
Maurice -Pierre Page
Department of Biology, University of Kentucky, Lexington, KY USA, 40506-0225
Walter Hailes
Department of Biology, University of Kentucky, Lexington, KY USA, 40506-0225
Robin L. Cooper
Department of Biology, University of Kentucky, Lexington, KY USA, 40506-0225
International Journal of Zoological Research
Year: 2007 | Volume: 3 | Issue: 3 | Page No.: 132-144







ABSTRACT
The neural circuitry used during the tail flip behavior of crayfish continues to serve as a model for investigating sensory integration and the actions of neuromodulators. Systemic injections of 5-HT produce an increase in responsiveness to sensory stimuli and decrease the rate of sensory habituation of the tail flip. In behaving animals the loss of the chelipeds also promotes a greater responsiveness to tail flipping on sensory stimulation of the telson. By transecting the Ventral Nerve Cord (VNC) it was demonstrated that the animals were less responsive to the tail flip reflex, implying that the sensory-VNC-motor circuit for the tail flip can be modulated in the animals by higher command neurons. However, the CNS isolated neuronal circuitry of the tail was still responsive to 5-HT. The results imply that there is direct CNS regulation as well possible hormonal action in intact behaving animals and that factors which alter one or the other could impinge on the tail flip escape behavior.
PDF Abstract XML References
How to cite this article
Maurice -Pierre Page, Walter Hailes and Robin L. Cooper, 2007. Modification of the Tail Flip Escape Response in Crayfish by Neuromodulation and Behavioral State with and without Descending CNS Input. International Journal of Zoological Research, 3: 132-144.
DOI: 10.3923/ijzr.2007.132.144
URL: https://scialert.net/abstract/?doi=ijzr.2007.132.144
DOI: 10.3923/ijzr.2007.132.144
URL: https://scialert.net/abstract/?doi=ijzr.2007.132.144
INTRODUCTION
Crayfish use the tail flip response as a means of evading predators and each other (Edwards et al., 1999; Krasne, 1969). The neuronal circuitry of the tail flip induced by a touch of the telson or the abdomen was one of the first neuronal circuits described (Johnson, 1924; Kennedy et al., 1969; Wine and Krasne, 1982) and is still an active preparation in understanding the circuitry and actions of neuromodulators. Earlier studies indicated that juvenile crayfish are more likely to tail flip, whereas an adult crayfish is more likely to respond to threatening stimuli with the defensive use of its claws. Fricke (1986) studied this behavioral phenomena in depth, showing that the change in responsiveness to a stimulus is due to size, not age. Fricke (1986) stated several reasons why the different responsiveness of small and large crayfish might be advantageous to the species: the relationship of cheliped mass to body mass, the energy required for a tail flip, the high predation on juveniles and the adults’ harder defensive shells. Thus, the smaller crayfish are ecologically selected for greater tail flipping abilities. In addition, Fricke (1986) postulated neurophysiological mechanisms through which the varying developmental responses might occur. Fricke (1986) noted the stable electronic synaptic input from tactile afferents weakens as crayfish grow.
Research by Don Edwards' group (Olson et al., 1996) developed this theme to show that the change in responsiveness of the tail flip response is a product of neuronal cable properties and synaptic efficacy. The rationale is that a depression-prone B synapse becomes more responsible for the tail flip than the A synapse because attenuation in the A synapse is greater in the adult. Additional findings (Herberholz et al., 2001; Krasne et al., 1997; Teshiba et al., 2001; Yeh et al., 1996) now show a link between the excitability of the tail flip response in dominant and submissive animals, which suggests a hormonal influence. It has been shown that the tail flip escape circuitry can excite neurons containing 5-HT within the Ventral Nerve Cord (VNC) in lobsters (Hörner et al., 1997). Earlier studies by Lang et al. (1977) first noted that when lobsters are induced to autotomize their chelipeds, a dominant animal will quickly switch to subordinate behavior. This integrates well with the suggestion that dominant and submissive animals have altered responses to sensory stimulation of the telson.
Recently, we demonstrated that the probability of the crayfish, P. clarkii, tail flipping in response to a touch on the dorsal tail fan is dependent on the size and the behavioral state (Kellie et al., 2001). We showed that a behavioral state can surpass the size dependence in the induced tail flip escape response. This was demonstrated by measuring the tail flip response before and after the animal autotomized its chelipeds. Larger sized animals would tail flip to a given stimulus more often when their chelipeds were lost. It is of interest to note that cheliped removal resulted in rapid alteration in tail flip behavior. This increased responsiveness of the animals remains for up to a week (Kellie et al., 2001). The past and current observations of a rapid reduction in tail flip habituation on removal of the chelipeds suggests a hormonal effect on the sensory-motor response contained within the abdomen, or possibly even a directly descending neural regulation of the localized circuit.
