infection are varied and conflicting. Harrod and Griffiths (2005) reported that the female hosts are more heavily infested than male ones. In contrast, Paling (1965) found that male hosts are heavily parasitized than female hosts. However, the sex differences for the preferences of the infection may be attributed to the immune response of the host or to the difference in endocrine glands activities between the male and female host fishes which have been suggested by many authors (Donaldson and Fagerlund, 1970). Males are usually more sensitive to the parasites than females apparently due to testosterone synthesis, which might act in decreasing immune competency (Gbankoto et al., 2001). This might explain the higher intensity of infection recorded in the males of L. auratus. However, Poulin (1996) concluded that the sex difference is irrelevant and no significant differences between the prevalence and intensity of infection in female and male hosts.
The prevalence of Heterophyidae metacercarial infection rate was clearly inversely proportional to the fish length of L. auratus where it decreased with increasing of the fish length (Table 3) no infection was recorded in length range 20.5-22.5 cm (class 5). Chappell (1969) also revealed that the infestation level of parasites generally attained a negative relationship with the length of host fish. This in turn may be attributed to the more developed immunity of the fish with greater size and age. Furthermore, Malek and Mobedi (2001) reported that the decrease in the prevalence of infection in the larger fish could be as a result of increase in the mortality of infected fish, increase in the resistance of the host with increase in length and a change of feeding habit of the host. In contrast, the present results showed that in O. niloticus, the highest infection rate was on fish with length Class 4 (15-16 cm) (Table 3). Wang et al. (2002) recorded that there is no positive correlation between infection rates of Haplorchis metacercariae in fish and sizes of infected fish.
In L. auratus, abundance and intensity of the infection (Table 3) was significantly the highest (1.1±0.5 and 2.5±1.1 cm-2, respectively) in length range 16.5-18.5 cm (class 3) as compared to the other classes (p<0.05) were recorded in L. auratus with the length (14-15 cm). In O. niloticus abundance and intensity of the infection (Table 3) was significantly the highest in the length class 2 (13-14 cm). Khidr (1990) found that the prevalence and intensity are higher in larger Tilapia fishes than in the smaller ones while Müller (1995) found that numbers of parasites are significantly correlated with fish length. However, in the present study, both fish showed no regular pattern for abundance and intensity of infection.
In this study, the prevalence of the metacercarial infection was inversely proportional with weight of L. auratus as it decreased with increasing the fish weight. While, in O. niloticus, it was the highest in fish with weight (22-32 g) and (52-62 g) with the same value (16.7%) and no infection was recorded in fish with weight (72-82 g). Abundance and intensity of infection was the highest in fish with weight (35.5-55.5 g) and the lowest in fish with weight (75.5-95.5 g) in L. auratus (Table 3). While, in O. niloticus, abundance is the highest in fish with weight (22-32 g) and the lowest in fish with weight (32-42 g) and (62-72 g). Intensity is the highest in fish with weight (42-52 g) and the lowest in fish with weight (62-72 g). Pampoulie et al. (2000) suggested that the parasite burden may be synchronized with the host condition inducing a modification of prevalence in relation with the life cycle of host. All these findings indicate that the fish host may play a role regarding the relation between the host lengths or weights and the incidence of the infection. No significant correlation regarding the intensity of infection with the host lengths or weights was recorded in both fish species.
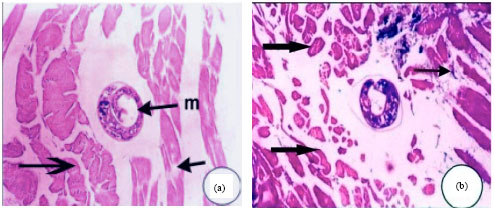 | |
| Fig. 1: | (a) Histopathological alterations in muscles of L. auratus and (b) accompanied with heterophyid metacercarial infection showing necrotic degeneration, destruction, atrophy and longitudinal splitting of muscles fibers (X 200 |
The GLM was used to test the interaction between the seasonal and monthly variations of infection, host sex and weight and length categories on prevalence, intensity and abundance of the infection. Results showed that only the prevalence of infection in L. auratus was significantly dependent on the monthly variations (p<0.001) and marginally dependent on the host sex (p = 0.059). On the other hand, abundance and intensity of the infection was independent of the tested factors. In contrast, in O. niloticus, the infection was independent on the entire tested factors confirming the opposite responses of both fish to the stressor.
