
G. Rosangkima
Cell and Tumor Biology Laboratory, Department of Zoology, North-Eastern Hill University, Shillong 793022, India
T. Rongpi
Cell and Tumor Biology Laboratory, Department of Zoology, North-Eastern Hill University, Shillong 793022, India
S.B. Prasad
Cell and Tumor Biology Laboratory, Department of Zoology, North-Eastern Hill University, Shillong 793022, India
International Journal of Zoological Research
Year: 2008 | Volume: 4 | Issue: 4 | Page No.: 203-213







ABSTRACT
In the present study, the antitumor potential of methanol extract of D. pentagyna and its effect on the level of sialic acid and agglutinability of normal and transformed cells with concanavalin A and wheat germ agglutinin were reported in order to find the possible role of sialic acid in the antitumor activity of D. pentagyna. Methanol extract of stem bark of D. pentagyna showed maximum survivability of Dalton`s lymphoma-bearing mice at a dose of 20 mg kg-1 (%ILS ~ 70%). The present finding shows a significant D. pentagyna extract-mediated decrease in sialic acid content of normal and transformed cells and increase in ascites supernatant. The plant extract treatment decreases DL cells agglutinability with concanavalin A and wheat germ agglutinin in vitro and in vivo. It also decreases normal lymphocytes agglutinability with wheat germ agglutinin while increased agglutinability was observed with concanavalin A. D. pentagyna-mediated release of sialic acid from the surface of DL cells and decrease in the degree of cell agglutination with conA and wheat germ agglutinin suggested the occurrence of topographical changes on the cell surface and rendering them more immunogenic or accessible to the cells of immune system in the hosts.
PDF Abstract XML References Citation
How to cite this article
G. Rosangkima, T. Rongpi and S.B. Prasad, 2008. Effect of Dillenia pentagyna Extract on Sialic Acid Content and Agglutinability of Normal and Tumor Cells with Concanavalin A and Wheat Germ Agglutinin. International Journal of Zoological Research, 4: 203-213.
DOI: 10.3923/ijzr.2008.203.213
URL: https://scialert.net/abstract/?doi=ijzr.2008.203.213
DOI: 10.3923/ijzr.2008.203.213
URL: https://scialert.net/abstract/?doi=ijzr.2008.203.213
INTRODUCTION
Dillenia pentagyna Roxb. (Dilleneaceae), distributed in Indo-Malaysian areas extending to tropical Australia; throughout India particularly in sub-tropical Himalayas, is a deciduous tree found in most places of Mizoram state, India. Present preliminary investigation through literature review and personal interview with local herbal practitioners revealed the use of this plant by the people of Mizoram for the treatment of gastric cancers, diarrhea and other stomach ailments. The antitumor potential of methanol extract of this plant against murine ascites Dalton`s lymphoma has also been reported by Rosangkima and Prasad (2004). However, there is no detailed scientific investigation about the possible mechanism(s) involved in its antitumor potential.
Sialic acids are a family of 9-carbon acidic sugars that can be O-acetylated at any one of four hydroxyl groups, located at position C-4, -7, -8 and -9. This modification, which is found in nearly all animals expressing sialic acids and certain bacteria, is known to be involved in regulating a variety of biological events. One of the more important processes that appear to be heavily influenced by O-acetylation is cancer development (Tiralongo and Schauer, 2004; Kelm and Schauer, 1997). In physiologic pH, the sialic acid is negatively charged and this affects the physiological and chemical properties of glycoproteins significantly (Filipovic and Buddecke, 1979; Orekhov et al., 1989). Lipid-bound sialic acid levels have been used as a tumor marker of various malignant neoplasms including bronchial, prostate, ovary, breast and colon cancer and malignant melanoma (Pulcinski et al., 1986; Stringou et al., 1992). It was believed that changes in bound carbohydrates at the cell surface might result in persistent cell division, decreased intercellular adhesiveness, altered transport, altered/masked immunogenicity and other specialized functions accompanying malignant transformation (Warren et al., 1978). The widely distributed sialic acid moieties of glycoprotein are reported to have protective and regulatory functions at the cell surface (Warren et al., 1978). Prasad (1986) reported that the lectin mediated agglutination behavior of normal and malignant cells depends upon the changes in cell surface sialic acid moieties. Since sialic acid occupies a terminal position in carbohydrate chains of mammalian glycoproteins, it might be expected that it would play an important role as a receptor at the cell surface. Yet on masking of cell surface antigens, it may be involved in nonspecific repulsion of cells or macromolecules by virtue of its negative charge (Nicol and Prasad, 2002). Thus, the cell surface components could play vital role in the malignant transformation of a variety of cells.
