
H. Vazquez -lopez
Ecology Laboratory, Faculty of Superlores Iztacala Studies,
National Autonomous University of Mexico. Av. de Los Barrios: C. P. 54090,
Los Reyes Iztacala, Tlalnepantla, State of Mexico, Mexico
F. Alvarez
Laboratory of the National Collection of Crustaceans, Institute of Biology, University
National Autonomous State of Mexico, Circuito Exterior S/n, Ciudad Universitaria, Copilco,
Coyoac�n AP. 70-153 Mexico, Federal District. C.P. 04510
J. Franco
Laboratory of the National Collection of Crustaceans, Institute of Biology, University
National Autonomous State of Mexico, Circuito Exterior S/n, Ciudad Universitaria, Copilco,
Coyoac�n AP. 70-153 Mexico, Federal District. C.P. 04510
A. Moran
Laboratory of the National Collection of Crustaceans, Institute of Biology, University
National Autonomous State of Mexico, Circuito Exterior S/n, Ciudad Universitaria, Copilco,
Coyoac�n AP. 70-153 Mexico, Federal District. C.P. 04510
S. Chazaro
Laboratory of the National Collection of Crustaceans, Institute of Biology, University
National Autonomous State of Mexico, Circuito Exterior S/n, Ciudad Universitaria, Copilco,
Coyoac�n AP. 70-153 Mexico, Federal District. C.P. 04510
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 4 | Page No.: 344-353







ABSTRACT
One hundred and fourty nine crabs of the species Callinectes rathbunae Contreras parasitized with the rhizocephalan Loxothylacus texanus Boschma were captured. The average shell width was 8.48 cm. Three types of externa were registered in the hosts: virgin, immature and mature externa; their number varied from one to four, being more common the crabs with one externa. The behavior of parasitized crabs was registered during six months. The most aggressive crabs were those that presented virgin externa (n = 82), while the less aggressive ones were those who presented three to four mature externa (n = 3). The crabs with virgin and immature externa conserved the capacity of burying themselves, while the organisms with mature externa lost that capacity. The animals with mature externa passed most of the time grooming and oxygenating the externa they carried. The crabs with mature externa presented bigger voracity that the crabs with immature externa. It can be concluded that the level of aggressiveness of the parasitized crabs is related with the degree of development of the externa, the degree of aggressiveness of the parasitized crabs is not related with the sex and mature externa represent a great metabolic load for their hosts.
PDF Abstract XML References
How to cite this article
H. Vazquez -lopez, F. Alvarez, J. Franco, A. Moran and S. Chazaro, 2006. Observations on the Behavior of the Dark Crab Callinectes rathbunae Contreras Parasitized with the Rhizocephalan Loxothylacus texanus Boschma. International Journal of Zoological Research, 2: 344-353.
DOI: 10.3923/ijzr.2006.344.353
URL: https://scialert.net/abstract/?doi=ijzr.2006.344.353
DOI: 10.3923/ijzr.2006.344.353
URL: https://scialert.net/abstract/?doi=ijzr.2006.344.353
INTRODUCTION
Rhizocephala is a group of uniquely specialized crustacean that parasitize other crustaceans (Hoeg and Lutzen, 1995; Shukalyuk and Isaeva, 2000; Bresciani and Hoeg, 2001). This group includes Akentrogonida and Kentrogonida. All kentrogonida parasitize Decapoda. The families Peltogastridae and Lernaeodiscidae infest Anomura a few Thalassinidea and Caridea, meanwhile the Sacculinidae parasitize exclusively Brachyura. The Akentrogonida have a much wider taxonomic range of hosts comprising acorn barnacles, Stomatopoda, Cumacea, Isopoda and such Decapoda as Caridea, Thalassinidea, Brachyura and paguroid and galatheoid Anomura. For unknown reasons, however, no Rhizocephala parasitize any of the Decapoda Astacidea or Palinura (Hoeg and Lutzen, 1995). The adult parasite (Kentrogonida) consists of an internal system of roots called interna that connects with a external reproductive body called externa through a stalk (Hoeg, 1995; Hoeg and Lutzen, 1995; Gould, 1996; Bresciani and Hoeg, 2001). The adult parasites lack segmentation, mouth, appendages and no excretory or respiratory organs (Hoeg, 1992, 1995; Hoeg and Lutzen, 1995; Bresciani and Hoeg, 2001). The rhizocephalan root system is a unique structure almost without parallel in other parasitic structure (Bresciani and Hoeg, 2001). The rhizocephalans have nauplii and cypris larvae too, how others crustaceans (Glenner et al., 1989; Hoeg and Lutzen, 1995; Walker and Lester, 1998).
