
O. A. Al-Dokhi
Department of Zoology, College of Science, King Saud University,
P.O. Box 2455, Riyadh� 11451, Saudi Arabia
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 1 | Page No.: 60-72







ABSTRACT
The aim of the present study was to describe the sperm head differentiation in the lizard Acanthodactylus boskinus at the ultrastructural level. For this purpose, five adult male lizards were collected during April and May, 2004 from the desert at the north-east area of Riyadh city, Saudi Arabia. Testes of the lizards were processed for transmission electron microscopy. Thorough observation of the ultrastructural differentiation events of the sperm head has led to a concept of four sequential morphogenetic phases; Golgi phase, cap phase, nuclear elongation and chromatin condensation phase and maturation phase. The Golgi phase is initiated by the proliferation of Golgi complexes in the early spermatids to generate numerous microvesicles and the phase is terminated by the formation of a voluminous acrosomal vesicle. Flattening of the developed acrosomal vesicle over the anterior nuclear portion to form a cap associated with dissolution of the acrosomal granule are the features of the cap phase. Subsequent elongation of the spermatid nucleus accompanied with nuclear chromatin condensation are the features of the following phase. The final maturation phase is characterized by complete chromatin condensation and disappearance of the microtubule manchette. The general features of the differentiation process, which accord with the common criteria of other reptiles, as well as the existing differences are discussed.
PDF Abstract XML References
How to cite this article
O. A. Al-Dokhi, 2006. Ultrastructure of Sperm Head Differentiation in the Lizard, Acanthodactylus boskinus (Squamata, Reptilia). International Journal of Zoological Research, 2: 60-72.
DOI: 10.3923/ijzr.2006.60.72
URL: https://scialert.net/abstract/?doi=ijzr.2006.60.72
DOI: 10.3923/ijzr.2006.60.72
URL: https://scialert.net/abstract/?doi=ijzr.2006.60.72
INTRODUCTION
The process of spermiogenesis involves biochemical, cytochemical and morphological changes which are necessary to transform spermatids into spermatozoa (Philips, 1974). Ultrastructural data on spermiogenesis of reptiles have improved the understanding of the adaptations accounting for fertilization and egg activation (Teixeria et al., 1999b). These data also provided an alternate source of characters for phylogenetic analysis (Jamieson et al., 1995, 1999; Tixira et al., 1999a).
Elucidation of the ultrastructure of spermiogenesis and spermatozoa in lizards has been the objective of some ultrastructural studies (Teixeria et al., 1999a, 1999b; Clark, 1967; Furieri, 1970, 1974; Del Xonte, 1976; Da Cruz-Landim and Dal Cruz-Hoffling, 1977; Butler and Gabri, 1984; Courtens and Depeiges, 1985; Al-Hajj et al., 1987; Dehlawi and Ismail, 1990, 1991; Dehlawi, 1992; Dehlawi et al.,1993; Ferreria and Dolder, 2002, 2003; Al-Dokhi, 2004b; Mubarak, 2004; Vieira et al., 2004). Only a reltively limited number of these studies have focused on sperm head differentiation in lizard species such as Tropidurus torquatus (dA cRUZ-LAMDIM AND dA cRUZ-hOFFLING, 1977), Podarcis tauria (Butler Gabri, 1984), Agama stellio (Al- Hajj et al., 1987), Agama blandfordi (Mubarak, 2004). However, the available references reveal some morphological variations in this process. Ultrastructure of spermiogenesis in the lizard Acanthodactylus boskinus has not been reported. Therefore, the current study is intended to provide ultrastructural data on the morphogenetic phases of sperm head differentiation in this species. Undoubtedly, these data would be beneficial to form a base for other studies concerned with phylogenesis in reptile species. The morphological comparison of sperm head differentiation in Acanthodactylus boskinus to other lizards or reptiles is also indicated.