A very likely candidate hormone is octopamine (OA). OA-containing neurosecreting OA cells are likely to release OA into the hemolymph when the chelipeds are autotomized, since sensory neurons in the chelipeds project to these central neurons (Heinrich et al., 1999). In addition, OA is known to enhance primary sensory neuron input in the tail flip circuit (Bustamante and Krasne, 1991). Earlier investigations into serotonin's (5-HT) role in affecting crustacean behavior have been addressed, but there is no definitive proof to link it to specific naturally occurring behaviors, since a myriad of neuromodulators are present at any one time during similar behaviors (Sneddon et al., 2000) and given that the relative ratios or previous exposure to one neuromodulator can alter the response to another (Djokaj et al., 2001). In addition, it was recently shown that crustaceans depleted of 5-HT do not cease to be aggressive (Dornberg et al., 2001). This implies that earlier studies’ findings of a 1-to-1 relationship between neuromodulator and aggressive/submissive behaviors may have suffered from an over simplification of complex behaviors (Livingston et al., 1980; Yeh et al., 1996; Kravitz and Huber, 2003; Sneddon et al., 2000; Cooper et al., 2001; Listerman et al., 2000; Strawn et al., 2000; Tabor and Cooper, 2002; Pagé and Cooper, 2004).
Three main types of tail flips are used in indexing the behavioral repertoire of crayfish. The non-giant neuron mediated tail flip produces a slow tail flip response, usually in a series of tail flips (Johnson, 1924; Kennedy et al., 1969; Krasne, 1969; Wine and Krasne, 1982). This is generally initiated by a tactile stimulation on the side of the crayfish, or by a general environmental disturbance such as dropping a small object into the animal's holding tank (Listerman et al., 2000). The giant mediated responses refer to recruiting the giant neurons (called as such because they have large axon diameters which run the length of the ventral nerve cord). The two pairs of giant axons are recruited by different means. The medial giants are activated by a strong sensory stimulation in front of the animal, which results in the animal’s initiating rapid, powerful tail flips to propel itself backwards, away from impending danger. The lateral giant axons within the nerve cord were activated by a strong stimulus behind the animal: for example, a forceful touch on the dorsal part of the telson. This produces a tail flip that propels the animal up in the water column and forwards.
There is a now a growing consensus in the field that there are numerous neuromodulators working in concert to exert their influence on synaptic function, which ultimately shapes the behavior of the animal (Djokaj et al., 2001; Mesce, 2002; Ayali and Harris-Warrick, 1999). Given the limitation of investigating the many known neuromodulators and their ratios in relation to crustacean behavior, we chose to address the issue of whether the two commonly investigated neuromodulators, 5-HT and OA, would enhance or dampen the tail flip escape response in the whole animal. Since a decrease in tail flip habituation occurs with cheliped autonomy and is thought to be driven by hormonal actions, we examined also whether these neuromodulators would change behavior when the animals were induced to autotomize their chelipeds. In order to determine whether direct descending neural input is responsible for the hyperexcitability to the induced tail flip after cheliped removal, the descending tracts within the ventral nerve cord were severed and the animals’ consequent behavior was monitored against sham controls.
MATERIALS AND METHODS
Crayfish Care
All experiments were performed using Procambarus clarkii (Atchafalaya Biological Supply Co., Raceland, LA). The animals were held in individual tanks in the laboratory for at least 2 weeks before being used in experiments. Only male crayfish were used. The crayfish were fed dry fish food once a week. Prior to testing, the animals were kept in isolation tanks in the test room, which was illuminated 24 h a day with a dim, indirect white light. The crayfish were returned to their isolation tanks after being tested. The large crayfish used in this study ranged in weight from 12 to 21 g.
Testing Procedure 1
During the primary tests, the crayfish were moved from their isolation tanks to test tanks, which were approximately 30x32x13 cm, at least 24 h before being tested. The examiner remained still and as inconspicuous as possible to reduce visual disturbances, while still having the ability to observe the crayfish. Testing followed a similar approach as that of Kellie et al. (2001). The crayfish received a tap on the tail with a glass rod once every minute and in some experiments, every 30 sec (Fig. 1A). Testing continued until the crayfish failed to tail flip at least 5 consecutive times. To insure uniformity of stimulus the best care was taken to use the same force in hitting the tail, however some unavoidable varibility probably occurred as would be expected in such behavioral testing. After testing was completed the crayfish were returned to their isolation tanks and the water in the test tanks was replaced.
Testing Procedure 2
For examining the effects of systemic injections of 5-HT and OA, the animals were injected, through the lateral aspect of the articulating membrane, in the second ventral abdominal segment. They were injected with saline (sham) or with a 5-HT or OA solution to provide a circulating concentration of 10 μM as determined in proportion to their mass. Stock solutions of 1 mM were used to provide a 10 μM circulating concentration, given that hemolymph volume equals 30% of the crayfish mass (Gleeson and Zubkoff, 1977).