Rhizocephala affect the host’s development, transform the morphology, change secondary sex characters of the host too (Reinhard, 1950; Wardle and Tirpak, 1991; Hoeg and Lutzen, 1995) and manipulate host behavior (Hoeg and Lutzen, 1995; Innocenti et al., 1998, 2003). The objective of present work was to assess the behavior of crabs C. rathbunae Contreras parasitized with L. texanus Boschma in laboratory conditions. This resaerch was generated how part of a general study on life cycle characterization of L. texanus.
Materials and Methods
Dark crabs of the species Callinectes rathbunae Contreras (known in Mexico as jaiba prieta) parasitized with the rhizocephalan Loxothylacus texanus Boschma were captured in the lagoon-estuarine subsystem of Alvarado, Veracruz, Mexico, in regular samplings between 1999 and 2000. The organisms were captured using handmade fishing equipment which is locally called aro jaibero (crab hoop) (Lázaro-Chávez et al., 1996). The main objective of the captures was to obtain larvae of L. texanus to characterize the development of the larval stages. The physicochemical parameters were taken in the capture places. All the captured organisms were measured and transported to the laboratory of the National Collection of Crustaceans in the Universidad Nacional Autónoma de Mexico, in Mexico City. In the laboratory, the crabs were classified following the Wardle and Tirpak (1991); criteria it is necessary to mention that this criteria is based on the color of the externa present in each host. The crabs were lodged in recirculation systems for their acclimatization. The salinity was maintained in 15 ‰ and the temperature in 25°C. The animals were feeding every third day ad livitum with fresh fish and/or with chicken liver. The food leftovers were retired to maintain the quality of the water. One week later, crabs (females and males) with mature externa (clear brown and intense brown) were selected, which were lodged individually in aquaria of 20 L, maintaining the salinity in 15‰ and the temperature in 25°C (following the criteria of Yanagimachi, 1961b; Hawkes et al., 1985; Walter, 1988, 2001). As it was already mentioned, the main objective was to obtain larvae of L. texanus. The hosts were observed every day, keeping an eye on the eclosion of larvae or eggs of the externa. The behavior of 149 individuals, including hosts with virgin, immature and mature externa was registered. During six months, behavior of parasitized crabs was registered every day from 9:00 to 20:00. The frequency data were compared employed a Chi-square test (Sokal and Rohlf, 1981).
Results
In the collected crabs (149), were observed crabs with virgin externa (60 females and 33 males) (externa that have not been fecundated and presented a milky white color, were considered as immature), crabs with immature externa (25 females and 11 males) (which presented a yellow color and are about to expel larvae) and crabs with mature externa (9 females and 11 males) (from clear to dark brown color, which are the ones that expel larvae periodically). The hosts, both females and males, presented one to four externa (Table 1 and Fig. 2).
The width average of the crabs’ carapace was 8.48 cm. In Fig. 1 it can be appreciated the distribution of the crabs.
Behavior of the Crabs Before Being Separated Individually
In the laboratory the hosts were selected and separated, they were accommodated in systems of continuous recirculation, in plastic containers with a 10 L capacity and in similar systems in containers with a 6 L capacity, in batches of variable number.