MATERIALS AND METHODS
Five adult males of the lizard Acanthodactylus boskinus were collected during April and May, 2004 (period of sexual activity), from the desert (25°11’N, 46°51’ E), north-east of the city of Riyadh, Saudi Arabia. After decapitation, the lizards were dissected and their testes were removed and diced into small pieces that were immediately fixed by immersion in 3% buffered glutaraldehyde (0.1 M sodium cacodylate buffer at pH 7.2) for at least 4h at 41C. The fixed tissue specimens were thoroughly washed in the same buffer and then post-fixed in 1% osmium tetroxide (OsO4) in 0.1M sodium cacodylate buffer (pH 7.2) for 2 h. Subsequent dehydration of the fixed tissues was done in ascending grades of ethanol and the tissues specimens were then transferred to epoxy resin (Epon/Araldite mixture) via propylene oxide. Thin sections (70-80 nm) were cut with a diamond knife on an ultramicrotome (Leica, UCT), double stained with uranyl acetate and lead citrate and examined under a transmission electron microscope (JEOL, 100 CX) operating at 80 kV.
RESULTS
Newly formed spermatids occupy the apical compartment of the seminiferous tubules adjacent to their lumina (adluminal epithelium). Early spermatids are spherical in shape and their oval or round nuclei are centrally located and have evenly distributed fine granular euchromatin and occasional clumped heterochromatin. The well-developed juxtanuclear Golgi complex, which consists of flattened cisternae and many associated round vesicles, is the prominent organelle in the spermatid cytoplasm. Mitochondria, with linear cristae, manifested early aggregation on one side of the spermatid cytoplasm (Fig. 1). Occasional lipid droplets and scattered lysosomes are seen in the spermatid cytoplasm. Randomly distributed numerous free monoribosomes and detached polyribosomes are present throughout the cytoplasm. Cellular interconnections, represented as cytoplasmic bridges, apparently hold together and provide intercellular communication between early spermatids (Fig. 2). A thin layer of dense material lines the inner cytoplasmic sides of the intercellular bridges.
With commencement of spermatid differentiation, the Golgi phase (formation of acrosomal vesicle and granule) is evidenced by noticeable sequential morphological changes displayed by the Golgi complex (Fig. 3a-c). The compact parallel cisternae of the Golgi complex are increased in number and form whorls of concentrically laminated tubular structures. Many microvesicles arise from these proliferated cisternae. These microvesicles coalesce and as a result a large proacrosomal vesicle is developed in a close proximity to spermatid nucleus. Subsequently, the vesicle becomes lodged in the anterior portion of the spermatid nucleus. In this way, the acrosomal vesicle is formed and the site of contact between it and the nuclear envelope markes the future anterior pole of the spermatid nucleus. Possibly because of increasing pressure created by the expanding acrosomal vesicle, a cup-shaped proximal nuclear depression (anterior nuclear concavity) results which partially houses the vesicle. After this lodgment of the acrosomal vesicle, a single large electron-dense granule developed at its base. A close contact between the flattened cisternae of the Golgi complex and the apical membrane of the lodged acrosomal vesicle is still discernible.