Therefore, it become an interesting investigation to determine the level of sialic acid in normal and transformed cells and their agglutinability with concanavalin A and wheat germ agglutinin under D. pentagyna extract treatment conditions in order to find the possible role of sialic acid in the antitumor activity of D. pentagyna.
MATERIALS AND METHODS
Plant Material
The stem bark of D. pentagyna was collected in September 2007 from Kawlkulh village, Mizoram state, India. The plant material was authenticated by Dr. P.B. Gurung, Herbarium specialist, Department of Botany, North Eastern Hill University, Shillong (India) and a voucher specimen (No. SBP 001) was deposited in the Department of Zoology, NEHU. The fresh plant material collected was used for this study.
Animals and Tumor Model
Inbred Swiss albino mice (male) in the age group of about 10-12 weeks old were used for the experiments. All mice are maintained in the laboratory under conventional conditions at room temperature of 20 ± 2 °C with free access to food pellets (Amrut Laboratory, New Delhi) and water ad libitum. Approximately 1x107 viable Ascites Dalton`s lymphoma cells is being intraperitoneally (i.p.,) transplanted per animal (0.25 mL in Phosphate-Buffered Saline (PBS), pH 7.4). Tumor transplanted hosts usually survived for 19-21 days.
Preparation of Methanol Extract
Methanol extract of D. pentagyna was prepared following the method described by Alasbahi et al. (1999). Briefly, stem bark of D. pentagyna was shade-dried and grounded. Two hundred and fifty gram of powdered material was extracted with 1 L of absolute methanol at room temperature for 24 h. The tissue-solvent mixture was filtered using Whitman No. 1 filter paper and the filtrate was evaporated to dryness in a rotary evaporator. The dried and powdered extract was collected and stored under 0 °C until used. Methanol extract was tried to dissolve preferentially in double distilled water, methanol, sodium hydroxide solution and phosphate-buffered saline (PBS, pH 7.4). 0.05% sodium hydroxide solution (minimum concentration showing maximum solubility of the extract) was used as the extract vehicle for the experiment.
Antitumor Activity Study
Antitumor activity of methanol extract of D. pentagyna was studied following the method described by Sakagami et al. (1987). Day of tumor transplantation was designated as day 0. D. pentagyna extract treatment (10 to 200 mg kg-1) was given for five consecutive days starting from day 1 of tumor transplantation. The antitumor efficacy was reported in percentage of average Increase in Life Span (ILS) calculated using the formula:
where, T and C are the mean survival days of treated and control groups of mice, respectively. Changes in the average body weight, ascites tumor volume and food consumption of animals in different groups were monitored for 18 days from the day of tumor transplantation. Different groups of mice with different doses consisted of 10 mice each. The most potent dose was selected for further biochemical and cell agglutination studies.
Sialic Acid Estimation
In sialic acid estimation, a single dose of D. pentagyna extract (20 mg kg-1), showing the most potent antitumor activity, was given intraperitoneally on the 10th day of tumor growth. Sialic acid estimation was done in normal lymphocytes, DL cells and ascites supernatant (SN) after 24, 48, 72 and 96 h of DPE treatment following the method of Warren (1959). Tumor-bearing control mice received 0.25 mL of extract vehicle alone. Changes in body weight of the hosts, ascites tumor volume and food consumption were also monitored for 18 days in tumor-bearing control and DPE treatment group of mice.
