
Nicolas H. Badre
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, USA
Robin L. Cooper
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, USA
International Journal of Zoological Research
Year: 2008 | Volume: 4 | Issue: 3 | Page No.: 152-164







ABSTRACT
The temperature sensitive nature of a mutation in the Cacophony gene, which codes for the alpha subunit in the voltage-gated Ca2+ channel, reduces Ca2+ influx when exposed to non-permissive temperatures. We investigated the subtle nature in the impact for this mutation on whole animal function, in regards to learning and memory, in larvae and adults. The effects in acutely reducing evoked Ca2+ influx in nerve terminals during various behavioural assays greatly decreased the ability of larval Drosophila to learn, as demonstrated in associative learning assays. These assays are based on olfaction and gustation with association to light or dark environments with negative reinforces. Adult flies also showed defects in olfaction and sense of light when the animal is acutely depressed in normal Ca2+ influx within the nervous system. We demonstrated that this particular mutation does not alter cardiac function acutely. Thus, implying that the alpha 1 subunit mutation which retards neuronal function is not relevant for the pace maker and cardiac contractility as indexed by heart rate.
PDF Abstract XML References Citation
How to cite this article
Nicolas H. Badre and Robin L. Cooper, 2008. Reduced Calcium Channel Function in Drosophila Disrupts Associative Learning in Larva and Behavior in Adults. International Journal of Zoological Research, 4: 152-164.
DOI: 10.3923/ijzr.2008.152.164
URL: https://scialert.net/abstract/?doi=ijzr.2008.152.164
DOI: 10.3923/ijzr.2008.152.164
URL: https://scialert.net/abstract/?doi=ijzr.2008.152.164
INTRODUCTION
The normal function of ion channels, such as Ca2+ channels, within a given tissue can range from being essential for cell survival to only slight disturbances in function depending on the cells requirements. Many of the known diseases related to ion channel function are due to mutational effects (Mulley et al., 2003). Some of these pathologies are not severe enough to be lethal in embryological development but will allow survival for a time albeit with a decreased quality of life. The use of genetic mutations in animal models allows one to investigate the broad actions of such mutations in various bodily systems over acute and chronic periods depending on the severity of the mutation and the functional needs of the animal. In addition, one can examine the occurrence of a specific mutated gene within a give population under various selective pressures.
The Cacophony gene (cac-recessive mutation) in Drosophila codes for the alpha1 subunit of a voltage-gated calcium channel. This gene is known to be expressed in neural tissue (Tomancak et al., 2002). This subunit produces the pore of the voltage-gated ion channel. With the use of a transgene to express Green Fluorescent Protein (GFP) with cac-encoded alpha1 subunit it was demonstrated that the protein is present at motor nerve terminals in Drosophila (Kawasaki et al., 2004). A mutation in this segment, has also been characterized in a temperature sensitive line (cacTS2) in which, Ca2+ influx is reduced in the nerve terminal due to a conformational alteration in the channel when the temperature is raised (Brooks et al., 2003; Dellinger et al., 2000; Kawasaki et al., 2000; Macleod et al., 2006). As expected, synaptic transmission is reduced at the neuromuscular junction for cacTS2 (Kawasaki et al., 2000; Rieckhof et al., 2003; Xing et al., 2005). The cac TS2 adults exposed to 38°C for 3 min stop flying and walking which implies the alterations measured electro-physiologically in the skeletal muscles of larvae also apply to motor units in skeletal muscles of adults (Kawasaki et al., 2004) and possibly depresses other sites of synaptic communication centrally. Mutations that are temperature sensitive provide a good avenue to study the mechanisms of how homeostasis of an organism is regulated over time. Whereas complete knockouts of gene function can sometimes be lethal, the temperature sensitive mutational forms can be pulsed for various increments of time to assess acute and chronic effects. These more subtle mutations also mimic various types that are present in more complex animals.
Here we used the cacTS2 strain to investigate the potential effects on learning and memory through associative learning assays in larvae. These learning paradigms make use of vision, gustatory and olfaction in larval Drosophila, where as in adults we examined vision and olfaction behavioral responses with acute dysfunction of the channel. To delineate the properties of the cacTS2 strain related to heart function, since voltage-gated calcium channels play an integral role in cardiac function, alterations in heart rate was examined at permissive and restrictive temperatures. As our knowledge of physiological mechanism of neural plasticity in Drosophila largely derives from experiments in the larva (Koh et al., 2000), it is relevant to look at learning behavior in larvae with an alteration in Ca2+ function at the synaptic level. Since associative learning assays have now been established for larvae, these assays can be used in context with an alteration in a multitude of channels. The two larval learning assays we used complement each other; however, one is a gustatory-visual relationship while the other is an olfactory-visual paradigm (Gerber et al., 2004; Hendel et al., 2005; Scherer et al., 2003).