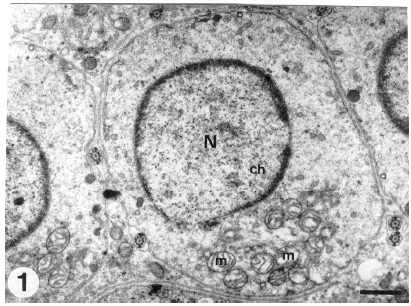 | |
| Fig. 1: | Transverse section through an early spermatid showing the centrally located round nucleus (N) which has evenly distributed fine chromatin (Ch). Note the round mitochondria (m), with linear cristae, which aggregated on one side of the spermatid cytoplasm. (Scale bar = 1 μm) |
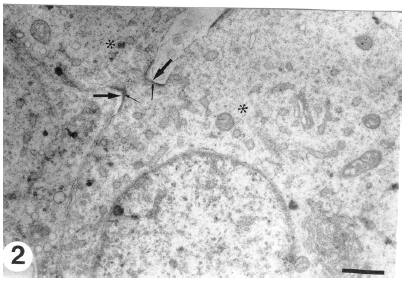 | |
| Fig. 2: | Cytoplasmic bridge (thick arrows) interconnecting the early spermatids (*). There is a dense material on the inner cytoplasmic sides of the intercellular bridges (thin arrows). (Scale bar = 1 μm ) |
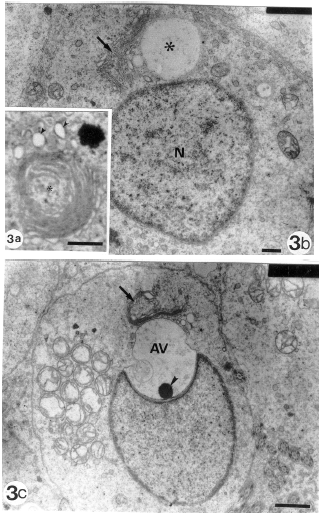 | ||
| Fig. | 3: | Sequential morphological changes of Golgi complex to form the acrosomal vesicle. |
| 3a: | Proliferated Golgi complex forming prominent whorls (*) of successive laminated tubular cisternae and associating microvesicles (arrowheads) (Scale bar = 1 μm) | |
| 3b: | Coalescence of the microvesicles to form a larger proacrosomal vesicle (*) which is still in contact with the proliferated cisternae of Golgi complex (arrow). Note that this vesicle is not yet lodged to the spermatid nucleus (N). (Scale bar = 0.2 μm) | |
| 3c: | Lodgment of the acrosomal vesicle (AV) into the proximal cup-shaped nuclear depression (anterior nuclear concavity). A single large electron-dense acrosomal granule ( arrowhead) is seen at the base of the acrosomal vesicle. Note that the Golgi complex (arrow) is still in a close association with the acrosomal vesicle. (Scale bar = 1 μm) | |
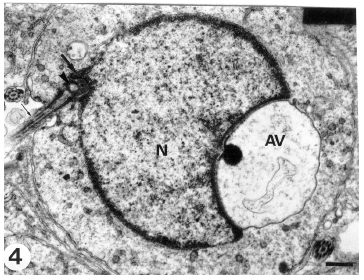 | |
| Fig. 4: | A section through an early spermatid showing the development a distal slightly arched nuclear depression (thick arrow) accomdating the proximal centriole (arrowhead). The acrosomal vesicle (AV) is still in contact with the spermatid nucleus (N). The distal centriole (thin arrow) is extended to form the microtubular axoneme. (Scale bar = 0.5 μm) |
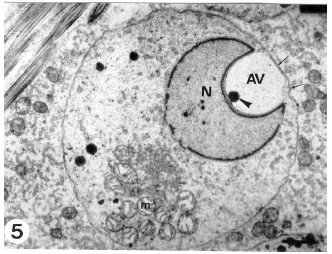 | |
| Fig. 5: | An early spermatid revealing a well-developed acrosomal vesicle (AV) and distinct acrosomal granule (arrowhead). The nucleus (N) and the attached acrosomal vesicle are displaced forward until the acrosomal vesicle membrane becomes apposed to the spermatid plasma membrane (arrows). Note the aggregated mitochondria (m) on one side o f the spermatid cytoplasm. (Scale bar = 1 μm) |
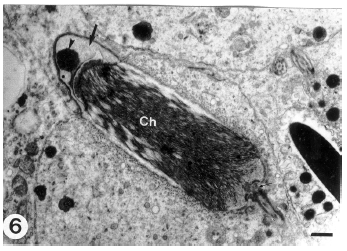 | |
| Fig. 6: | A longitudinal section through a middle stage spermatid. The acrosomal vesicle (thick arrow) is flattened over the nuclear sides. Acrosomal granule (arrowhead) is still prominent. Note the filamentous chromatin (Ch) and the nuclear fossa (thin arrow) which accommodates the proximal centriole. (Scale bar = 0.2 μm) |
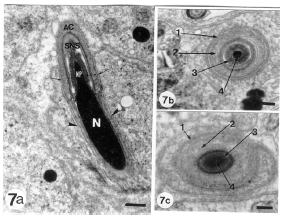 | |