Light Microscopical Studies
Tumor-bearing mice were given a single dose of DPE (20 mg kg-1) on the 10th day of tumor growth and tumor-bearing control received 0.25 mL of extract vehicle alone. Animals in different treatment groups were sacrificed by cervical dislocation after 24, 48, 72 and 96 h of treatment and ascites tumor was collected and centrifuged at 1000 rpm for 5 min at 4 °C, washed in phosphate-buffered saline (PBS, 0.15 M NaCl, 0.01 M sodium phosphate buffer, pH 7.4) . The cell pellet was suspended again in PBS (1:4) and a drop of the cell suspension was taken on a clean slide and a thin smear was prepared. The smear was air-dried, fixed in absolute methanol for 15 min and stained the following day with Leishman`s stain. The cells in different slides were studied and photographed.
Cells and Cell Suspension
Normal lymphocytes were collected from the spleen of tumor-bearing mice by squeezing the spleen between two sterilized glass slides. The cells were washed with phosphate-buffered saline (PBS, 0.15 M NaCl, 0.01 M sodium phosphate buffer, pH 7.4) and erythrocytes were removed by an osmotic shock in distilled water. The cells were again washed with Phosphate-Buffered Saline (PBS) and a single cell suspension of splenocytes was prepared in Phosphate-Buffered Saline (PBS). Dalton`s lymphoma cells collected from the peritoneal cavity of mice was lysed with 0.85% NH4 Cl to remove erythrocytes, washed twice with PBS and a single cell suspension was prepared in PBS.
Agglutination Assay
The cell suspensions of both normal and transformed cells (3°106 cells mL-1 PBS) were incubated with 10, 25, 50, 100 and 200 μg DPE mL-1 for 10, 30 and 60 min at 37 °C in sterile tissue culture tubes. The controls were run parallel without DPE. The cell suspensions were shaken intermittently. After appropriate time of incubation, 0.5 mL of cell suspension from different batches was mixed with concanavalin A (250 μg mL-1) and wheat germ agglutinin (100 μg mL-1) in a shallow concavity slides and incubated for 15 min at room temperature. The cell agglutination was examined under the ordinary light microscope at low magnification. The degree of agglutination was denoted by using +ve and -ve signs. Agglutination of 5-6 cells at minimum of 10 places under the microscope was marked as single + . The points in graph and degree of cell agglutination indicated as + signs are mean values of 6 sets of experiments.
To study the effect of DPE on the agglutinability of normal and transformed cells in vivo, tumor-bearing mice were given a single intraperitoneal injection of DPE at a dose of 20 mg kg-1. After 1, 2 and 4 h of DPE treatment, the animals were sacrificed and splenocytes and DL cells were collected as described above. The cell suspension (3x106 cells mL-1 PBS) of both normal and transformed cells were prepared and processed for agglutination studies with concanavalin A (250 μg mL-1) and wheat germ agglutinin (100 μg mL-1) as described above.
Cell viability was also checked with Trypan blue exclusion test at different levels of experiments. Cell viability was normally 85-90%.
Statistical Analysis
Data were shown as the Mean ± SD. Analysis of statistical significance was done using Student`s t-test. Number of replicates (N) = 6. A value of p = 0.05 was considered to be significant for comparison between data sets.
RESULTS
Antitumor Activity
Among different doses of the plant extract studied, 20 mg kg-1 exhibited maximum antitumor potential (%ILS~70) (Fig. 1). The pattern of changes in body weight, ascites tumor volume and food consumption were as shown in the Fig. 2a-c. As compared to the tumor-bearing control, a significant DPE-mediated decrease in body weight and ascites tumor volume and increased food consumption were noted in DPE treated mice after 14th day of tumor growth.
Sialic Acid Content
In tumor-bearing control during 11-14 days of tumor growth, there was a slight increase of sialic acid concentration in normal lymphocytes, DL cells and ascites supernatant. DPE single treatment (20 mg kg-1) causes a significant decrease of sialic acid content in normal lymphocytes (Fig. 3a) and DL cells (Fig. 3b) during 72-96 h of treatment, while a significant increase was observed in ascites supernatant during 72 to 96 h of treatment (Fig. 3b).