Clinical approaches are used to regulate calcium channel function as a means to control pathological conditions such as epilepsy and convulsions (Araújo et al., 2003). The outcome of long-term treatments of calcium channel blockers for regulation of such diseases needs to be addressed in terms of potential consequences in which homeostatic synaptic mechanisms may be compromised. This is relevant for the cacophony gene since it has recently been shown to be important in rapid homeostatic modulation of transmitter release (Frank et al., 2006). Possible by examining effects on the long-term reduction of calcium influx and by simply revealing the mechanism induced by calcium blocking in defined model systems will allow insight for clinical application as well as why some subtle mutations remain in the heritable genome.
MATERIALS AND METHODS
Drosophila Strains and Rearing
Canton S (CS) flies, a wild type of Drosophila, were used for comparison to cacTS2 (Kawasaki et al., 2004). Both strains were maintained at 20-21°C on a standard cornmeal-dextrose-agar-yeast medium. The cacTS2 strain was provided by Dr. Richard Ordway (The Pennsylvania State Univ.). These studies were conducted in 2005 through 2006 in Lexington, KY. To obtain staged larvae, flies were pulsed to lay eggs for 2 h and then the eggs were incubated at 21°C for 4 days. Heat shock experiments were performed at 33°C.
For the associative learning tests, both training and testing were done at the restrictive temperature. Impaired locomotion was accounted for since larvae are placed on the border of the light and dark regions and accounted for in the final test in a choice test. In addition, the paradigm tests for a difference in choice so if no difference is detected but the larvae still move on one or another environment a choice is made.
Learning Paradigms: Straight Alleyway
Mazes are often used to measure perception of different mutants (Störtkuhl et al., 2005). The experimental chamber we used was not a T-maze but a straight alleyway as the adult flies had to choose to go one direction or the other from a center point. The exact measurement of the alleyway chamber used for this study is shown in Fig. 1. Besides being able to have differential degrees of lighting on either side, olfactory cues can be placed at ends. In olfactory based paradigms, snug fitting, 0.43 g sponges were used which contained either 2 mL of water or 2 mL of 1/500 ethyl acetate (ethyl acetate anhydrous 99.5%, EM Science EX0240-9). The small hole between the middle tube and the ends is just big enough for a fly to land and move across the hole to then take off into the end tube. The flies do not just fly through the hole. Between 50 to a 100 flies were used in each alleyway trial.
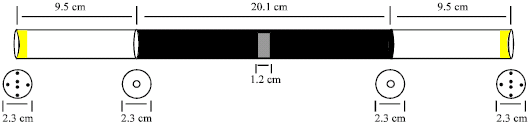 | |
| Fig. 1: | Experimental design for examining adult behaviors related to vision and olfaction. Standard Drosophila culture vials are used for this design. Two are placed together with their open ends facing each other for the center compartment. A wooden tongue depressor is taped across one side. Black electrical tape is used to cover the two center tubes with leaving a hole in the gap. Aluminum foil is used in the gap to wrap around the hole after placing flies into the chamber. The ends of the two center vials have a hole in the center (2 mm dia.). Two vials are taped, with standard office opaque tape, with the open side facing the ends of the center chamber. These end vials have several small holes (each 1 mm dia) at the most distal ends. Sponges can be placed in these end vials for testing preference to odors |
For each test, the flies were placed on the center of the alleyway, after which the opening was closed. The flies are left trapped in the alleyway for 20 min. Afterwards carbon dioxide is place in the chamber or the entire apparatus in place in the -20°C freezer to prevent the flies from moving anymore. The number of flies on each side was counted. For each series of tests, one sidepiece and the centerpiece are kept constant and the other is changed as can be shown in Fig. 2. In the vision test, only the wrapping of the sidepieces were changed whereas in the olfaction test, the tubes with liquids applied to the sponges were changed.
There is not likely any significant difference in temperature on the different sides due to radiant heat generated by light as the light was provide by overhead room florescent light tubes approximately 6 feet above the testing table.
Larval learning: Gustatory-Vision
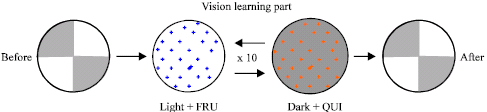
The gustatory-vision associated learning test followed a paradigm described earlier (Hendel et al., 2005) for gustatory reinforcement with some slight modifications. We primarily used 2nd instar instead of 3rd instar. All the larvae used in this test were raised at 18°C. In each learning experiment, we compared individual animals that underwent one of the training regimes. Larvae were first tested for their visual preference and then underwent a learning regime which included repeated positive gustatory phases in the light and negative gustatory phases in the dark (Fig. 3). For this experiment only 2nd instars were used since they normally prefer being in the dark. The reason for the negative reinforcer being associated with the dark side is because the 2nd instars prefer the dark and if we can switch their preference then one knows the larvae learned.
The 1% agarose plates were used for the initial and final visual (i.e., light/dark) preference test. For the learning assay, two other dishes were used: one for the positive gustatory reinforcement dishes with 1% agarose and 1 M fructose (FRU, purity: 99%) and one for the negative reinforcement with 1% agarose and 0.2% quinine hemisulfate (QUI, purity 93%). The gustatory reinforcers were added after the agarose was dissolved and slightly cooled just prior to pouring the agar for the plates. Glass Petri dishes were used as the agarose plates in these studies (90 mm diameter).