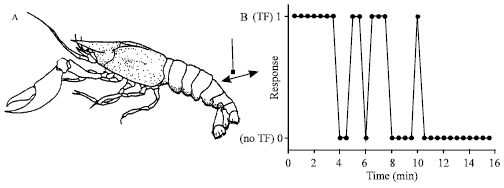 | |
| Fig. 1: | The lateral giant mediated tail flip response is induced by forcefully tapping the dorsal side of the telson (A), Here a fire-polished glass rod is shown to tap the dorsal aspect. When the crayfish exhibited a lateral giant mediated tail flip, a score of 1 was given and when a tail flip did not follow a tap on the telson, a 0 was recorded. The animal was tapped on the telson at 30 sec intervals and the scores were recorded to obtain an index of the rate of habituation to sensory stimulus (B) |
The crayfish saline was a modified Van Harreveld's solution (in mM: 205 NaCl; 5.3 KCl; 13.5 CaCl2.2H2O; 2.45 MgCl2.6H2O; 5 HEPES adjusted to pH 7.4 and injected through a syringe filter of 0.2 μm pore size). During the second series of tests in relation to cheliped removal, the crayfish were moved into the test tanks at least 24 h before testing. They were tested until the significant number of failures to tail flip were observed. The animals were left for 1 h to recover. They were injected with 5-HT, OA or saline, both their chelipeds were removed, then they were immediately retested. Saline was the vehicle used to inject 5-HT and OA. The crayfish were tested again one day and one week after the chelipeds were removed.
Testing Procedure 3
In a third set of experiments the neural input from the rostral centers to the abdominal segment 1 was examined for its role in maintaining large crayfish in a state responsive to tail flipping. This was examined by transecting the VNC between A1 and A2 of the first cuticular rib. Sham treatment consisted of making a cut in the ventral articulating membrane but not cutting the VNC. The connective tissue was cut to expose the VNC. The gap made by the incision was then closed with cyanoacrylate ester and accelerator (super glue; HobbyTown USA, Lexington, KY). The use of this rapidly drying glue reduced the stress of handling on the animals, which is known to have an effect on the heart rate of crayfish (Wilkens et al., 1985; Listerman et al., 2000) and on tail flipping behavior in response to a stimulus. Animals that had the VNC transected were handled in the same manner, with the exception that there was one additional cut on the VNC. During this procedure the animals were held by hand, with their abdomen extended. The entire process from picking the animal up to placing it back into its container took less than 1 min. After cutting the VNC, or sham treatment, the animals were retested 1 week later, just prior to removing their chelipeds. After removing their chelipeds, the animals were retested within minutes for habituation to telson stimulation. They were then housed as isolates and retested after 1, 6 h and 3 days.
After the 3rd day of testing, the animals were dissected to view the sham and VNC transected site. The sham animals showed no apparent damage of the VNC: only scar tissue formed by a mass of connective tissues where the articulating membrane had been cut. The animals with a transected VNC showed a nerve cord that had separated substantially at the two cut ends. The rostal and caudal sections of the cut were pulled towards the remaining halves of the cords. Upon staining with 1% methylene blue, the cut ends could readily be observed. Staining with methylene blue was performed as described in Strawn et al. (2000). No physical connection existed between the two cut ends. Prior to dissection, a strong touch with a glass rod on their rostrum tested the sham and experimental animals for medial mediated tail flip induction. Only the sham animals exhibited a giant medial backward tail flip.
Testing Procedure 4
In a fourth series of experiments, 36 animals were divided into two groups of 18 to be treated as sham VNC or transected VNC. The earlier procedure of removing chelipeds, as outlined in experimental testing procedure 3, was repeated for both groups to enhance the animals’ ability to tail flip. The animals were housed for 3 weeks, then the sham VNC and transected VNC were further subdivided into three subgroups (sham injected, 5-HT injected, OA injected; n = 6 each). The animals were tested for tail flip habituation, then injected with either saline or 5-HT or OA, as in the systemic injections in testing procedure 1 described earlier. The animals were allowed to recover for 2 h, then were tested for tail flip habituation.
The statistical comparison of groups proved to be difficult because the rate of habituation varied in time, with some groups habituating in less than one minute and others taking several minutes and because there was individual variation within the groups. To facilitate comparison, the percentage of the group responding was graphed over time for the different paradigms. To control for repetitive testing over time with changing variables, parallel sham controls were run.
RESULTS
The procedure we used was to strike the telson with a glass rod, smoothed on the end (Fig. 1A), promotes the lateral giant mediated tail flip response; however, a non-giant mediated response might also have been activated. In some cases, the tail is tucked under the animal at the time of testing. In such instances a forceful tap was applied to the dorsal abdomen or to the side of the abdomen, as caudal as possible. In order to index the tail flip response, a score of 1 was given if the animal tail flipped. A score of 0 was given if the animal failed to tail flip or merely moved away (Fig. 1B). This index standardized the responses under the various experimental conditions and is comparable with the indices used in previously published reports (Kellie et al., 2001; Pagé and Cooper, 2004). In order to better describe the behavior of a group as a whole, the number of crayfish responding at a particular time was divided by the total number of crayfish tested in that group. This produced a percentage response for a group: The large crayfish (weight range 12-21 g in this study) tended not to tail flip as much as the smaller crayfish (Kellie et al., 2001).