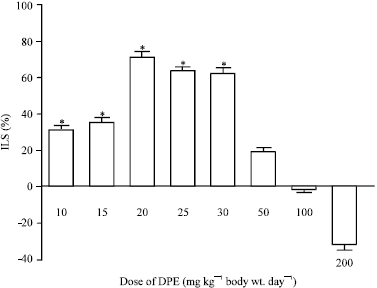 | |
| Fig. 1: | Percentage increase in life span (%ILS) of tumor-bearing mice treated with different doses of DPE. Results are Mean ± SD *Doses of DPE showing %ILS = 20% were considered to possess antitumor potential |
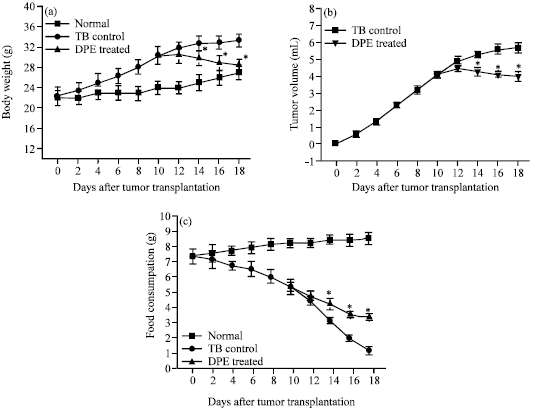 | |
| Fig. 2: | The pattern of changes in (a) the average body weight, (b) ascites tumor volume and (c) food consumption of normal, tumor-bearing control and DPE treated mice (20 mg kg-1 , on the 10th day of tumor growth) |
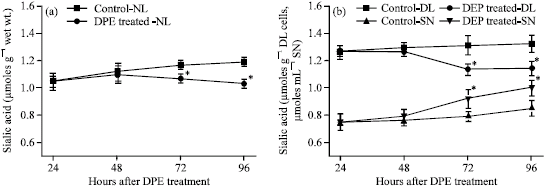 | |
| Fig. 3: | The pattern of changes in sialic acid content of normal lymphocytes (a), DL cells and ascites supernatant (b) under different DPE treatment conditions in vivo. Results are Mean ± SD. Student`s t-test; as compared to the corresponding control, n = 6, *p ≤ 0.05. Sialic acid content of ascites supernatant was expressed as μmoles mL-1 SN |
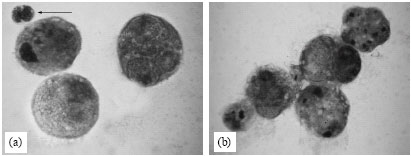 | |
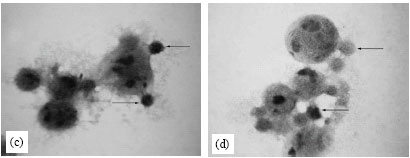
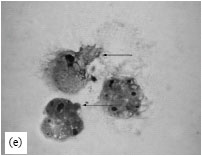
| Fig. 4: | Light micrographs of tumor cells under different treatment conditions in vivo. (a) Control tumor cells, (b) Twenty four hours of DPE treatment showing infiltration of leukocytes towards the tumor cells, (c and d) DPE treatment for 72 and 96 h showing more infiltration of leukocytes and (e) Tumor cells showing gradual disintegration of plasma membrane with membrane vacuoles |
Light Microscopy
Control tumor cells showed rounded shape and surrounded by a very few leukocytes (Fig. 4a). DPE treatment caused more leukocyte infiltration towards tumor cells (Fig. 4b, c). DPE treatment for 96 h also resulted in the appearance of membrane vacuoles and gradual disintegration of plasma membrane leading to lysis of tumor cells (Fig. 4d, e).