Regarding the pathological response of the fish hosts to infection, results showed that heterophyid metacercariae in the muscles of L. auratus and O. niloticus led to formation of fibrous capsules surrounding the metacercariae, necrotic degeneration, destruction, atrophy and longitudinal splitting. The damages caused to the muscles of O. niloticus were more severe than those found in L. auratus (Fig. 1a, b) indicating that both host showing different resistance to pathogens. The histopathological alterations found in both fish in this study can be compared with those, Mahmoud et al. (1989), Taraschewski (2000), Malek and Mobedi (2001), Villeneuve et al. (2005) and Umadevi and Madhavi (2006). In contrast, Raef (1994) stated that, the metacercariae of heterophyid trematodes have little pathological changes in fish muscles and only apparent tissue response is the formation of dense fibrous layer around the encysted parasite.
Using SDS-PAGE (Polyacryamide Gel Electrophoresis), metacercarial infection caused dramatic changes in the muscle proteins pattern in both fish species in three main ways; disappearance of some protein bands, fading away of some existing bands or appearance of new bands. In addition, infection also affects the relative quantities of protein fraction either by being increased or decreased. The alteration was more severe in L. auratus as compared to O. niloticus (Table 4). Damage to muscle of fish and other edible aquatic organisms have been widely reported from various infective agents, including helminthes (Soliman et al., 2004; Ghobashy et al., 2006). However, this fluctuation in protein bands might be considered as a reflection of stressor impact on ribosome and RNA levels and consequently on protein synthesis (De Bruin, 1976). The changes in the protein pattern could adversely affect the taste, texture and marketability of the infected meat (Stentiford et al., 2003). The serious alterations induced in the protein composition of the infected fish would affect in all probability the nutritional value of this vital food source (Ghobashy et al., 2006).
| Table 4: | SDA-PAGE pattern of muscles proteins of the non-infected and infected L. auratus and O. niloticus with heterophyid metacercariae |
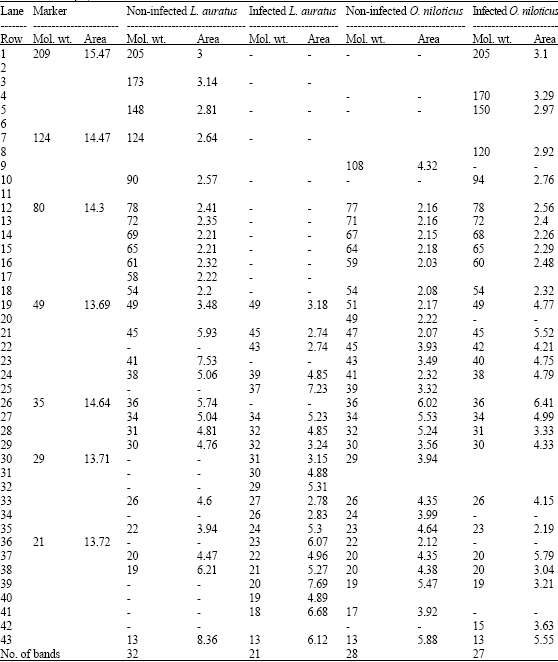 | |
| -: Indicators of disappearance of existing protein bands or appearance of new ones | |
In conclusion, many biological and environmental factors do affect the occurrence of heterophyid infection in addition to the anthropogenic activity and Liza auratus was more susceptible to the infection as compared to O. niloticus from the same habitat. Further studies are needed for more explanation and interpretation of our findings.