| Fig.7a: | A longitudinal section through the head of a late formed spermatid. The elongated nucleus (N) appears with completely condensed chromatin and nuclear prolongation (NP). As a result of the nuclear compression, acrosomal vesicle is progressively flattened over the nuclear sides to form a cap (acrosomal cap) (AC) which covers the the proximal portion of the spermatid nucleus. The two lateral arms of the acrosomal cap (arrows) terminate at the nuclear shelves and rest on them. The Subacrosomal Nuclear Space (SNS) contains a striated material. The acrosomal granule has been dispersed in the acrosomal vesicle cavity. Note the longitudinal microtubular manchette (arrowheads) surrounding the nucleus in an anterior-posterior orientation parallel to the longitudinal axis of the nucleus. (Scale bar = 0.5 μm) |
| 7b and c: | Transverse sections through the nuclear rostrum and acrosomal complex. Nuclear rostrum is enclosed within successive membranes of the spermatid and acrosomal complex. Plasma membrane (1), acrosome (2), subacrosomal nuclear space (3), Nuclear rostrum (4). (b and c, scale bar = 0.2 μm) |
 | |
| Fig. 8: | Longitudinal (8a) and transverse (8b) sections through a completely condensed nucleus (N) of a late spermartid. Note the manchette microtubules (arrows) which appear in longitudinal (8a) and cross (8b) sections as a cylindrical structure enveloping and compressing the nucleus. (a and b, scale bar = 0.5 μm) |
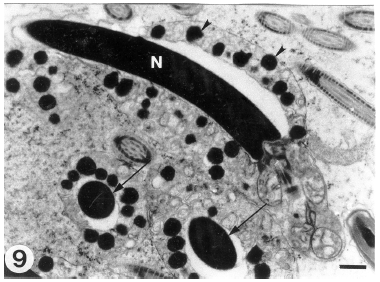 | |
| Fig. 9: | A longitudinal section through the head of a fully differentiated sperm. The elongated nucleus (N) has a completely condensed chromatin. The manchette microtubules have disappeared. Note the electron-dense bodies (arrowheads) encircling the sperm head. Arrows indicate transverse sections through the condensed nuclei which are encircled by the electron dense bodies. (Scale bar = 0.5 μm) |
The space between the nuclear envelope and vesicular membrane (subacrosomal space) shows the presence of a dense material. Occasionally, in some spermatids a distal slightly arched nuclear depression accommodating the proximal centriole appears while the acrosomal vesicle is still in contact with the spermatid nucleus (Fig. 4). As differentiation advances, the nucleus and the attached vesicle becomes dislocated forward until the acrosomal vesicle membrane is directly apposed to the spermatid plasma membrane (Fig. 5). This may represent the first morphological sign of the cap phase (formation of acrosomal cap). The Golgi complex is displaced gradually towards the posterior of the developing spermatid. Thereafter, sequential changes indicating nuclear elongation and chromatin condensation (nuclear elongation and chromatin condensation phase) proceeds and the elongated nuclei appear to push the acrosomal vesicle against the spermatid plasma membrane. As a consequence, the anterior nuclear pole attached to the acrosomal vesicle again becomes convex. As a result of the nuclear compression on the acrosomal vesicle, the latter is remodeled and progressively collapsed over the nuclear sides to form a cap (acrosomal cap) limited by the acrosomal membranes. The acrosomal cap covers a large proportion of the spermatid nucleus (Fig. 6) and its electron density increases.