We used the actions of the two biogenic amines, 5-HT and OA to examine the modulation of the sensory-motor reaction that evokes an escape behavior. The experiment was to monitor tail flip response by repeatedly tapping the telson of a freely behaving animal. The crayfish were tested for the rate of habituation (n = 24; Fig. 2A), then randomly divided into three groups (n = 8 in each group) and injected with either 5-HT, OA or treated as a sham (saline injection only). One hour after injection the animals were again tested. Only the group injected with 5-HT showed a pronounced resistance to habituation of the tail flip response (Fig. 2B). Even after 6 h, the 5-HT group was the only one that demonstrated a pronounced sensitivity to tail tapping (Fig. 2C). Possibly, being held as isolates produced the initial sensitivity in all three when tested 2 days later (Fig. 2D); however, the 5-HT group demonstrated a prolonged resistance to habituation, which was even greater than it had been 1 h after injection. A similar situation of a delayed super sensitivity with a reduced habituation rate to the tail taps was observed in animals that had their chelipeds removed (Kellie et al., 2001).
Since it is already established that cheliped removal produces an enhanced tendency for large crayfish to tail flip when they otherwise would not, we examined the effects of cheliped removal on the tail flipping responses of animals injected with the biogenic amines. The animals injected with 5-HT, OA, or treated as shams were examined just prior to cheliped removal and after cheliped removal, then 1 h after injections. All three groups showed an enhanced response immediately after the chelipeds were removed (Fig. 3A). The 5-HT group showed only a slightly enhanced responsiveness than did the sham group after the chelipeds were removed. The three groups were tested again 1 day (Fig. 3B) and 1 week (Fig. 3C) after the chelipeds were removed. All three groups showed a responsiveness upon initial testing 1 day after, with the 5-HT group still manifesting a slightly decreased rate of habituation. However, 1 week later there was only a slight difference for the 5-HT group among the three groups of injected animals that also had their chelipeds removed.
To determine whether direct descending neural input has an influence on the responsiveness of large animals such that they persist in showing a reduction in habituation to stimulation on the tail, the ventral nerve cord was severed between the thoracic and abdomen segments (A1 and A2). Sham operated animals served as controls for comparison. The animals were examined for habituation to the tail tap stimulus (Fig. 4A), then examined 1 week post-VNC transection but prior to the cheliped removal (Fig. 4B). The results indicate that the crayfish which exhibited a tail flip response had a reduced habituation rate in comparison with the VNC transected (Fig. 4B).
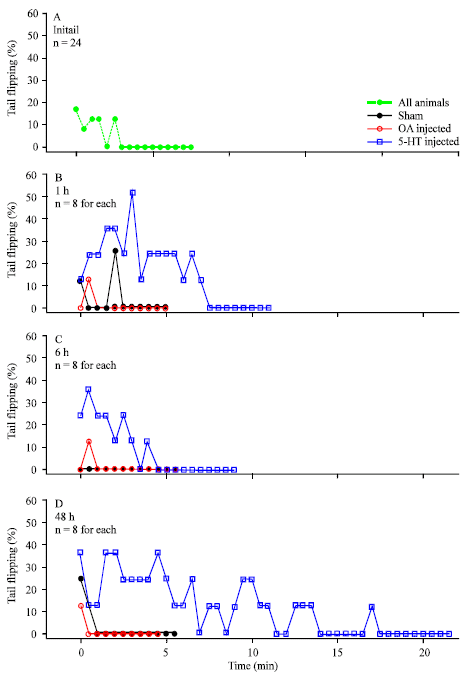 | |
| Fig. 2: | The acute effects of 5-HT and OA on the habituation of the tail flip response in representative animals are shown. Sham injections of saline were used as control for handling and the volume of injection into the hemolymph. A group of 24 animals was tested until each habituated to the tail taps (A), The amount injected should provide an approximately 10 μM circulating level of 5-HT or OA throughout the hemolymph. No significant effects were observed for OA (B), However, 5-HT increased the responsiveness or decreased the rate of habituation. To compare differences in experimental groups, a percentage of the total number of crayfish (n = 8 for each group) responding over time is plotted for sham controls, 5-HT and OA. Note that the acute effects of 5-HT decrease the rate of habituation, where as OA and sham groups show similar overall responses. The animals are were tested one hour (B), 6 h (C) and 48 h (D) after injection. Even after two days, the 5-HT injected animals showed a decrease in habituation |
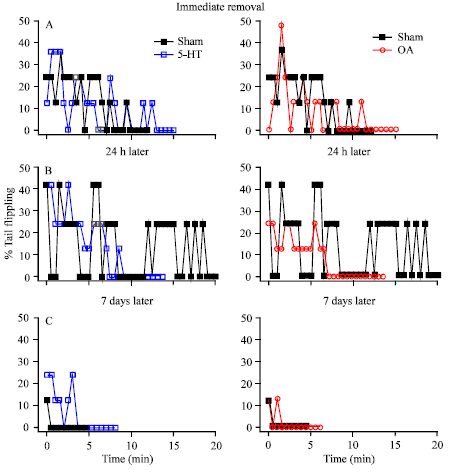 | |
| Fig. 3: | The effects of cheliped removal for animals exposed to exogenous biogenic amines are depicted. After the animals were injected with the 5-HT or OA, (A) an hour was allowed to pass before inducing the chelipeds to autotomize. The animals were then retested for the rate of habituation. This habituation assay was repeated 24 h later (B), as well as 7 days later (C). Animals exposed to 5-HT showed a decrease in the rate of habituation after 7 days after as compared to the OA injected group |