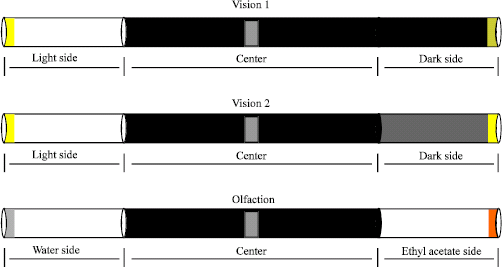 | |
| Fig. 2: | Behavioral tests for adult flies. The two vision paradigms made use of Vision 1 and Vision 2 alleyway designs. Alleyway for Vision 1 test had one of the end vials completely darkened by being wrapped with black electrical tape. For Vision 2, one vial at the end of the alleyway had 5 wrappings of standard opaque office supply tape. Examination of preference based on olfaction, sponges were placed into the ends of the vials and then tapped to the center piece immediately prior to the addition of flies to the central chamber. The whole apparatus is then placed in a functional fume hood so that air is lightly drawn out both ends of the apparatus |
 | |
| Fig. 3: | Associative learning assay for larva based on gustatory-vision cues. Plates are made with 1% agarose. For positive reinforcement 1 M fructose (FRU, purity: 99%) is used and for the negative reinforcement with 0.2% quinine hemisulfate (QUI, purity 93%) is used. The quadrants are made dark or light by a cut out of aluminum foil. The test are run on top of a light box, normally use to view slides. Since 2nd instar larva are used, they prefer the dark quadrant to start with but after associating dark with QUI their preference is changed |
All experiments were done in a dark room at room temperature (21°C). The dishes were placed on a light box that only allowed light to go through on the two quarters of the dishes. The learning regime consisted of 1 min in the light with a positive gustatory reinforcement, followed by 1 min in the dark with a negative gustatory reinforcement. This procedure was repeated 10 times. The learning assay takes longer than 20 min since the 1 min conditioning does not include the time for transferring the larvae. Transferring the larvae occurred as rapidly as possible (~10 sec). For the visual preference test, the larvae are recorded every 30 sec as being either on a dark side or on a light side for a total of 5 min. The % of larvae before and after training on the various dark/light locations are then compared for significant difference in learning.
 | |
| Fig. 4: | Associative learning assay for larva based on olfaction-gustatory cues. Plates are made with 1% agarose. Larvae are examined for their olfactory preference and then exposed to the learning paradigm with a gustatory cue to fructose and a positive association to 1-octanol (99%). The larvae are switched to a negative gustatory cue with QUI associated with a negative olfactory cue to 1:50 amyl acetate (99%). After repeat exposures back and forth the larvae are reexamined to the initial olfactory cues to index a change in preference. Modified from Scherer et al. (2003) |
Larval Learning: Olfaction-Gustatory
The olfaction-gustatory associative learning test followed Scherer et al. (2003) for gustatory reinforcement with some slight modifications. In the test we performed, we used 2nd instars that were raised at 18°C. In each learning experiment, we compared individual animals that underwent one of the training regimes. The larvae were first tested for their olfactory preference and then underwent a learning regime which included repeated positive gustatory phases with 1-octanol (99%) and negative gustatory phases with 1:50 amyl acetate (99%) in paraffin oil (Fig. 4).
The 1% agarose plates were as described above for the gustatory-vision associated learning test. All experiments were conducted in a dark room at room temperature (21°C). Ten microliter of each odorant was placed on pieces of filter paper that were placed on the opposite sides of the Petri dishes. The learning regime consisted of 1 min exposure to 1-octanol with a positive gustatory reinforcement, followed by 1 min exposure to amyl acetate with a negative gustatory reinforcement. This procedure was repeated 10 times (the learning takes longer than 20 min as the 1 min does not include the time for transferring the larvae). For the olfactory preference test, the larvae are recorded every 30 sec as being either on the 1-octanol side or on the amyl acetate side for a total of 5 min. The location of the larvae on the dish before and after training is then compared for significant difference from the half-way mark on the dish.
Heart Rate Monitoring
The movement of the trachea is commonly used to monitor Drosophila larval heart rate because of the clear contrast of the structures (Dasari and Cooper, 2006; White et al., 1992). The larval heart is viewed with the aid of a microscope (adjustable zoom 0.67 to 4.5; World Precision Instrument) fitted with a 10X eye objective and a 2x base objective. In addition, the images were recorded on to VHS tape by the use of a microscope-mounted camera (Mintron, MTV; World Precision Instrument). The movements of the trachea or heart were used for direct counts.
An ant farm apparatus was used to monitor the heart rate as this technique was shown not to induce stress in the animal and was not affected by increasing temperatures (Cooper and Cooper, 2004; Dasari and Cooper, 2004). Larvae raised at 21°C, were placed in an incubator at 33°C and the heart rate was measured every 5 min for 30 min. Five larvae were examined in each CS and cac strains.
All statistical analyses used are presented in the results for the particular measurements used.