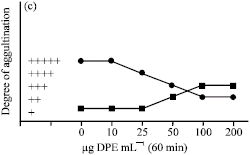
Agglutination of Normal Lymphocytes
Lymphocytes obtained from the spleen of mice showed a little degree of cell agglutination with concanavalin A while a higher degree of cell agglutination was noted with wheat germ agglutinin (Fig. 5, 6). When lymphocytes were treated with DPE for different time interval of time, a dose-dependent increase in the degree of lymphocytes agglutination with concanavalin A was observed. On the other hand, DPE treatment causes a dose-dependent decrease in the degree of lymphocytes agglutination with wheat germ agglutinin (Fig. 5, 6). Maximum increase in the degree of lymphocytes agglutination with concanavalin A after DPE treatment was noted after 60 min at a dose of 100 and 200 μg mL-1 (Fig. 5c), while maximum decrease in the degree of lymphocytes agglutination with wheat germ agglutinin was observed during 10 to 30 min of DPE treatment at a dose of 100 and 200 μg mL-1 (Fig. 6a, b).
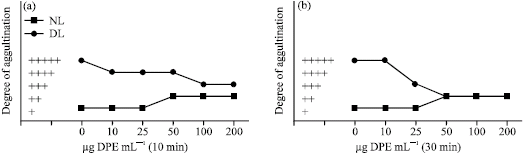 | |
| Fig. 5: | Agglutination pattern of normal and transformed (DL) cells with concanavalin A (250 μg mL-1) after incubation with and without (control) DPE in vitro at 37 °C for different intervals of time |
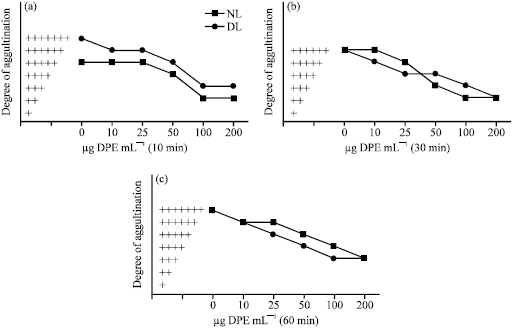 | |
| Fig. 6: | Agglutination pattern of normal and transformed (DL) cells with wheat germ agglutinin (100 μg mL-1) after incubation with and without (control) DPE in vitro at 37 °C for different intervals of time |
Agglutination of Dalton`s Lymphoma Cells
Dalton`s lymphoma cells showed a very high degree of agglutination with concanavalin A and wheat germ agglutinin during 10, 30 and 60 min of incubation. As compared to the controls, treatment with DPE dose-dependently decreased the degree of agglutination of Dalton`s lymphoma cells with concanavalin A and wheat germ agglutinin during 10, 30 and 60 min of incubation (Fig. 5, 6).
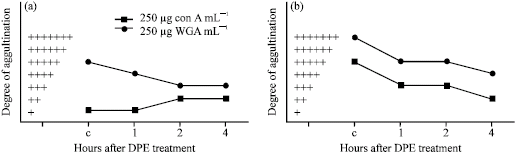 | |
| Fig. 7: | Agglutination pattern of normal lymphocytes (a) and DL cells (b) with concanavalin A (250 μg mL-1) and wheat germ agglutinin (100 μg mL-1) after in vivo treatment with a single dose of DPE (20 mg kg-1) for different intervals of time. C represented control without DPE |
Maximum decrease of agglutination with concanavalin A was observed at a dose of 100 and 200 μg mL-1 during 30 and 60 min of incubation (Fig. 5b, c), while maximum decrease of agglutination with wheat germ agglutinin was noted at a dose of 100 and 200 μg mL-1 during 30 min of incubation (Fig. 6b).
As compared to the control values, in vivo DPE-treatment at a single dose of 20 mg kg-1 showed decreased degree of normal and transformed cell agglutination with wheat germ agglutinin (Fig. 7a, b). Decreased degree of transformed cell agglutination was also observed with concanavalin A, while increased normal cell agglutination was noted with concanavalin A after 2 to 4 h of treatment (Fig. 7a, b). In DL cells, maximum decrease in the degree of cell agglutination with concanavalin A and wheat germ agglutinin were observed after 4 h of DPE treatment (Fig. 7b).