Peripheralization of chromatin on the inner nuclear membrane is an early sign of chromatin condensation. The chromatin of the progressively prolongated nuclei undergoes a gradual condensation and is first organized as closely packed coarse filaments which form an interconnected meshwork (filamentous chromatin). The dense acrosomal granule disappears, probably owing to dispersion in content of the compressed acrosomal vesicle. The two lateral arms of the acrosomal cap terminate at the shelves of the condensed nucleus and rested on it. The highly condensed and elongated nuclei are initially embedded in the apical recesses of Sertoli cells and enclosed within successive membranes including those of Sertoli cell, spermatid and acrosomal vesicle (Fig. 7A-c). During the intermediate stage of differentiation, numerous microtubules (manchette) are closely associated with the nuclear envelope. The proximal end of the manchette microtubules was attached to the posterior end of the acrosomal vesicle lateral arms at the nuclear shelves. The elongated nuclei, with filamentous or condensed chromatin, are encircled by the longitudinally oriented microtubular manchettes (Fig. 8a and b) smanchette is a cylindrical structure possibly compresses the nuclei and having anterior-posterior orientation parallel to the longitudinal axis of the nucleus.
A noticeable depression (nuclear fossa) at the caudal pole of spermatid nucleus appears at a later stage of head differentiation and represents the implantation site for the neck region of the future tail. Thereafter, the maturation phase commences and the microtubular manchette disappears when the full differentiation of the sperm head region has been attained. The acrosome becomes elongated and cone-shaped and at the final stage a subacrosomal nuclear cap containing dense material is developed at the place of the pre-existing subacrosomal space. Thus, the nearly mature sperm has a highly condensed elongated cylindrical nucleus constituting the main portion of the head region and well-organised acrosomal and subacrosomal moieties capping the nucleus.
A prevalent ultrastructural feature in the head region of the released late spermatids is the existence of small electron-dense bodies encircling spermatid head (Fig. 9). Translocation of the late spermatid cytoplasm posteriorly constitutes the initial process of cytoplasmic shedding. The spermatid plasmalemma becomes apposed exactly to the contour of the fully differentiated spermatid components.
DISCUSSION
The presently studied Acanthodactylus boskinus reveal the general ultrastructural features of the sperm head differentiation which are in common with those described in other reptiles including lizard species (Courtens and Depeiges, 1985; Dehlawi and Ismail, 1991; Dehlawi, 1992; Dehlawi et al., 1993; Ferreira and Dolder, 2002, 2003; Al-Dokhi, 2004b; Mubarak, 2004; Charnier et al., 1967; Camps and Bargallo, 1977; Saita et al., 1987, Sprando and Russell, 1988; Healy and Jamieson, 1994; Ismail and Dehlawi, 1994; Ismail et al., 1995; Al-Dokhi, 1996, 2004a; Al-Dokhi and Al-Wasel, 2001a, 2001b, 2002). Early spermatids of Acanthodactylus boskinus contain aggregated mitochondria on one side of the cytoplasm before the commence of nuclear elongation. Some reptiles manifest aggregation of spermatid mitochondria (Ismail and Dehlawi, 1994; Ismail et al., 1995), while others reveal dispersion of mitochondria in their spermatid cytoplasm (Al-Dokhi and Al-Wasel, 2001a).
The early spermatids of Acanthodactylus boskinus are obviously interconnected by cytoplasmic bridges. These intercellular bridges are not a consistent feature of all lizards, since some species such as the lizard Stenodactylus selvini (Dehlawi and Ismail, 1991) shows no bridges between their early spermatids. However, these bridges were described in some other lizard species (Da Cruz-Landim and Da Cruz-Hoffling, 1997; Al-Hajj et al., 1987; Dehlawi, 1992; Mubarak, 2004; Dehlawi et al., 1990). Intercellular bridges were considered as an indication of incomplete meiotic divisions (Weiss and Greep, 1977; Fawcett, 1991). This incomplete meiosis results in formation of clusters of interconnected haploid spermatids (syncytial clones). Interchange of information from cell to cell (cell communication) takes place through these cytoplasmic bridges. These cellular connections together with some ectoplasmic specializations such as the dense plates between Sertoli cells contribute greatly to achievement of synchronous d1ifferentiation of spermatids (Junqueira and Carneiro, 1980; Hiatt et al., 1997).