RESULTS
Since the study by Chan et al. (2002) showed impaired locomotor behaviors of cacTS2 the % difference in CS and cacTS2 flies that choose one side over the other are used instead of absolute numbers since at the restrictive temperature a number of flies remained in the center of the alleyway apparatus.
Alleyway Maze: Vision 1 Protocol
Mutants and wild type showed similar behavior at room temperature. However, when the temperature is raised a significant shift occurs in the mean response between the mutant and the wild type populations (Fig. 5). At the restrictive temperature, 67.27% (±1.87) of CS and only 38.87% (±2.03) of cac went to the light side. This difference, while significant, does not necessarily represent a difference in preferences to light, as CS and cac showed no difference in the percent that went to the light side at room temperature; however, at 33°C there was a decrease of cac moving into the light side and an increase in preference to the dark side. There was no difference between CS and cac (1.07±0.1% of CS and 1.08±0.34% of cac) staying in the center tube at 33°C. In short, cac showed less preference to the light as compared to CS and among themselves for the dark side when tested at the restrictive temperature. Some flies did not move at all and thus remained in the center tube. The explanation for the lack of flies in the two ends could be explained by the fact that at higher temperatures, the mutant flies cannot move properly. To answer if this was the rational for the difference, we developed the second vision test which had a smaller difference in the light exposure between the two sides thus making a dim lit side and a regular lit side.
Alleyway Maze: Vision 2 Protocol
As for the previous alleyway maze (vision 1 protocol) there is no difference in preference of the mutants and wild type for the lighted and semi-lighted ends at room temperature (Fig. 6). However, as in the vision 1 protocol, once the temperature was raised the behavior of mutant and the wild type were significantly different for their preference for the lighted and semi-lighted tubes. In room temperature 47.26% (±0.88) of CS and 44.44% (±0.63) of cac went to the bright lit side. At 33°C, 44.86% (±1.91) of CS and 27.24% (±1.79) of cac went to the bright lit side. This test clearly shows a significant difference between mutants and wild type`s preference to light at higher temperature when the difference in lighting is subtle. The trend of the cac at room temperature in comparison to heat shock would appear that cac shunned the light as they stayed in the center (dark) or tended to move to the semi-lit (labeled as dark in Fig. 6) tube on the far extreme of the bright lit side. Possible the flies are processing information differently when light is present at the two ends as compared to one end. Vision test 2 as compared to vision test 1 does demonstrate that cac at high temperature move out of the center tube.
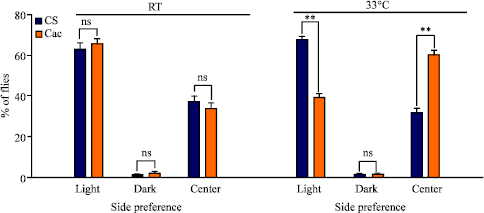 | |
| Fig. 5: | Light preference for CS and cac adults in the Vision 1 alleyway apparatus. Difference between wild type and cac preference to light at room temperature (21°C) and 33°C. Comparisons are made between CS and cac. (N = 445 total number of flies tested; *p<0.05; **p<0.001; ***p<0.0001; ns is not significant) |
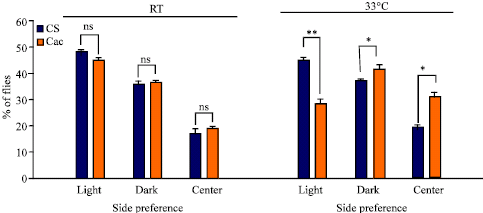 | |
| Fig. 6: | Light preference for CS and cac adults in the Vision 2 alleyway apparatus. Difference between wild type and cac preference to light at room temperature (21°C) and 33°C. Comparisons are made between CS and cac. (N = 639 total number of flies tested; *p<0.05; **p<0.001; ***p <0.0001; ns is not significant) |
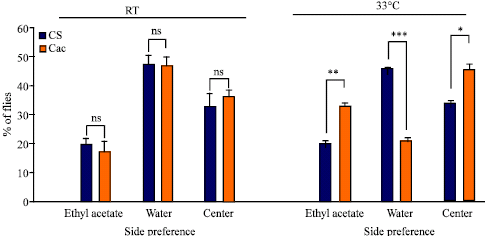 | |
| Fig. 7: | Olfactory preference for CS and cac adults in the alleyway apparatus. Difference between wild type and cac preference to light at room temperature (21°C) and 33°C. The concentration of ethyl acetate (1/500) used here acts as a repellant. Comparisons are made between CS and cac. (N = 415 total number of flies tested; *p<0.05; **p<0.001; ***p <0.0001; ns is not significant) |
Alleyway Maze: Olfaction
Similarly cac and wild type showed no difference in olfactory based behavior at room temperature; however, once the temperature was raised then differences were apparent (Fig. 7). At room temperature, 20.36% (±0.77) of CS and 33.22% (±1.13) of cac are attracted to the ethyl acetate side. A large shift in behavior occurred at 33°C with 45.75% (±0.41) of CS and 21.25% (±0.78) of cac being repelled by ethyl acetate or attracted to the water side. This test clearly shows a significant difference between mutants and wild type`s preference to ethyl acetate at higher temperature. One interpretation of these results is that at room temperature ethyl acetate is acting as a repellent for both CS and cac however at higher temperature it is still a repellent for CS but not for cac. This may well be that cac cannot effectively smell ethyl acetate as well due to a decreased processing of sensory information and if this is the case then ethyl acetate may even be sensed as an attractant. This would make sense since at low concentration ethyl acetate is an attractant but at the high concentration, as we used, it is a repellent. Thus, with decreased integration of sensory input of cac, the high concentration of ethyl acetate would appear weaker.