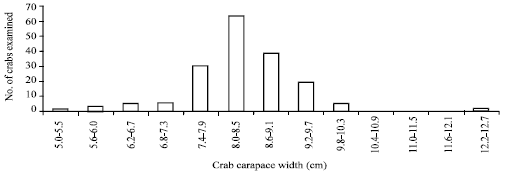 | |
| Fig. 1: | The width of the carapace of the parasitized crabs can be appreciated. The biggest frequency interval is the one ranging between 8.0-8.5 cm |
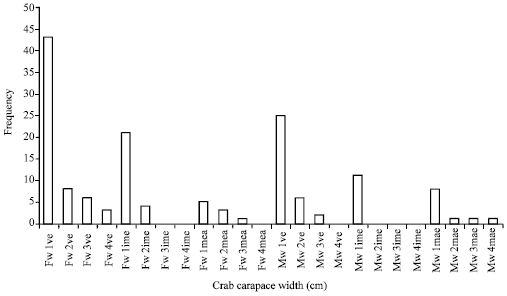 | |
| Fig. 2: | Number of externa, according to the host’s category. It can be appreciated that the more common number of externa it is one (Fw 1ve: Female crabs with 1 virgin externa; Fw1ime: female crabs with 1 immature externa, Mw 1ve: male crabs with 1 virgin and Mw 1ime: Male crabs with 1 immature externa) while the less common number is 4 (Fw 4ima: Female crabs with 4 immature externa, Mw 4ime: male crabs with 4 immature externa, Fw 4mae: Female crabs with 4 mature externa and Mw 4mae: Male crabs with 4 mature externa. The above mentioned is regarding the category of the number of externa) |
In the week of acclimatization, all individuals avoided contact with each other; nevertheless, they didn't show any sign of aggression. Each one of the organisms wanted to be isolated from the rest. When the food was provided, all individuals showed great anxiety, moving quickly their antennules and chelipeds; at this time, most of the crabs showed aggressiveness in different degrees. The animals that had a piece of chicken liver or a piece of fish devoured it quickly and defended themselves from the rest, escaping or using one of their chelipeds to attack the aggressor. During the experiment, the crabs were fed in different hours of the day. Most consumed more food when they were fed in the mornings or in the afternoons. When they were fed at noon-evening, the crabs left more food leftovers and they didn't show great activity; just a few seemed to be hungry all the time. This was observed only during a one week period because, after this, the organisms were selected and conditioned in an individual way.
| Table 1: | In the captured crabs, there were observed three types of externa: virgin, immature and mature externa |
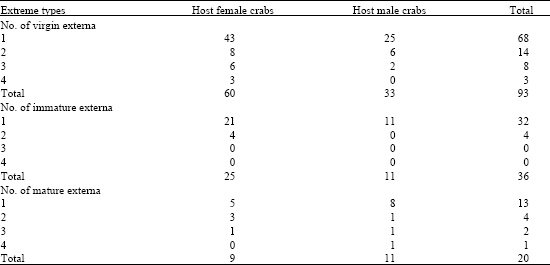 | |
Behavior of the Crabs after Being Selected
All the crabs were selected according to the number and type of externa that they presented. This way, they were grouped in twenty four categories (Table 1).
After the Crabs Were Separated, the Following Was Observed
Crabs with Virgin Externa
The animals, both females and males, with 1 and 2 virgins externa (82) (Table 1) presented the most aggressive behavior when someone came closer to the aquaria. The animals adopted a defensive position, rising and crackling the chelipeds, if any person brought near a hand or some object (a small aquarium net, a piece of food) to these animals, they attacked immediately and were calmed only when the person retired. This behavior appeared at any time. When food was provided, the crabs showed quick movements of the antennules and buccal appendixes and they went directly to the food. Regarding the grooming, these organisms constantly cleaned their eyestalks, using one of their chelipeds. Constantly, they opened the abdomen any time and introduced the dactyl of the 1st, 2nd or 3rd pereiopods and apparently, they cleaned the small externa. The behavior of this type of individuals is almost the same as that of the healthy crabs of the same species, maintained under similar conditions in the laboratory, which are extremely aggressive (personal observations). Just two crabs with an immature externa shed.
All crabs with virgin externa after a defensive attitude want to move away from the aggression, trying to hide in the corners of the aquaria, making movements as if they tried to be buried in the sediment. When the food was provided to them, they also presented quick movements of the antennules and the buccal appendixes.