DISCUSSION
In the antitumor activity studies, ascites Dalton`s lymphoma has been commonly used as an important murine experimental tumor model (Prasad and Giri, 1974; Nicol and Prasad, 2002). Among different doses of aqueous and methanol extracts studied, methanol extract of D. pentagyna at a dose of 20 mg kg-1 showed comparatively better antitumor potential against murine ascites Dalton`s lymphoma. As compared to the regular increase in the body weight and ascites tumor volume of tumor-bearing control mice, the observed significant decrease in the body weight and ascites tumor volume of DPE-treated mice may indicate an inhibitory effect of D. pentagyna on tumor growth.
It has been reported that sialic acid influences many properties of the cell surface such as the determination of cell surface negativity and the loss of contact inhibition during malignancy and antigen masking agent. Cell surface glycoproteins and glycolipids are susceptible to such elevations as soon as a malignant growth starts to develop and metastasize, that they are referred to as tumor markers (Stringou et al., 1992) and also a decrease in the sialic acid content of cells causes a decrease in the negative charge of the molecule and this changes the interactions of cells with components inside and outside of the cells (Serdar et al., 2002). The present findings showed an increase in sialic acid concentrations in DL cells during tumor growth which may be due to enhanced activity of enzymes involved in sialic acid synthesis. Some reports have also indicated increased level of sialyl transferase activity in various virally transformed cells as compared to the corresponding normal cells, an event that may be associated with the increase in the amount of sialic acid in the transformed cells (Onodera et al., 1976). The elevated sialic acid levels in malignant cells have also been observed for murine Yoshida ascites sarcoma (Rao and Sirsi, 1973). DPE treatment decreases sialic acid content of normal lymphocytes in tumor-bearing control bringing back closer to that of normal value. The decrease in sialic acid content in DL cells after DPE treatment may be associated with an increase in tumor cell immunogenicity thereby enhancing host`s immune response or it may have a role in enhancing DL cells antigenicity, thus facilitating immunological recognition. DPE-mediated increase in the sialic acid content in the SN should be associated with the release of sialic acid moieties from DL cells. The increase in the number of leukocytes in tumor cell population after DPE treatment suggests the infiltration of many leukocytes towards tumor cells and the plasma membrane disintegration observed after 96 h of DPE treatment could lead to the lysis of tumor cells. The formation of membrane vacuoles on the tumor cells (Fig. 4e) following DPE treatment could be an indication of tumor cell lysis, eventually leading to cell death.
The sialic acid present at the surface of animal cells are of great interest because of their possible role in recognition phenomena. Thus, lectins which exhibit specific interactions with sialic acid moieties have been extensively used to monitor changes in cell surface carbohydrates/sialic acids. Lectins are oligomeric proteins or glycoproteins of non-immune origin with specific binding affinities for the carbohydrate moieties of glycoconjugates, widespread in all living organisms, from microbes and invertebrates to plants and vertebrates and are key players in many recognition events involved in fertilization, embryogenesis, metastasis and host-parasite recognition (Imberty et al., 2000). Alteration in the structure of cell membrane and cell surface sialic acid has been found to accompany the neoplastic transformation of cells and differential expression of glycoconjugates has been identified during malignant transformation (Keiko et al., 2004; Iwakawa et al., 1996; Remani et al., 2000). Various studies have shown altered carbohydrate composition in colorectal adenomas and carcinomas (Patricia et al., 2004) and altered cell surface sialic acid in DL cells has also been reported (Prasad and Sodhi, 1982). It has also been suggested that receptors for lectins are exposed in transformed cells whereas in normal cells some of the lectin binding sites are hidden which become exposed after treatment with proteolytic enzymes (Prasad and Sodhi, 1982) and removal of terminal sialic acids by sialidase has been reported to augment sensitivity to apoptosis induction in various cell lines (Peter et al., 1995). This alteration in surface properties has been studied by using plant lectins such as concanavalin A and wheat germ agglutinin which preferentially agglutinate transformed cells or normal cells after proteolytic enzyme treatment.