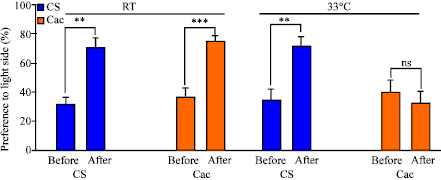 | |
| Fig. 8: | Effects of associative learning in 2nd instar larvae based on gustatory and vision. The assay examines the preference to light in CS and cac larva that are conditioned at 21°C or 33°C. Comparisons are made before and after conditioning. (N = 24; *p<0.05; **p<0.001; ***p <0.0001; ns is not significant) |
The short time frame does not take into account the possibility of increased adaptation or habituation to the odor. Once flies go through the small hole, from the center tube into the side tubes they do not go back into the dark center tube. We were not concerned if they habituated once within the side tubes but just the fact that they choose one side over the other was sufficient.
Larval Associative Learning: Vision and Gustatory
The cac and CS both demonstrate similar abilities to associatively learn based on light with positive and negative gustatory reinforces as the preference to light is substantially increased at room temperature after the training period (Fig. 8). At higher temperature, CS larvae still responded to the learning assay as previously seen for room temperature. However, cac did not learn as well at a higher temperature. This may well be related to processing of complex sensory tasks associated with the learning assay considering neuronal communication is compromised, as one would expect.
Larval Associative Learning: Olfaction and Gustatory
As with the vision-gustatory learning paradigm, the olfactory-gustatory associative learning worked sufficiently well to demonstrate learning at room temperature for both CS and cac larvae (Fig. 9). With disturbed synaptic transmission induced at higher temperature for the cac strain it is not surprising that a learning deficiency is apparent. The CS strain continued to demonstrate a learning ability at the same temperature. Thus, the complex central processing is not functional for the cac to learn when the mutation is manifested.
Heart Rate
Even though cac larvae reduce locomotion during a heat pulse, their heart rate continues to beat over long periods of time. The heart rate appears to be sometimes irregular but on average the number of beats is not significantly different to wild type larvae at room temperature and during a heat pulse (Fig. 10). The cac group had a decrease in beats per minute on average of -9.15 (±1.97 SEM), where the CS had an average decrease in beats per minute of -4.64 (±1.49) after heat shock. These results imply that the subtype of voltage-gated Ca2+ channels in the heart is still functioning in larvae even after 30 min of heat shock.
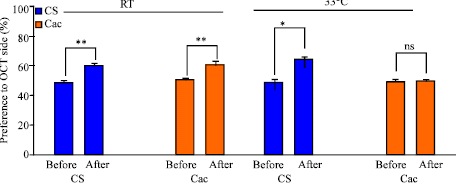 | |
| Fig. 9: | Effects of associative learning in 2nd instar larvae based on gustatory and olfaction cues. The assay examines the preference to olfactory cues for CS and cac larva after they are conditioned at 21°C or 33°C. A percent difference is shown. (N = 42; *p<0.05; **p<0.001; ***p <0.0001; ns is not significant) |
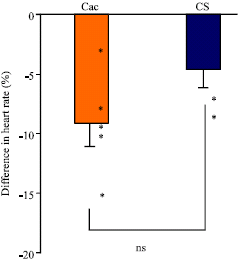 | |
| Fig. 10: | The % difference in heart rate for CS and cac strains when measured at 21°C and after exposure to 33°C for 30 min. The individual values are shown as circles. (N = 5; ns is not significant) |
DISCUSSION
In this study we demonstrated that the Drosophila strain, cacTS2, that is defective in Ca2+ influx at nerve terminals, does not perform well as a larva or as an adult in several different behavioral assays which are dependant on integrative abilities within the CNS. Simple sensory response behaviors to complex associative learning, involving multiple sensory modalities appear to be affected by this mutation. We also demonstrated that this particular mutation does not alter cardiac function acutely. Thus, implying that the alpha 1 subunit mutation which retards neuronal function is not relevant for the pace maker and cardiac contractility as indexed by heart rate.