All the crabs with virgin externa satisfied their appetite once a day; in several occasions, they were offered food hours after they had been fed and while some organisms just nibbled the food, others rejected it and some grabbed the food during half an hour and then let it go. The crabs with 3 and 4 virgin externa introduced some dactyl in the abdomen one or 2 times in the day.
Crabs with Immature Externa
A difference between crabs with virgin externa and immature externa is the speed of defense reaction, the movements were not so violent, but the use of the chelipeds continues being effective against other organisms (personal experiences). These crabs (36) making movements as if they tried to be buried in the sediment too but in minor grade. Another aspect observed was that female and malecrabs with immature externa (1-4) constantly lifted the last pair of pereiopods at the same time that they constantly moved the abdomen. Some time any crabs to make tried buried in the sediment.
Crabs with mature externa The crabs with mature externa (20) presented a completely different behavior. The females and males with 1 and 2 mature externa (17) were less aggressive than those already mentioned; while the less aggressive organisms in this category were the crabs (of both sexes) with 3 and 4 externa (3). When someone came closer to the aquaria that housed these organisms or they were brought near some object (an aquarium net, a piece of food), most responded presenting the chelipeds, but without moving from a place; after a few seconds, most of these crabs seemed to lose interest in the persons's presence or in the objects that he brought near them. In some cases, the researcher could approach a hand to two crabs with 4 externa, without receiving any damage at all (personal experiences). Unlike the animals with immature externa, the crabs with mature externa were more voracious, so they decided to feed them twice a day; so, when food was offered to them, they ate it. When food was offered to them, these crabs also presented movements of the antennules and the buccal appendixes, but these movements were less vigorous than those observed in crabs with immature externa. Of these, those that presented one externa, went direct to the food and consumed it, unlike the ones that presented 2, 3 and 4 externa. The crabs with 2 externa reacted to the food, but they slowly came closer to it. The crabs with three and four externa delayed more their arrival to the food, but the crabs with four externa gave the impression of not finding it visually, since they began an erratic search in the aquaria but, once they located it, they ate it quickly. All the organisms with mature externa (20) stayed in their place of preference during prolonged periods of time in one of the corners of their respective aquaria, lifting the back of their body. They just move the abdomen and pass some of the dactyls of the 1st, 2nd or 3rd pereiopods over the externa, as the healthy ovigerous females when they oxygenate and clean their ovigerous mass (Ritchie and Hoeg, 1981; personal observations). All the crabs with mature externa pass most of the time carrying out these activities, neglecting the cleaning of the carapace, so it could be observed that, in these crabs, a kind of sediment is accumulated, which was not observed in the organisms with immature externa.
Another important aspect is the fact that the seven crabs with 2, 3 and 4 mature externa, it was observed that the pair of swimming appendixes didn't present an apparent movement; in all cases it was observed that throughout the experiment, these organisms didn't move the swimming appendixes. For that reason, these animals (20) were taken to the surface of their aquaria and were loosed to observe their descent. In the 20 cases it was observed that the crabs touched the bottom without moving the swimming appendages, but moving the others. They pulled the swimming appendages of same organisms but when they were released, they bent again and settle over the carapace. Because of this, a possible paralysis was considered. These crabs introduced some of the dactyls of the 1st, 2nd or 3rd pereiopods over the externa many time along the day. Of these organisms, those that presented less activity during the hours of observation were those with 3 and 4 externa.
It was decided to compare the type of externa and the degree of aggressiveness of all organisms. The degree of aggressiveness was divided in three categories: passive, moderate and aggressive behavior, following the Innocenti et al. (1998) and Innocenti et al. (2003) criteria.
In this Case We Defined the Crabs Behaviour How
Aggressive: Are the crabs (both sexes) that attack quickly when close or approach any object to them, are very alert, they attempt buried in the sediment, they clean constantly their body surface and their behaviour is similar to healthy crabs.
Moderate: Intermediate behaviour between aggressive and passive, they clean any times in the day the externa and their body surface, are more relaxed that the previous.
Passive: Are the crabs more relaxed, non violent, they no attack, they no attempt buried in the sediment, they maintain elevated their body and fanning constantly the abdomen and the externa, occasionally they clean their body surface, are very relaxed.