Acrosomal vesicle is the prominent initial feature of the present differentiation process. The acrosomal vesicle develops in a close association to the proliferated Golgi complexes indicating its origin from this organelle. The currently illustrated acrosomal vesicle is partilly housed in an anterior nuclear cup-shaped depression which is considered a characteristic feature of spermiogenesis in Squamata (Clark, 1967; Butler and Gabri, 1984; Courtens and Depeiges, 1985; Dehlawi and Ismail, 1990, 1991; Dehlawi, 1992).
The early spermatid of Acanthodactylus boskinus has a single large acrosomal granule at the base of the acrosomal vesicle. The acrosomal granule is finally dissoluted to form the electron dense acrosomal medulla. The presence of a single acrosomal granule is a common feature in reptiles (Clark, 1967; Furieri, 1974; Del Conte, 1976; Butler and Gabri, 1984). However, some reptilian species such as Scincus mitranus (Al-Dokhi, 1996) and Agama adramitana (Dehlawi et al., 1992) reveal several acrosomal granules either attached or separated. In such species, the multiple acrosomal granules give rise to various parts of different electron density in the acrosomal medulla. The time of appearance of the acrosomal granule is also variable among lizard species. In Acanthodactylus boskinus, the early spermatid acrosomal granule becomes distinct after lodgment of the acrosomal vesicle on spermatid nucleus. This differentiation event is in accordance with that takes place in many other reptiles (Dehlawi and Ismail, 1990). In contrast, the lizard Stenodactylus selvini develops the acrosomal granule before this attachment (Dehlawi and Ismail, 19914). In the presently differentiated sperm head subacrosomal granule has not been noticed. Similarly, the lizard Agama agama (Chrnier et al., 1967) lacks a subacrosomal granule in its differentiating spermatids. Some reptiles shows the development of a subacrosomal material from the subacrosomal granule (Butler and Gabri, 1984; Dehlawi and Ismail, 1991; Capms and Bargallo, 1977). The final stages of sperm head differentiation in Acanthodactylus boskinus manifest the formation of a cup-shaped acrosome similar to that of other reptiles (Bao et al., 1991).
In accordance with the other reptilian groups, the nuclear differentiation described here includes nuclear elongation and condensation. The nuclear chromatin undergoes gradual condensation associated with increase in its electron density. Increased electron density of nuclear chromatin in the differentiating spermatids is associated with a high index of histonic proteins (Bao et al., 1991). Chromatin condensation followed the complex fibrous-lamellar pattern that is commonly manifested by squamates (Teixeira et al., 1999a; Clark, 1967; Al-Dokhi, 1996; Da Cruz-Hoffling and Da Cruz- Landim, 1978). The nuclear chromatin during spermiogenesis is changed into granular form which is built up of nucleohistones which is latter transformed into nucleoprotamine. Chromatin condensation may contribute in determining the nuclear shape through concentration of the nuclear histonic proteins and DNA (DNA-protein association) in a specific pattern. Also, chromatin condensation reduces nuclear volume to streamline the cell and, also it is crucial to preserve the genome (Fawcett, 1981, 1991).
The microtubular manchette surrounding the nucleus of growing spermatids is described as a helical system of microtubules (Bao et al., 1991). Longitudinally oriented manchettes surrounding the progressively elongated nuclei are recognizable in the developing spermatids of Acanthodactylus boskinus. In this respect, nuclear morphogenesis to acquire the peculiar nuclear shape may be the result of a sequence in progressive reorientation of the manchette from a circular to a longitudinal one (Fawcett, 1975, 1991). Nuclear shape is also determined by chromatin compaction in association with the role of manchette (Ferreira and Dolder, 2002). However, chromatin condensation may occur in absence of the microtubular manchette (Asa and Philips, 1988). Moreover, some investigators (Bao et al, 1991; Amaral et al., 1999) suppose the manchette is not an absolute requirement for nuclear morphogenesis and that the nuclear shape may be determined by the pattern of DNA-protein association. The arrangement of microtubules supports cellular and nuclear elongation during spermatid differentiation (Bao et al., 1991).