All of the experimental paradigms presented in this study are during acute dysfunction in the calcium channel, which raises questions what would happen over longer periods, for example during larval or adult development. Would learning be impaired in the 3rd instar or in an adult if synaptic transmission was retarded only during a short period within the 1st instar? We are just now starting chronic and pulsed heat shocks throughout development in regards to these behavioral assays and learning paradigms. However, previous studies have been conducted in respect to chronic dysfunction of the associated calcium channels in motor nerve terminals (i.e., neuromuscular junctions) in regards to nerve terminal development and homeostatic compensation for reduced synaptic function. When larvae are raised at a restrictive temperature from 1st instar to 3rd instar, the motor nerve terminals do not grow properly and synaptic transmission at the NMJ does not undergo homeostatic compensation but instead remains depressed (Rieckhof et al., 2003; Xing et al., 2005). The mean quantal content per synaptic varicosity is even decreased which mechanistically could mean that synapses did not develop properly and/or that reduced Ca2+ influx during evoked responses with acute stimulation is still occurring in 3rd instars without a compensatory mechanism to regain normal synaptic function. The mechanism behind a reduced quantal output remains to be determined, but given that this occurs spurred us to investigate central neuronal function in larvae and adult flies. These prior studies used a relatively strong activation of the temperature sensitivity of the channel by using 29°C. This resulted in a drastic reduction of synaptic transmission. In a recent study that used a milder heat shock (22°C or shorter pulses at 30°C) but enough to reduce synaptic transmission, demonstrated an enhanced ability of the nerve terminal to regain function rapidly once brought back to permissive temperatures (Frank et al., 2006). This suggests a level of synaptic homeostasis when calcium is not as drastically reduced. The acute behavioral assays presented herein demonstrate the wide effects this mutation has on the animal. It is feasible to assume central synapses are not as functional for cac as compared to CS during a restrictive temperature based on the past evidence for the NMJs (Brooks et al., 2003; Dellinger et al., 2000; Frank et al., 2006; Kawasaki et al., 2000, 2002, 2004; Xing et al., 2005); thus, possibly explaining the observed phenomenon in behaviors we presented. Comparing behavior before versus after training as a measure of learning could lead one astray easily since such changes may come about by the passage of time, by sensory adaptation to the odours/ light and by motivational changes (Boyle and Cobb, 2005; Colomb et al., 2007; Gerber and Stocker, 2007; Rescorla, 1988). Also the effects are likely global from a reduction in sensory and motor neurons in addition to central interneurons.
In synaptic preparations that are used to model sensitization, a form of associative learning, intracellular calcium dynamics is known to be crucial in mediating cellular cascades that are established key players in a learned response (Alkon and Nelson, 1990; Li et al., 2005). Experimental paradigms using mammalian hippocampal slice preparations, in which Long-Term Potentiation (LTP) is induced, involves both pre- and post-synaptic localized calcium transients (Franks and Sejnowski, 2002; Lisman, 2003; Miyamoto, 2006). Similarly, the time frames in which gene transcription can impact a learning process, such as LTP, (Huang and Kandel, 2006) is within the 20 to 30 min training periods used in our studies. Calcium`s role in behavioral habituation and sensitization in defined neural circuits of Aplyisia has been shown to be mediating a calcium/calmodulin-activated protein kinase II (CaMKII) and regulate gene transcription through CREB (Bartsch et al., 1998; Kennedy et al., 1992). Since Ca2+ is a key player in many divergent cellular pathways it is not so surprising that even slight reductions in influx within pre- or post-synaptic sites will result in acute behavioral problems. The trigger of Ca2+ for broad secondary cascades such as CaMKII, CICR and cADPr (Emptage et al., 2001; Hachisuka et al., 2007) presents a challenge for defining the mechanisms in behavioral defects. A classic example for calcium involvement related to learning defects in Drosophila is the rutabaga (rut) mutant (Aceves-Piña et al., 1983). The rut gene codes for a Ca2+/calmodulin- stimulated adenylyl cyclase, so likely a disturbed calcium homeostasis could indirectly mimic some of the deficiencies demonstrated in rutabaga. It would be reasonable to target similar molecular mechanisms in the CNS of larval Drosophila since we have shown defective calcium channels produces behavioral and learning abnormalities. Since the olfaction and gustatory sensory projections are established in the larval brain (Python and Stocker, 2002; Tissot et al., 1997) the task now is to complete the central circuit related to information processing and learning and correlate with the molecular mechanisms responsible for learning and memory.
Many regions of the fly CNS are analogous to those with the human brain in the sense of processing centers with subdivision difference for perception and relay paths to particular motor command nuclei. To examine normal developmental processes, various degrees of perturbation in behavioral assays can help to determine the importance of particular neural pathways and the plasticity of the system in chronic as well as acute periods. Given that this mutation affects various neuronal and motor functions in flies, it is likely that the observed larval defects are caused by the defects in these peripheral functions as well as central brain functions. For learning and memory to occur sensory function is essential and is an integral process of learning and memory.Given that behavioral assays are used to access the learning, the motor unit function is as essential as sensory or CNS function. In our larval learning studies we did not examine non-associative phenomenon such as if both genotypes fed at the same rate during training. We folded such behaviors into the overall learning paradigm. One could argue, for example a genotype in humans that is defective in calcium channel function results in malnutrition and this genotype also do not learn as well. The malnutrition could be a direct result of the geneotype but in short there is also a learning deficit due to genotype as compared to the background strain; thus how the learning difference comes about in our study needs to be further examined.