To the Previous Data Was Apply a Chi-square Test (Table 2 and 3) Obtaining the Following Results
| • | Include all hosts: |
χ2O = 91.78, p>0.05
χ20.05,4 = 9.49
| • | Organisms with virgin externa: |
χ2O = 44.69, p>0.051
χ20.05,6 = 12.59
| • | Organisms with immature externa: |
χ2O = 98, p>0.05
χ20.05,6 = 12.59
| • | Organisms with mature externa: |
χ2O = 17.86, p>0.05
χ20.05,6 = 12.59
The Hypothesis Was the Following One:
Ho = The degree of aggressiveness of the parasitized crabs is not related with the degree of development of the externa
Ha = The degree of aggressiveness of the parasitized crabs is related with the degree of development of the externa
Rule of Decision
to reject Ho if χ20 > χ20.05, 4, χ20 > χ20.05, 6 and χ20 > χ20.05, 6
In all cases, Ho was rejected and this was the reason to conclude that the degree of aggressiveness of the crabs is related with the degree of development of the externa.
In all anterior cases were included females and males.
Was compared the aggressiveness level for sex vs number of externa (included virgin, immature and mature externa) too.
| • | Aggressiveness level of the females vs number of externa: |
χ2O = 4.06, p>0.05
χ20.05,6 = 12.59
| Table 2: | Frequencies of externa type and aggressiveness level of the crabs |
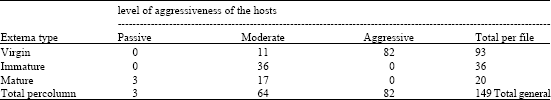 | |
| Table 3: | Observed frequencies are compared to the expected frequencies |
 | |
| • | Aggressiveness level of the males vs number of externa: |
χ2O = 6.01, p>0.05
χ20.05,6 = 12.59
The Hypothesis Was the Following One
Ho = the degree of aggressiveness of the parasitized crabs is not related with the sex.
Ha = the degree of aggressiveness of the parasitized crabs is related with the sex.
Rule of Decision
to reject Ho if χ20 > χ20.05, 6 and χ20 > χ20.05, 6
In both cases, Ho was accepted and this was the reason to conclude that the degree of aggressiveness of the crabs is not related with the sex.
DISCUSSION
The crabs that were captured had an average of carapace’s width of 8.48 cm because the opening of the mesh in the fishing equipment allows that small organisms can escape. This is easily verified because in samplings performed in the same zone of recollection, but using spoon nets of those which are employed for capturing juvenile shrimps, there have been captured organisms of the species C. rathbunae parasitized with L. texanus with sizes of 4 cm (personal observation). This is reinforced by Reinhard (1950) who argues that L. texanus parasitizes organisms of the species C. sapidus with a size interval of 38-85 mm; Lázaro - Chávez et al. (1996) who registered the presence of L. texanus in crabs of the species C. sapidus in the Tamiahua lagoon of Veracruz, Mexico, has registered parasitized crabs with sizes ranging from 26 to 169 mm for feminized organisms and from 45 to 115 mm for crabs with an externa (considering that the males with rounded abdomen and atrophied swimmerets are feminized).
Were observed three types of externa: virgin, immature and mature externa, being virgin the externa that can’t produce larvae yet, immature those that have been already fecundated and are close to produce larvae and mature externa those that have been already fecundated and produce larvae periodically. Wardle and Tirpak (1991) captured crabs of the species C. sapidus parasitized with L. texanus in the bay of Galveston, Texas and found female and male crabs with white externa considered immature and discovered immature eggs when examining internally. They also found brown and purple externa in which mature eggs were observed. Ritchie and Hoeg (1981) found that the mature externa of Lemaeodiscus porcellanae which infect porcelain crabs produce eggs periodically and that the immature externa are not sexually mature. Vázquez-Lspez et al. (2006) observed that the mature externa of Loxothylacus texanus parasitizing C. rathbunae Contreras, produce eggs constantly.