The results of present investigations suggest that D. pentagyna extract (DPE) may have a definite effect on the surface of both normal and transformed lymphocytes with reference to the lectin binding sites. This result is supported by the fact that normal lymphocytes (splenocytes), which showed a very low degree of agglutination with concanavalin A, when treated with DPE showed higher degree of agglutination with concanavalin A. On the other hand, a higher degree of agglutination of normal lymphocytes with wheat germ agglutinin was decreased after DPE treatment. Dalton`s lymphoma cells agglutinate to a very high degree with concanavalin A and wheat germ agglutinin. When DL cells were treated with DPE in vitro and in vivo, the degree of cell agglutination with concanavalin A and wheat germ agglutinin decreased significantly. Present investigations showing increase and decrease of cell agglutination of normal and transformed cells with concanavalin A after DPE treatment suggest that it is probably due to the removal of terminal sialic acid from normal lymphocytes resulting to the exposure of lectin binding sites, particularly concanavalin A binding sites and removal of lectin binding sites which are already exposed in transformed cells. This suggestion is also supported by the finding that in vivo DPE treatment decreases DL cell sialic acid content while an increased level of sialic acid was noted in ascites supernatant. The observed DPE-mediated decrease in agglutination of normal lymphocytes with wheat germ agglutinin may probably be due to the removal of wheat germ agglutinin binding sites along with the release of sialic acid from the cell surface.
It can be suggested that decrease in sialic acid content in DL cells after DPE treatment in vivo may play a role in the antitumor activity of D. pentagyna and the release of sialic acid from the surface of DL cells and decrease in the degree of DL cell agglutination with concanavalin A and wheat germ agglutinin after DPE treatment in vivo suggest that topographical changes does occur on the cell surface, which could possibly render them more immunogenic or accessible to the cells of immune system in the hosts.
ACKNOWLEDGMENTS
The financial support provided by the University Grants Commission, New Delhi, under Rajiv Gandhi National Fellowship (UGC. No. F-14-91/2006(SA-III) to G. Rosangkima and University with Potential for Excellence programme (F-1A/UPE/Biosc/APPT/2007/14) to T. Rongpi is duly acknowledged.
REFERENCES
- Alasbahi, R.H., S. Safiyeva and L.E. Craker, 1999. Antimicrobial activity of some Yemeni medicinal plants. J. Herbs Spices Med. Plants, 6: 75-83.
CrossRef - Filipovic, I. and E. Buddecke, 1979. Role of net charge of low-density lipoproteins in high affinity binding and uptake by cultured cell. Biochem. Bioph. Res. Co., 88: 485-490.
CrossRef - Imberty, A., C. Gautier, J. Lescar, S. Perez, L. Wyns and R. Loris, 2000. An unusual carbohydrate binding site revealed by the structures of two Maackia amurensis lectins complexed with sialic acid-containing oligosaccharides. J. Biol. Chem., 275: 17541-17548.
PubMed - Iwakawa, K., N. Ueda, S. Murao and N. Kobayashi, 1996. Altered carbohydrate composition in colorectal adenomas and carcinomas: Histochemical characterization of N-acetylgalactosamine, L-fucose and o-acetylated sialic acid. J. Gastroenterol., 31: 24-32.
CrossRef - Keiko, M., O. Katsuyuki, I. Mineko, K. Tetsufumi and K. Kensuke et al., 2004. Loss of disialyl Lewisa, the ligand for lymphocyte inhibitory receptor sialic acid-binding immunoglobulin-like lectin-7 (siglec-7) associated with increased sialyl Lewisa expression on human colon cancers. Cancer Res., 64: 4498-4505.
PubMed - Kelm, S. and R. Schauer, 1997. Sialic acids in molecular and cellular interactions. Int. Rev. Cytol., 175: 137-240.
CrossRef - Onodera, K., N. Yamaguchi, T. Kuchino and Y. Aoi, 1976. Alterations in surface glycoproteins and level of sialyltranferase of cells transformed by a temperature-sensitive mutant of SV40. Pak. J. Natl. Acad. Sci. USA, 73: 4090-4094.