The broader impact of this study is that it demonstrates a single gene mutation related to neuronal function can have pronounced learning and behavioral defects which would likely be selected out during natural selection in a wild population; however, the defective gene could be established within an environment that did not manifest the consequences of the defect (Darwin, 1859; Gerber et al., 2004).
ACKNOWLEDGMENTS
Support was provided by a G. Ribble Fellowship for undergraduate studies in the Department of Biology at the University of Kentucky (NB), a Undergraduate Research Scholarship awarded by the Arnold and Mabel Beckman Foundation (NB) and in part by NSF grant IBN-0131459 (RLC).
REFERENCES
- Aceves-Piña, E.O., R. Booker, J.S. Duerr, M.S. Livingstone and W.G. Quinn et al., 1983. Learning and memory in Drosophila, studied with mutants. Cold Spring Harb. Symp. Quant. Biol., 48: 831-840.
CrossRefPubMedDirect Link - Alkon, D.L. and T.J. Nelson, 1990. Specificity of molecular changes in neurons involved in memory storage. FASEB J., 4: 1567-1576.
Direct Link - Araújo, D.A., R.A. Mafra, A.L. Rodrigues, V. Miguel-Silva and P.S. Beirao et al., 2003. N-Salicyloyltryptamine, a new anticonvulsant drug, acts on voltage-dependent Na+, Ca2+ and K+ ion channels. Br. J. Pharmacol., 140: 1331-1339.
Direct Link - Bartsch, D., A. Casadio, K.A. Karl, P. Serodio and E.R. Kandel, 1998. CREB1 encodes a nuclear activator, a repressor and a cytoplasmic modulator that form a regulatory unit critical for long-term facilitation. Cell, 95: 211-223.
CrossRefDirect Link - Boyle, J. and M. Cobb, 2005. Olfactory coding in Drosophila larvae investigated by cross-adaptation. J. Exp. Biol., 208: 3483-3491.
Direct Link - Brooks, I.M., R. Felling, F. Kawasaki and R.W. Ordway, 2003. Genetic analysis of a synaptic calcium channel in Drosophila: Intragenic modifiers of a temperature-sensitive paralytic mutant of cacophony. Genetics, 164: 163-171.
Direct Link - Chan, B., A. Villella, P. Funes and J.C. Hall, 2002. Courtship and other behaviors affected by a heat-sensitive, molecularly novel mutation in the cacophony calcium-channel gene of Drosophila. Genetics, 162: 135-153.
Direct Link - Colomb, J., N. Grillenzoni, A. Ramaekers and R.F. Stocker, 2007. Architecture of the primary taste center of Drosophila melanogaster larvae. J. Comp. Neurol., 502: 834-847.
Direct Link - Cooper, A.S. and R.L. Cooper, 2004. Monitoring activity of Drosophila larvae: Impedance and video microscopy measures. Drosophila Inform. Ser., 87: 85-87.
Direct Link - Dasari, S. and R.L. Cooper, 2004. Modulation of sensory-CNS-motor circuits by serotonin, octopamine and dopamine in semi-intact Drosophila larva. Neurosci. Res., 48: 221-227.
CrossRefPubMedDirect Link - Dasari, S. and R.L. Cooper, 2006. Direct influence of serotonin on the larval heart of Drosophila melanogaster. J. Comp. Physiol. B, 176: 349-357.
CrossRefDirect Link - Dellinger, B., R. Felling and R.W. Ordway, 2000. Genetic modifiers of the Drosophila NSF mutant, comatose, include a temperature-sensitive paralytic allele of the calcium channel alpha1-subunit gene, cacophony. Genetics, 155: 203-211.
Direct Link - Emptage, N.J., C.A. Reide and A. Fine, 2001. Calcium stores in hippocampal synaptic boutons mediate short-term plasticity, store-operated Ca2+ entry and spontaneous transmitter release. Neuron, 29: 197-208.
Direct Link - Frank, C.A., M.J. Kennedy, C.P. Goold, K.W. Markek and G.W. Davis, 2006. Mechanisms underlying the rapid induction and sustained expression of synaptic homeostasis. Neuron, 52: 663-677.
Direct Link - Franks, K.M. and T.J. Sejnowski, 2002. Complexity of calcium signaling in synaptic spines. Bioassays, 24: 1130-1144.
Direct Link - Gerber, B., S. Scherer, K. Neuser, B. Michels, T. Hendel, R.F. Stocker and M. Heisenberg, 2004. Visual learning in individually assayed Drosophila larvae. J. Exp. Biol., 207: 179-188.
CrossRefPubMedDirect Link - Gerber, B. and R.F. Stocker, 2007. The Drosophila larva as a model for studying chemosensation and chemosensory learning: A review. Chem. Senses, 32: 65-89.
CrossRefPubMedDirect Link - Hachisuka, J., S. Soga-Sakakibara, M. Kubota, K. Narita and K. Kuba, 2007. Enhancement of Ca2+-induced Ca2+ release by cyclic ADP-Ribose in frog motor nerve terminals. Neuroscience, 146: 123-134.