REFERENCES
- Abou-Zakham, A., A. El-Shazley, F. El-Ganayni, S.A. Romia, A.F. Abou-Shady and E.I. El-Kholy, 1990. Seasonal variation and incidence of Stictodora tridactyla in fish from Lake Manzala. J. Egypt. Soc. Parasitol., 20: 117-121.
Direct Link - Chappell, H.L., 1969. The parasites of the Three-spined stickleback Gasterosteus aculeatus L. from a Yorkshire Pond II. Variation of the parasite fauna with sex and size of fish. J. Fish Biol., 1: 339-347.
Direct Link - Elsheikha, H.M. and A.M. Elshazly, 2008. Host-dependent variations in the seasonal prevalence and intensity of heterophyid encysted metacercariae (Digenea: Heterophyidea) in brackish water fish in Egypt. Vet. Parasitol., 153: 65-72.
CrossRefDirect Link - Etienne, M., M. Jerome, J. Fleurence, H. Rehbein and R. Kundiger et al., 1999. A standardized method of identification of raw and heat-processed fish by urea isoelectronic focusing: A collaborative study. Electrophoresis, 20: 1923-1933.
Direct Link - Fahmy, M.A.M. and M.K. Selim, 1959. Studies on some trematode parasites of dogs in Egypt with special reference to the role played by fish in their transmission. Parasitol. Res., 19: 3-13.
Direct Link - Gbankoto, A., C. Pampoulie, C.A. Marques and G.N. Sakiti, 2001. Occurrence of Myxosporean parasites in the gills of two tilapia species from Lake Nokoue (Benin, West Africa): Effect of host size and sex and seasonal patterns of infection. Dis. Aquat. Organ., 44: 217-222.
Direct Link - Guerrero, R.D. and W.L. Shelton, 1974. An aceto-carmine squash technique for sexing juvenile fishes. Prog. Fish-Cultur., 36: 56-56.
CrossRefDirect Link - Guk, S.M., E.H. Shin, J.L. Kim, W.M. Sohn and K.S. Hong et al., 2007. A survey of Heterophyes nocens and Pygidiopsis summa metacercariae in mullets and gobies along the coastal areas of the Republic of Korea. Korean J. Parasitol., 45: 205-211.
Direct Link - Harrod, C. and D. Griffiths, 2005. Ichthyocotylurus erraticus (Digenea: Strigeidae): Factors affecting infection intensity and the effects of infection on pollan (Coregonus autumnalis), a glacial relict fish. Parasitology, 131: 511-519.
Direct Link - Kang, S.Y., S.I. Kim and S.Y. Cho, 1985. Seasonal variations of metacercarial density of Clonorchis sinensis in fish intermediate host, Pseudorasbora parva. Kisaengchunghak Chapchi., 23: 87-94.
Direct Link - Keiser, J. and J. Utzinger, 2004. Chemotherapy for major food-borne trematodes: A review. Exp. Opin. Pharmacother., 5: 1711-1726.
Direct Link - Lee, J.T., 1968. Studies on the metacercariae from freshwater fishes in the Kum-Ho River. Kisaengchunghak Capchi., 6: 77-99.
Direct Link - Mackie, I., A. Craig, M. Etienne, M. Jerome and J. Fleurenceet al., 2000. Species identification of smoked and gravad fish products by sodium dodecylsulphate polyacrylamide gel electrophoresis urea isoelectric focusing and native isoelectric focusing: A collaborative study. Food Chem., 71: 1-7.
CrossRef - Makhalouf, L.M., A.A. Zakham, K. EI, A.A.F. Shady, S. Abdel-Magied and A. El-Shazley, 1987. Heterophyids of some fresh water fish from Mansoura, Egypt. J. Egypt. Soc. Parasitol., 17: 573-576.