Direct Link - Orekhov, A.N., V.V. Tertov, D.N. Mukhin and I.A. Mikhailenko, 1989. Modification of low density lipoprotein by desialylation causes lipid accumulation in cultured cells: discovery of desialylated lipoprotein with altered cellular metabolism in the blood of atherosclerotic patients. Biochem. Bioph. Res. Co., 162: 206-211.
CrossRef - Patricia de Albuquerque, G.R., C.V. Nakamura, W. De Souza and J.A. Morgado-Díaz, 2004. Differential expression of sialic acid and N-acetylgalactosamine residues on the cell surface of intestinal epithelial cells according to normal or metastatic potential. J. Histochem. Cytochem., 52: 629-640.
PubMed - Peter, M.E., S. Hellbardt, R. Schwartz-Albiez, M.O. Westendorp and H. Walczak et al., 1995. Cell surface sialylation plays a role in modulating sensitivity towards APO-1-mediated apoptosic cell death. Cell Death Differ, 2: 163-171.
PubMed - Prasad, S.B., 1986. Studies on the sialic acid in tumor and normal cells using cisplatin as a probe. Curr. Sci. India, 55: 651-654.
Direct Link - Prasad, S.B. and A. Giri, 1974. Antitumour effect of cisplatin against murine ascites Dalton's lymphoma. Indian J. Exp. Biol., 32: 155-162.
PubMed - Prasad, S.B. and A. Sodhi, 1982. Effect of cis-dichlorodiammine platinum (II) on surface of tumor and normal cells: Biochemical, fluorescence and electron microscopical studies. Indian J. Exp. Biol., 20: 559-571.
PubMed - Pulcinski, M.C., W.M. Riley, J.J. Prorok and J.A. Alhadeff, 1986. Total and lipid associated serum sialic acid levels in cancer patients with different primary sites and differing degrees of metastatic involvement. Cancer, 58: 2680-2685.
CrossRef - Rao, V.S. and M. Sirsi, 1973. Studies on sialic acid in Yoshida ascites sarcoma cells. Indian J. Biochem. Biol., 10: 37-41.
PubMed - Remani, P., R.A. Nair, T.T. Sreelekha, J. Madhavan, T. Vijayakumar and M.K. Nair, 2000. Altered expression of jack fruit lectin specific glycoconjugates in benign and malignant human colorectum. J. Exp. Clin. Cancer Res., 19: 519-523.
PubMed - Rosangkima, G. and S.B. Prasad, 2004. Antitumour activity of some plants from Meghalaya and Mizoram against murine ascites Dalton's lymphoma. Indian J. Exp. Biol., 42: 981-988.
PubMed - Sakagami, H., M. Ikeda, S. Unten, K. Takeda and J.I. Murayama et al., 1987. Antitumor activity of polysaccharide fractions from pine cone extract of Pinus parviflora Sieb. Et Zucc. Anticancer Res., 7: 1153-1160.
PubMed - Serdar, M.A., K. Yilmaz, A. Gunaydin, S. Tapan and K. Caglar et al., 2002. Sialic acid content of low-density lipoprotein in hemodialysis patients. Dialysis Transplant, 31: 97-105.
Direct Link - Stringou, E., K. Chondros, J. Kouvaris, S. Kakari and K. Papavassilious, 1992. Serum sialic acid (TSA/LSA) and carcinoembryonic antigen (CEA) levels in cancer patients undergoing radiotherapy. Anticancer Res., 12: 251-255.
PubMed - Tiralongo, J. and R. Schauer, 2004. The enigma of enzymatic sialic acid O-acetylation. Trends Glycoscience Glycotechnol., 16: 1-15.
Direct Link - Warren, L., 1959. The thiobarbituric acid assay of sialic acids. J. Biol. Chem., 234: 1971-1975.
PubMed - Warren, L., C.A. Buck and G.P. Tuszynski, 1978. Glycopeptide changes and malignant transformation. A possible role for carbohydrate in malignant behavior. Biochim. Biophys. Acta, 516: 97-127.
PubMedDirect Link