The electron-dense bodies encircling the head region are an outstanding feature in the fully differentiated spermatids of Acanthodactylus boskinus. The electron density of these structures suggests a lipoidal nature as supposed by (Hamilton and Fawcett, 1968) for similar structures. These structures were described as a tegosomial sheet in Python sebea (Boisson and Mattei, 1966), Lacerta vivipara (Courtens and Depeiges, 1985) and Chalcides ocellatus (Dehlawi, 1992).
In conclusion, the spermatids of Acanthodactylus boskinus undergo specific morphological modifications involving acrosomal complex formation, nuclear elongation and chromatin condensation. The sequence of events in sperm head differentiation described here for the lizard Acanthodactylus boskinus indicates the following sequential morphogenetic phases; Golgi phase (formation of acrosomal vesicle and granule), cap phase (formation of acrosomal cap), nuclear elongation and chromatin condensation phase and maturation phase (disappearance of manchette).
ACKNOWLEDGEMENT
This research was supported by the Research Center, College of Science, King Saud University, Saudi Arabia, project No. (Zoo/ 24-25/ 02).
REFERENCES
- Dokhi, O.A.A., 2004. Electron microscopic study of sperm head differentiation in the arabian horned viper Cerastes cerastes (squamata reptilia). J. Biol. Sci., 4: 111-116.
Direct Link - Asa, C.S. and D.M. Philips, 1988. Nuclear shaping in spermatids of the thai leaf frog, Megophrys Montana. Anatomical Record, 220: 287-290.
PubMed - Del Conte, E., 1976. The subacrosomal granule and its evolution during spermiogenesis in a lizard. Cell Tiss. Res., 171: 483-498.
CrossRef - Da Cruz-Landim C. and M. da Cruz-Hoffling, 1977. Electron microscope study of the lizard spermiogenesis in Tropidurus torquatus (Lacertilia). Caryologia, 30: 151-162.
Direct Link - Ferreira, A. and H. Dolder, 2002. Ultrastructural analysis of spermiogenesis in Iguana iguana (reptilia sauria iguanidae). Eur. J. Morphol., 40: 89-99.
Direct Link - Ferreira, A. and H. Dolder, 2003. Cytochemical study of spermiogenesis and mature spermatozoa in the Tropidurus itambere (reptilia squamata). Acta Histoch., 105: 339-352.
CrossRef - Hamilton, D.W. and D.W. Fawcett, 1968. Unusual features of the neck and middle piece of snake spermatozoa. J. Ultrastructure Res., 23: 81-97.
PubMed - Healy, J.M. and B.G.M. Jamieson, 1994. The ultrastructure of spermatogenesis and epididymal spermatozoa of the tutara Sphenodon punctatus (Sphenodontida amniota). Biol. Sci., 344: 187-199.
Direct Link - Jamieson, B.G.M., 1995. The Ultrastructure of Spermatozoa of the Squamata (Reptilia) with Phylogenetic Considerations. In: Advances in Spermatozoal Phylogeny and Taxonomy, Jamieson, B.G.M., J. Ausio and J.L. Justine (Eds.). Memooires du Museum National dHitoire Naturelle, Paris, France, pp: 359-383.
- Saita, A., M. Comazzi and A. Perrotta, 1987. Electron microscope study of spermiogenesis in Caiman crocodilus L. Boll. Zool., 4: 307-318.
Direct Link - Sprando, R.L. and L.D. Russell, 1988. Spermiogenesis in the red ear turtle (Pseudemys scripta) and domestic fowl (Gallus domesticus) a study of cytoplasmic events including cell volume changes and cytoplasmic elimination. J. Morphol., 198: 95-118.
PubMed - Vieira, G.H.C., G.R. Colli and N. Bao, 2004. The ultrastructure of the spermatozoon of the lizard Iguana iguana (reptilian Squamata iguanidae) and the variability of sperm morphology among iguanian lizards. J. Anatomy, 204: 451-464.
CrossRef