Direct Link - Malek, M. and I. Mobedi, 2001. Occurrence of clinostomum complanatum (Rudolphi, 1819) (Digenea: Clinostomatidae) in Capoeta capoeta gracilis (Osteichthys: Cyprinidae) from Shiroud River, Iran. Iranian J. Publ. Health, 30: 95-98.
Direct Link - Mansour, N.S., M.M. Youssef, H.N. Awadalla, N.A. Hammouda and L.M. Boulos, 1987. Heterophyid metacercariae in the fish Tilapia sp. (cichlidae) from Edku, Maryout and Manzala Lake in Egypt. J. Egypt. Soc. Parasitol., 17: 481-493.
Direct Link - Margolis, L., G.W. Esch, J.C. Holmes, A.M. Kuris and G.A. Shad, 1982. The use of ecological terms in parasitology (Report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol., 68: 131-133.
CrossRefDirect Link - Muller, G., 1995. Prevalence and abundance of two trematode parasites, Diplostomum phoxini and Macrolecithus papilliger in European minnows (Phoxinus phoxinus) in an artificial Swiss Alpine Lake. Aquat. Sc. Res. Across Boundaries, 57: 119-126.
Direct Link - Paling, I., 1965. The population dynamics of the monogenean gill parasite Discocotyle sagittata Leuckart on Windemere trout, Salmo trutta, L. Parasitology, 55: 667-694.
Direct Link - Poulin, R., 1996. Sexual inequalities in helminth infections: A cost of being male? Am. Nat., 147: 287-295.
Direct Link - Sithithaworn, P., V. Pipitgool, T. Srisawangwong, D.B. Elkins and M.R. Haswell-Elkins, 1997. Seasonal variation of Opisthorchis viverrini infection in cyprinoid fish in northeast Thailand: Implications for parasite control and food safety. Bull. World Health Organ., 75: 125-131.
Direct Link - Sohn, W., J. Kim and H. Song, 2005. Two species of goby, Boleophthalmus pectinirostris and Scartelaos sp., as the new second intermediate hosts of heterophyid fluke in Korea. Korean J. Parasitol., 43: 161-164.
Direct Link - Soliman, M.F.M., N.S. El-Shenawy and M.A. Ghobashy, 2004. Parasitological aspects and biochemical changes of infected cultured tilapia (Oreochromis hybrid). Acta Ichthyol. Piscat., 34: 21-32.
Direct Link - Stentiford, G.D., M. Longshaw, B.P. Lyons, G. Jones, M. Green and S.W. Feist, 2003. Histopathological biomarkers in estuarine fish species for the assessment of biological effects of contaminants. Mar. Environ. Res., 55: 137-159.
CrossRefPubMedDirect Link - Taraschewski, H., 1985. Investigations on prevalence of Heterophes species in 12 populations of the first intermediate host in Egypt and Sudan. J. Top. Med. Hyg., 88: 265-271.
Direct Link - Taraschewski, H., 2000. Host-parasite interactions in acanthocephala: A morphological approach. Adv. Parasitol., 46: 1-179.
CrossRef - Umadevi, K. and R. Madhavi, 2006. The life cycle of Haplorchis pumilio (Trematoda: Heterophyidae) from the Indian region. J. Helminthol., 80: 327-332.
CrossRefDirect Link - Villeneuve, D.L., L.R. Curtis, J.J. Jenkins, K.E. Warner and F. Tiltonet al., 2005. Environmental stresses and skeletal deformities in fish from the Willamette River, Oregon. Environ. Sci. Technol., 39: 3495-3506.
Direct Link - Wang, J.J., L.Y. Chung, J.D. Lee, E.E. Chang, E.R. Chen, D. Chao and C.M. Yen, 2002. Haplorchis infections in intermediate hosts from a clonorchiasis endemic area in Meinung, Taiwan, Republic of China. J. Helminthol., 76: 185-188.
Direct Link - Wells, W.H. and W. Blagg, 1956. A survey of human intestinal parasites in a fishing village of Northern Egypt. Am. J. Med. Hyg., 5: 266-268.
Direct Link