Two weeks after having either a sham experience or a VNC transection, the crayfish were tested prior to (Fig. 5A) and after having their chelipeds removed (Fig. 5B). The sham operated animals showed the most robust increase in tail flipping behavior. The transection of the VNC reduced the enhancing effect of removing the chelipeds. A low percent of the VNC transected animals demonstrated tail flipping behavior 1 h (Fig. 5C), 6 h (Fig. 5D) and 3 days (Fig. 5E) after the chelipeds were removed. Since each paradigm resulted in different rates of habituation for the sham, the overall rate of habituation should be examined. In some cases, it is obvious that a 100% of the experimental animals habituated before the sham controls.
Since it became apparent that transecting the VNC resulted in a reduced responsiveness to the enhanced tail flipping behavior when the chelipeds were removed, the experimental paradigm was reversed to first remove the chelipeds and then transect the VNC. This procedure would further demonstrate the effect of transecting the VNC on the tail flip response. Along with this procedure, the effect of OA and 5-HT was examined. Three days after the removal of the chelipeds, 18 animals were treated as VNC sham (Fig. 6A) or with a VNC transection (Fig. 6C) and tested for tail flip responsiveness. The VNC transected animals were markedly reduced in their responsiveness. Since the responses to neuromodulators could now be acutely tested independently of direct CNS innervation, the initial animals were subdivided into groups of 6 to examine the effect of OA or 5-HT treatment within the VNC sham and VNC transected groups.
 | |
| Fig. 4: | Severing the ventral nerve cord reduces the animals’ responsiveness to telson stimulation intended to induce tail flips. The rate of habituation was examined prior to transecting the nerve cord (A) and 1 week afterwards (B). Sham controls for the surgery were used for comparison. To compare differences in experimental groups, a percentage of the total number of crayfish (n = 8 for each group) responding over time is plotted |
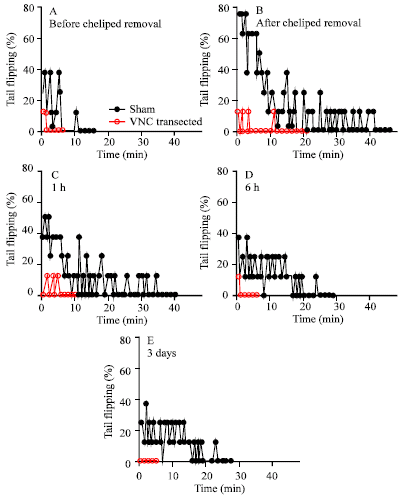 | |
| Fig. 5: | Animals without descending central commands showed a reduced tail flip response when the chelipeds were removed. The animals tested prior to (A) and after cheliped removal (B) showed the most pronounced effects. The enhanced effect of cheliped removal in prolonging the rate of habituation was observed only in animals with an intact VNC. This was examined immediately after cheliped removal, as well as 1 h (C), 6 h (D) and 3 days (E) later |
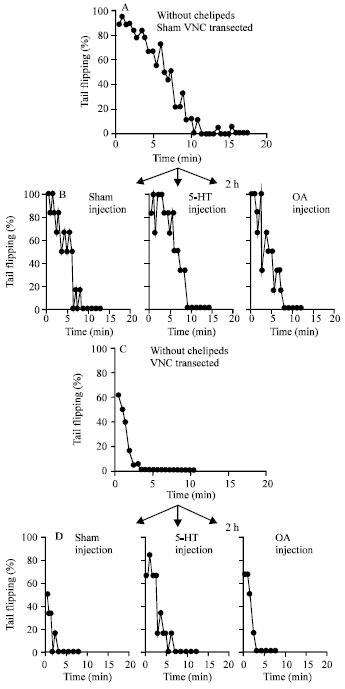 | |
| Fig. 6: | 5-HT and OA was exogenously applied to animals with autotomized chelipeds and transected VNCs to examine the direct effects of neuromodulators on the tail circuits of animals lacking direct central influences. The animals with autotomized cheliped were divided into two groups, one of which was treated with a VNC transection (A, n = 18) and the other used as a surgery sham control (C, n = 18). After recovering for 3 weeks from the sham or VNC transection, the animals were injected with 5-HT, OA or sham saline and left for 2 h before being tested for the tail flip habituation assay. In all cases, the VNC transected groups showed a reduced responsiveness to tail flips as compared to the VNC sham (A compared to C and injected groups (D compared to B). The 5-HT injected (VNC sham and VNC transected) did exhibit a slightly decreased rate of habituation compared to the OA and sham injected subgroups |
After the initial habituation trials subsequent to the VNC transection or sham handling, the animal were injected accordingly and examined 2 h later for their habituation rate. As compared to the sham VNC-treated animals, the ones injected with 5-HT (Fig. 6B) showed the greater resistance to habituation in comparison with the saline (Fig. 6B) and OA injected (Fig. 6B) animals. Among the VNC animals, 5HT injected animals demonstrated a reduced habituation (Fig. 6D) when compared with the OA and sham injected ones. Among the VNC sham animals, the VNC transected ones also demonstrated a reduced habituation when injected with 5-HT (Fig. 6D) when compared with the OA and sham injected ones with the VNC cut. The sham injected (Fig. 6D) and OA injected (Fig. 6D) animals in the VNC transected group were very prone to habituation in comparison with the sham VNC groups.