The number of immature and mature externa observed in the studied crabs was in the range of one to four. Wardle and Tirpak (1991) reported the same number of externa in C. sapidus. In a total of 285 parasitized organisms, 196 presented 1 externa, 76 2 externa, 12 presented 3 and 1 organism presented 4 externa. This distribution is similar to the find in the present study in which 113 crabs presented 1 externa, 2 with 22 externa, 10 had 3 and 4 crabs with 4 externa (included in each category: immature and mature externa). It is necessary to highlight that in any organism with 2, 3 and 4 externa, the presence of immature externa along with mature externa was observed. Hoeg and Lutzen (1995) mention that most of the rhizocephalans emerge like one external alone, which is connected to a radicular system and that multiple infections are rare; however, it is frequent to find two or three externa in L. panopaei and more than five in L. texanus. Galil and Lutzen (1998) mention that seasonal infections with Heterosaccus dollfusi are common in juvenile crabs of the species Charybdis longicollis Leene. And although there are no records in literature on the appearance of externa with a different degree of development in the same host, it can be speculated that the double, triple and quadruple externa (immature and mature) in a single crab, are the result of a multiple infection caused at the same time. Alvarez and Calderón (1996) suggest that L. texanus presents a contagious distribution; which would increase the probability that two or more cypris infect a crab at the same time.
Behavior Of the Crabs
The result obtained with the statistics (a) χ2O = 91.78, p>0.05, b) χ2O = 44.69, p>0.05, c) χ2O = 98, p>0.05 and d) χ2O = 17.86, p>0.05) regarding the aggressiveness of the animals it consistent with what has been observed in the laboratory, since the crabs with virgin externa are more aggressive than those that have immature and mature externa. This makes sense if we consider that the regeneration of the externa doesn't exist or, at least, this phenomenon has not been observed in L. texanus and other species like L. panopaei and Sacculina benedeni. An aggressive behavior of the host could result in the loss of the externa, with which the probability of reproduction of the parasite would be of 0%. In this respect it has been observed that the rhizocephalans take absolute control of their host since, among other things, they modify the behavior and the morphology of their hosts (in both sexes) (Reinhard, 1950; Ritchie and Hoeg, 1981; Wardle and Tirpak, 1991; Hoeg, 1995; Hoeg and Lutzen, 1995; Galil and Lutzen, 1998; Innocenti et al., 1998, 2003). Innocenti et al. (2003) observed that the crabs parasitized with H. dollfusi escape from the aggressions of other crabs, possibly increasing the expectation of its own life and the one of the parasite.
It was concluded that the degree of development of the externa related to the degree of aggressiveness of the crabs. Gherardi and Cassidy (1995) mention that a possible effect on the behavior of the hermit crabs Discorsopagurus schmitti parasitized with Peltogaster boschmae and Thilacoplethus (= Thompsonia) reinhardi, be the presence of lethargy, which could reduce the ability of the crabs to find an empty refuge. Wardle and Tirpak (1991) establish that the crabs of the species C. sapidus Rathbunae with mature externa were less active and therefore less aggressive that the crabs of the same species with immature externa. Innocenti et al. (2003) conducted several experiments to compare the behavior patterns among healthy organisms of the species C. longicollis Leene parasitized with the rhizocephalan H. dollfusi Boschma. They observed that both the healthy females and males differ in the degree of aggressiveness, while their homologous parasitized presents a similar pattern of behavior regarding the aggressiveness; they mention that the healthy organisms are more aggressive than the parasitized ones.
Regarding the alimentary habits, Wardle and Tirpak (1991) observed that the parasitized crabs are less aggressive and less active when food is present. Innocenti et al. (2003) concluded that the presence of food does not alter significantly the behavior of the parasitized animals. However, there are no papers regarding the alimentary behavior of crabs parasitized with rhizocephalans. Innocenti et al. (1998) carried out observations on the behavior of the portunid crab C. longicollis Leene parasitized with the rhizocephalan H. dollfusi Boschma. They took into account the behavior regarding excavation movements and burial, the cleaning of the externa and the crab’s locomotion and shedding among other aspects, although they mentioned that they didn't consider the movements of the antennas, antennules and maxilipeds.