Direct Link - Hendel, T., B. Michels, K. Neuser, A. Schipanski and K. Kaun et al., 2005. The carrot, not the stick: Appetitive rather than aversive gustatory stimuli support associative olfactory learning in individually assayed Drosophila larvae. J. Comp. Physiol. A, 191: 265-279.
CrossRefDirect Link - Huang, Y.Y. and E.R. Kandel, 2006. Age-related enhancement of a protein synthesis-dependent late phase of LTP induced by low frequency paired-pulse stimulation in hippocampus. Learn Mem., 13: 298-306.
Direct Link - Kawasaki, F., R. Felling and R.W. Ordway, 2000. A temperature-sensitive paralytic mutant defines a primary synaptic calcium channel in Drosophila. J. Neurosci., 20: 4885-4889.
Direct Link - Kawasaki, F., S.C. Collins and R.W. Ordway, 2002. Synaptic calcium-channel function in Drosophila: Analysis and transformation rescue of temperature-sensitive paralytic and lethal mutations of cacophony. J. Neurosci., 22: 5856-5864.
Direct Link - Kawasaki, F., B. Zou, X. Xu and R.W. Ordway, 2004. Active zone localization of presynaptic calcium channels encoded by the cacophony locus of Drosophila. J. Neurosci., 24: 282-285.
Direct Link - Kennedy, T.E., D. Kuhl, A. Barzilai, J.D. Sweatt and E.R. Kandel, 1992. Long-term sensitization training in Aplysia leads to an increase in calreticulin, a major presynaptic calcium-binding protein. Neuron, 9: 1013-1024.
CrossRefDirect Link - Koh, Y.H., L.S. Gramates and V. Budnik, 2000. Drosophila larval neuromuscular junction: Molecular components underlying synaptic plasticity. Microscopy Res. Technol., 49: 14-25.
Direct Link - Li, Q., A.C. Roberts and D.L. Glanzman, 2005. Synaptic facilitation and behavioral dishabituation in Aplysia: Dependence on release of Ca2+ from postsynaptic intracellular stores, postsynaptic exocytosis and modulation of postsynaptic AMPA receptor efficacy. J. Neurosci., 25: 5623-5637.
Direct Link - Lisman, J., 2003. Long-term potentiation: Outstanding questions and attempted synthesis. Philos. Trans. R. Soc. Lond. B Biol. Sci., 358: 829-842.
Direct Link - Macleod, G.T., L. Chen, S. Karunanithi, J.B. Peloquin and H.L. Atwood et al., 2006. The Drosophila cacts2 mutation reduces presynaptic Ca2+ entry and defines an important element in Cav2.1 channel inactivation. Eur. J. Neurosci., 23: 3230-3244.
Direct Link - Miyamoto, E., 2006. Molecular mechanism of neuronal plasticity: Induction and maintenance of long-term potentiation in the hippocampus. J. Pharmacol. Sci., 100: 433-442.
Direct Link - Mulley, J.C., I.E. Scheffer, S. Petrou and S.F. Berkovic, 2003. Channelopathies as a genetic cause of epilepsy. Curr. Opin. Neurol., 16: 171-176.
Direct Link - Python, F. and R.F. Stocker, 2002. Adult-like complexity of the larval antennal lobe of D. melanogaster despite markedly low numbers of odorant receptor neurons. J. Comp. Neurol., 445: 374-387.
Direct Link - Rescorla, R.A., 1988. Behavioral studies of pavlovian conditioning. Annu. Rev. Neurosci., 11: 329-352.
CrossRefDirect Link - Rieckhof, G.E., M. Yoshihara, Z. Guan and J.T. Littleton, 2003. Presynaptic N-type calcium channels regulate synaptic growth. J. Biol. Chem., 278: 41099-41108.
Direct Link - Scherer, S., R.F. Stocker and B. Gerber, 2003. Olfactory learning in individually assayed Drosophila larvae. Learn. Memory, 10: 217-225.
CrossRefDirect Link - Störtkuhl, K.F., R. Kettler, S. Fischer and B.T. Hovemann, 2005. An increased receptive field of olfactory receptor Or43a in the antennal lobe of Drosophila reduces benzaldehyde-driven avoidance behavior. Chem. Senses, 30: 81-87.
Direct Link - Tissot, M., N. Gendre, A. Hawken, K.F. Stortkuhl and R.F. Stocker, 1997. Larval chemosensory projections and invasion of adult afferents in the antennal lobe of Drosophila. J. Neurobiol., 32: 281-297.
PubMed - Tomancak, P., A. Beaton, R. Weiszmann, E. Kwan and S. Shu et al., 2002. Systematic determination of patterns of gene expression during Drosophila embryogenesis. Genome Biol.
Direct Link - White, L.A., J.M. Ringo and H.B. Dowse, 1992. Effects of deuterium oxide and temperature on heart rate in Drosophila melanogaster. J. Comp. Physiol. B, 162: 278-283.
CrossRefDirect Link - Xing, B., A.A. Long, D.A. Harrison and R.L. Cooper, 2005. Developmental consequences of NMJs with reduced presynaptic calcium channel function. Synapse, 57: 132-147.
Direct Link