DISCUSSION
The main findings of this study are that systemic injections of 5-HT into a behaving crayfish results in an increase in responsiveness to a sensory stimulus as measured by a reduction in habituation of the tail flip assay. In addition, removal of the animal's chelipeds promotes a greater responsiveness to tail flipping. Combined cheliped autotomy and systemic injection produced no difference in animals with 5-HT, OA or saline injections. The transection of the VNC did result in the animals’ reduced response to tail flip stimulation and in an increased habituation rate. In animals with a transected VNC that were subsequently induced to autotomize their chelipeds there was a transitory (few hours’) enhancement of tail flip. However, the most striking findings are that animals with a transected VNC showed a marked enhanced response when injected with 5-HT. Together these findings demonstrate that the sensory-VNC-motor circuit for the tail flip can be modulated in the animal by 5-HT in the absence of higher command centers; however, with the intact CNS command centers, systemic injections of 5-HT promote the tail flip response and reduce the rate of habituation.
In isolated telson preparations, exogenously applied 5-HT increases sensory nerve transmission onto the lateral giant (Teshiba et al., 2001). However, the rate of 5-HT's application plays a role in the overall responsiveness (Teshiba et al., 2001; Krasne and Edwards, 2002). With the systemic injections the application would be similar to exchanging the bathing media rapidly for an isolated preparation. Such actions of 5-HT, it has been suggested, relate to the whole animal’s behavior and social status within crayfish hierarchies (Krasne and Edwards, 2002; Kravitz and Huber, 2003; Sneddon et al., 2000; Tierney and Mangiamele, 2001; Yeh et al., 1997). In our studies, all crayfish were housed as isolates in the preceding few weeks and during the experiments to control for the variable of social interactions and hierarchical status. It was shown also that novelty stress can alter the effects of 5-HT applied exogenously at the neuromuscular junctions of crayfish (Pagé and Cooper, 2004). The extent to which 5-HT and OA determines a crayfish's social status has not been delineated fully and is still an area of controversy (Doernberg et al., 2001; Kravitz and Huber, 2003; Listerman et al., 2000; McRae, 1996; Pagé and Cooper, 2004; Sneddon et al., 2000; Strawn et al., 2000). In spite of not fully knowing the role of 5-HT and OA in modulating crustacean social behavior, we have shown that the CNS does play a synergistic role for 5-HT and OA action. Thus, a behavioral state of the animal could well alter the circuit. Glanzman and Krasne (1983) were the first to show that 5-HT and octopamine have opposite modulatory effects on the crayfish's lateral giant escape reaction. Building on these past studies, several questions about intact animals remain to be answered: (1) What drives the release of these compounds? (2) What are the hemolymph concentrations in relation to behaviors? (3) What are the rates of release? (4) What is the duration of the cellular effects for the compounds? (5) What kinds of cross-interaction do the various neuromodulators have on the various target tissues? The more that is uncovered in physiology and pharmacology with reduced preparations, the sooner testing can occur of whole animals in a feasible, mechanistic manner.
Since the animals become more responsive to tail touches without their chelipeds suggests a neural and/or hormonal component on the tail sensory-motor circuit. To try to determine whether a descending neural path might be responsible, we transected the VNC in experimental paradigms before as well as after cheliped removal. We were surprised to observe the pronounced reduction in tail flip response, even before the chelipeds were removed, on transecting the VNC. The cause of this change is not known. It is probably due to nerve injury, since isolated preparations of the crayfish tail with fragments of the VNC stay functional for hours in minimal physiological saline. In some cases, after cutting the VNC the tail did flip once or twice, so it is not likely that a rapid sequence of electrical events caused the electrical synapses of the lateral giant axons to motor the nerves to depress. Also not likely is synaptic fatigue at the neuromuscular junctions, since little tail flipping activity was induced. A possibility, since we were working on a whole animal, is that the LG activity or inactivity after the transection might signal to the CNS that it is to release a hormonal factor to suppress the sensory tail circuit. Additionally, when stimulating the caudal aspect of the animal, we might have initiated a non-giant response from sensory structures on the cuticle of the animal, which are harder to control in an intact, freely behaving animal in the water than in an isolated preparation.