It is mentioned that the crabs Carcinus maenas parasitized with the rhizocephalan Sacculina carcini constantly elevate the last pair of pereiopods and extend the abdomen (Hoeg and Lutzen, 1995). With this action, they allow that the male cypris larvae may locate the externa they carry and, therefore, fecundate it. It has been observed that the crabs with immature externa have not totally lost the ability to be buried in the silt and not the crabs with mature externa, which lose the capacity to be buried and, on the other hand, it has been observed that the braquiurus crabs constantly ventilate and groom the mature externa they carry, moving the abdomen more quickly that the crabs with immature externa. The behavior of lifting the rear part of the body has the purpose of maintaining the externa’s opening far from the silt, which facilitates the expulsion of the larvae. This would be affected if the crabs didn't lose their habit of burying themselves. It is mentioned that the behavior of flapping the abdomen is common in healthy ovigerous females (Ritchie and Hoeg, 1981; Wardle and Tirpak, 1991; Hoeg, 1995; Hoeg and Lutzen, 1995; Galil and Lutzen, 1998; Innocenti et al., 1998, 2003).
On the other hand, Hoeg and Lutzen (1995), Galil and Lutzen (1998), Innocenti et al. (1998 and 2003) affirm that the parasitized crabs, both females and males, expend a lot of time cleaning and ventilating the mature externa. Ritchie and Hoeg (1981) state that if the cleaning appendixes cleaners are removed from a parasitized crab, the externa soon becomes fouled and necrotic.
The above-mentioned can explain what was observed in this study. With this, it can be affirmed that: A) the crabs of the species C. rathbunae contreras with mature externa are less aggressive than those that present virgin and immature externa, since the parasite, when controlling the behavior of his host, channels all its energy to satisfy its alimentary and reproductive requirements and cannot expose its externa to a possible loss because it cannot be regenerated. B) the crabs with virgin and immature externa have not lost their ability to bury themselves; but not the crabs with mature externa. C) the crabs clean and oxygenate the mature externa, the same as other parasitized crabs, D) the crabs with mature externa increase their appetite because the parasite represents a great metabolic load, E) the degree of aggressiveness of the parasitized crabs is related with the degree of development of the externa and finally, F) the degree of aggressiveness of the parasitized crabs is not related with the sex.
Analyzing the existent literature on the effects that cause the rhizocephalans on their hosts, is possible that the apparent paralysis observed in the crabs with mature externa could be another mechanism to avoid that the crabs consume energy in the swimming displacement, since the healthy organisms of the same species can displace themselves swimming swiftly from one place to another (personal observations). Perhaps a crab spends more energy swimming than walking.
REFERENCES
- Bresciani, J. and J.T. Hoeg, 2001. Comparative ultrastructure of the root systemin rhizocephalan barnacles (Crustacea: Cirripedia: Rhizocephala). J. Morphol., 249: 9-42.
Direct Link - Galil, B.S. and J. Lutzen, 1998. Jeopardy: Host and parasite lessepsian migrants from the mediterranean coast of Israel. J. Natural History, 32: 1549-1551.
Direct Link - Gherardi, F. and P.M. Cassidi, 1995. Life history patterns of Discorsopagurus schmitti, a hermit crab inhabiting polychaete tubes. Biol. Bull., 188: 68-77.
Direct Link - Hoeg, J.T., 1995. The Biology and Life Cycle of the Rhizocephala (Cirripedia). J. Marine Biol. Assoc. UK., 75: 517-550.
Direct Link - Innocenti, G., N. Pinter and B.S. Galil, 2003. Observations on the agonistic behavior of the swimming crab Charybdis longicollis leene infected by the rhizocephalan barnacle Heterosaccus dollfusi boschma. Can. J. Zool., 81: 173-176.
Direct Link - Shukalyuk, A.I. and V.V. Isaeva, 2000. Intravital and histological studies of the internal organization of the rhizocephalan barnacle Sacculina polygenea. Russ. J. Marine Biol., 26: 209-211.
CrossRefDirect Link - Walker, G., 2001. Introduction to the rhizocephala (Crustacea: Cirripedia). J. Morphol., 249: 1-8.
Direct Link - Yanagimachi, R., 1961. The life-cycle of Peltogasterella (Cirripedia, Rhizocephala). Crustaceana, 12: 183-186.
Direct Link