There are several studies which show inhibitory descending input depresses the lateral giant fibers (Krasne and Wine, 1975; Krasne and Lee, 1988; Vu and Krasne, 1992; Vu et al., 1993). However, cutting the inhibitory input was previously shown to have no role on habituation of the tail flip (Wine et al., 1975). But it was shown by Krasne and Teshiba (1995) that training altered the actions of the inhibitory paths, suggesting that the CNS does have a role in modulating this tail flip circuit.
The transected VNC resulting in a depressed responsiveness of the escape behavior suggesting that isolated tail preparations might be further removed in their physiological profiles and pharmacological responsiveness than they are in intact preparations. In the experimental paradigm with the chelipeds removed and an intact VNC (Fig. 3), the animals did not show any tail flipping behaviors that were different from those after the systemic injections with either 5-HT or OA. In fact, the injected animals with OA as well as 5-HT appeared to habituate even sooner than the sham injected animals. This was also an unexpected result. Perhaps the autotomization of the chelipeds was stressful and resulted in the releasing of various neuromodulators, such that any released neuromodulators masked the injected compounds.
As to the mechanism of a depressed tail flip response from transecting the VNC, only speculation remains. Possibly, putting the whole VNC and CNS in a dish, an intact telson and the recording of synaptic responses (induced by sensory neurons on the LG while transecting the VNC) could yield some understanding. This type of dissection might be too traumatic to the preparation, but it would allow one to examine whether there is a higher CNS component to the altered responses. Also, the exact cause of the increased tail flip response in whole animals when the chelipeds are autotomized remains to be determined. The results of our study suggest that hormones could be responsible, since even animals with a transected VNC increase in responsiveness to systemic injections of 5-HT (Fig. 6). The section of the ventral nerve cord may even result in changes in the excitability of the laterial giants. Previously, we suspected (Kellie et al., 2001) that OA might have been the biogenic amine to cause this altered responsiveness, since neurohumoral cells that project into the chelipeds contain OA and thus could release OA into the hemolymph with autotomy (Heinrich et al., 1999). However, there are many likely biogenic amines and peptides, not to mention the unexpected effects of cocktails of two or more substances (Djokaj et al., 2001). We feel that more detailed studies of the hormonal actions and combinations of hormones on crustacean neuronal circuits are required before speculating further on this matter.
ACKNOWLEDGMENTS
The support was provided by a G. Ribble Fellowship for undergraduate studies in the Department of Biology at the University of Kentucky (M-P.P.) and in part by NSF grants IBN-9808631 and ILI DUE-9850907 (R.L.C.). We thank Ms. Sophie Johnson (Australia) for editing.
REFERENCES
- Ayali, A. and R.M. Harris-Warrick, 1999. Monoamine control of the pacemaker kernel and cycle frequency in the lobster pyloric network. J. Neurosci., 19: 6712-6722.
Direct Link - Bustamante, J. and F.B. Krasne, 1991. Effects of octopamine on transmission at the first synapse of the crayfish lateral giant escape reaction pathway. J. Comp. Physiol. A, 169: 369-377.
Direct Link - Cooper, R.L., R.J. Chase and J. Tabor, 2001. Altered responsiveness to 5-HT at the crayfish neuromuscular junction due to chronic p-CPA and m-CPP treatment. Brain Res., 916: 143-151.
Direct Link - Djokaj, S., R.L. Cooper and W. Rathmayer, 2001. Effects of octopamine, serotonin and cocktails of the two modulators on synaptic transmission at crustacean neuromuscular junctions. J. Comp. Physiol. A, 187: 145-154.
Direct Link - Doernberg, S.B., S.I. Cromarty, R. Heinrich, B.S. Belt and E.A. Kravitz, 2001. Agonistic behavior in naive juvenile lobsters depleted of serotonin by 5,7?dihydroxytryptamine. J. Comp. Physiol. A, 187: 91-103.
Direct Link - Edwards, D.H., W.J. Heitler and F.B. Krasne, 1999. Fifty years of a command neuron: The neurobiology of escape behavior in the crayfish. Trends Neurosci., 22: 153-161.
Direct Link - Gleeson, R.A. and P.L. Zubkoff, 1977. The determination of hemolymph volume in the blue crab Callinectes sapidus, utilizing 14C-thiocyanate. Comp. Biochem. J. Neurosci., 6: 411-413.
CrossRefDirect Link - Herberholz, J., F.A. Issa and D.H. Edwards, 2001. Patterns of neural circuit activation and behavior during dominance hierarchy formation in freely behaving crayfish. J. Neurosci., 21: 2759-2767.
Direct Link - Horner, M., W.A. Weiger, D.H. Edwards and E.A. Kravitz, 1997. Excitation of identified serotonergic neurons by escape command neurons in lobsters. J. Exp. Biol., 200: 2017-2033.
Direct Link - Kellie, S., J. Greer and R.L. Cooper, 2001. Alterations in habituation of the tail flip response in epigean and troglobitic crayfish. J. Exp. Zool., 290: 163-176.
CrossRefDirect Link - Krasne, F.B., 1969. Excitation and habituation of the crayfish escape reflex: The depolarizing response in lateral giant fibres of the isolated abdomen. J. Exp. Biol., 50: 29-46.
Direct Link - Krasne, F.B. and T.M. Teshiba, 1995. Habituation of an invertebrate escape reflex due to modulation by higher centers rather than local events. Proc. Natl. Acad. Sci. USA., 92: 3362-3366.
Direct Link - Krasne, F.B., A. Shamsian and R. Kulkarni, 1997. Altered excitability of the crayfish lateral giant escape reflex during agonistic encounters. J. Neurosci., 17: 709-716.
Direct Link - Krasne, F.B. and D.H. Edwards, 2002. Modulation of the crayfish escape reflex-physiology and neuroethology. Integr. Comp. Biol., 42: 705-715.
Direct Link - Kravitz, E.A. and R. Huber, 2003. Aggression in invertebrates. Curr. Opin. Neurobiol., 13: 736-743.
Direct Link - Lang, F., C.K. Govind, W.J. Costello and S.I. Greene, 1977. Developmental neuroethology: Changes in escape and defense behavior during growth of the lobster. Science, 197: 682-685.
CrossRef - Listerman, L.R., J. Deskins, H. Bradacs and R.L. Cooper, 2000. Heart rate within male crayfish: Social interactions and effects of 5-HT. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 125: 251-263.
CrossRefPubMedDirect Link - Mesce, K.A., 2002. Metamodulation of the biogenic amines: Second-order modulation by steroid hormones and amine cocktails. Brain Behav. Evol., 60: 339-349.
Direct Link - Olsen, O.H., F. Nadim, A.A.V. Hill and D.H. Edwards, 1996. Uniform growth and neuronal integration. J. Neurophysiol., 76: 1850-1857.
Direct Link - Page, M.P. and R.L. Cooper, 2004. Novelty stress and reproductive state alters responsiveness to sensory stimuli and 5-HT neuromodulation. Comp. Biochem. Physiol. A, 139: 149-158.
Direct Link - Sneddon, L.U., A.C. Taylor, F.A. Huntingford and D.G. Watson, 2000. Agonistic behaviour and biogenic amines in shore crabs Carcinus maenas. J. Exp. Biol., 203: 537-545.
PubMedDirect Link - Strawn, J.R., W.S. Neckameyer and R.L. Cooper, 2000. The effects of 5-HT on sensory, central and motor neurons driving the abdominal superficial flexor muscles in the crayfish. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 127: 533-550.
PubMedDirect Link - Tabor, J.N. and R.L. Cooper, 2002. Physiologically identified 5-HT2-like receptors at the crayfish neuromuscular junction. Brain Res., 932: 91-98.
Direct Link - Teshiba, T., A. Shamsian, B. Yashar, S.R. Yeh, D.H. Edwards and F.B. Krasne, 2001. Dual and opposing modulatory effects of serotonin on crayfish lateral giant escape command neurons. J. Neurosci., 21: 4523-4529.
Direct Link - Tierney, A.J. and L.A. Mangiamele, 2001. Effects of serotonin and serotonin analogs on posture and agonistic behavior in crayfish. J. Comp. Physiol. A, 187: 757-767.
Direct Link - Vu, E.T. and F.B. Krasne, 1992. Evidence for a computational distinction between proximal and distal neuronal inhibition. Science, 255: 1710-1712.
Direct Link - Vu, E.T., S.C. Lee and F.B. Krasne, 1993. The mechanism of tonic inhibition of crayfish escape behavior: Distal inhibition and its functional significance. J. Neurosci., 13: 4379-4393.
Direct Link - Wilkens, J.L., A.J. Mercier and J. Evans, 1985. Cardiac and ventilatory responses to stress and to neurohormonal modulators by the shore crab Carcinus maenas. Comp. Biochem. Physiol. C, 82: 337-343.
CrossRef - Yeh, S.R., R.A. Fricke and D.H. Edwards, 1996. The effect of social experience on serotonergic modulation of the escape circuit of crayfish. Science, 271: 366-369.
Direct Link - Yeh, S.R., B.E. Musolf and D.H. Edwards, 1997. Neuronal adaptations to changes in the social dominance status of crayfish. J. Neurosci., 17: 697-708.
Direct Link