
Z. P. Shuranova
Institute of Higher Nervouse Activity and neurophysiology, University of Kentucky, Lexington, KY
Y. M. Burmistrov
Institute of Information transmission Problem, Russian Acadmey of Scinces, Moscow, Russia
J. R. Strawn
Institute of Higher Nervouse Activity and neurophysiology, University of Kentucky, Lexington, KY
R. L. Cooper
Department of Biology, University of Kentucky, Lexington kY
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 242-283







ABSTRACT
In vertebrates, the regulation of internal organs is primarily driven by the autonomic nervous system. In crustaceans, nervous control of organs has been investigated elaborately but primarily on the isolated organs. The increased interest in examining the whole system in crustaceans, especially in freely behaving animals, indicates that an ample and sophisticated nervous control of these internal organs is typical for these higher invertebrates. To prepare for rapid escape or confrontation, or for maintenance of a resting state during a relaxed period, these animals appear to need certain neural structures which can be compared to the autonomic nervous system of vertebrates. This review is to examine evidence for an analog to the autonomic nervous system in the crayfish and other decapod crustaceans. It was concluded that, notwithstanding obvious structural differences, some functional similarities take place in both groups of animals. The common features include the existence of its own nerve nets in the heart and digestive system as well as an inhibitory or acceleratory extrinsic control of their functions by specialized neurotransmitters. Additionally, the release of neuromodulators and neurohormones into the circulation appears to be widespread and extremely important for control of the vegetative organs in decapods as in vertebrates.
PDF Abstract XML References
How to cite this article
Z. P. Shuranova, Y. M. Burmistrov, J. R. Strawn and R. L. Cooper, 2006. Evidence for an Autonomic Nervous System in Decapod Crustaceans. International Journal of Zoological Research, 2: 242-283.
DOI: 10.3923/ijzr.2006.242.283
URL: https://scialert.net/abstract/?doi=ijzr.2006.242.283
DOI: 10.3923/ijzr.2006.242.283
URL: https://scialert.net/abstract/?doi=ijzr.2006.242.283
INTRODUCTION
The Autonomic Nervous System in Vertebrates
A steadily growing number of reports show that in decapods various behavioral acts are coupled with specific reactions of the internal organs. First, the cardio-vascular system is tightly connected with the ventilatory system. On the other hand, functioning of the internal organs strongly depend on the circumstances and current state of the animal. The mounting evidence impelled us to assume that the normal vital activity of decapods requires a well-integrated nervous system to control not only somatic but also visceral end organs. This system parallels the autonomic nervous system in vertebrates in many ways.
The Autonomic Nervous System (ANS) is a complex of central and peripheral nerve cells that are associated with a number of internal body functions. In vertebrates, the ANS has three subdivisions each of them with a distinctive anatomical organization: sympathetic, parasympathetic and “metasympathetic” (Nozdrachev, 1983) commonly named enteric. Together, they innervate the blood vessels, heart, smooth muscles, viscera and glands.
A distinctive feature of the sympathetic and parasympathetic systems, in comparison to the cerebrospinal somatic nervous system, is that their innervation of the effector organs is not direct: there are preganglionic neurons located inside the CNS and postganglionic neurons whose processes innervate the effector structures. Preganglionic neurons of both sympathetic and parasympathetic divisions are known to be cholinergic. Postganglionic parasympathetic also release acetylcholine, whereas sympathetic postganglionic neurons release noradrenaline. There are some exceptions to this rule and in addition, transmitter specificity of the postganglionic neuron may depend on the target and the developmental stage. Both of these subdivisions of the ANS are quite different. The parasympathetic system is made up of relatively independent components, so its action is rather restricted. On the contrary, the sympathetic, or, more correctly, adreno-sympathetic system, appears to be tied together and functions almost as a unit. Earlier, it was suggested that the sympathetic and parasympathetic systems have different functions and act mostly as antagonists. It is clear recently that (1) both divisions operate in conjunction with each other (“accentuated antagonism”) and (2) with the somatic motor system to regulate most behaviors, maintaining an internal stable environment in the face of changing external conditions. It should be emphasized that the main effect of both ANS subdivisions is not triggering but modulating the own activity of visceral muscles by releasing a lot of neuroactive substances (biogenic amines, such as serotonin, dopamine etc. and neuropeptides, such as substance P, calcitonin-gene-related peptide, vasoactive intestinal peptide, etc.) (Nozdrachev, 1996).
The third subdivision of the ANS, often described as “intrinsic” or “enteric”, because it was studied mainly in the digestive tract, is also termed “metasympathetic” (Nozdrachev, 1983). It includes neuronal structures located inside different internal organs which are predominantly involved in the functioning of these organs. The sympathetic and parasympathetic systems exert their influence on the vital functions of the body through the intrinsic neural elements located in the internal organs.
The higher autonomic center within the vertebrate’s brain is the hypothalamus which controls the main vital functions of the organism (Iversen et al., 2000). The hypothalamic nuclei are neither sympathetic or parasympathetic-they control functional activity of both parts of the ANS. Moreover, many hypothalamic cells are neurosecretory; they can influence the internal organs directly and/or through the neuroendocrine cells situated in the hypophysis. The hypothalamus integrates behavioral and emotional responses arising from the forebrain, with ongoing metabolic need to produce highly coordinated control of behavior.
Historical Notes about the ANS in Arthropods
Clearly the higher invertebrates, arthropods, have an ample innervation of the internal organs. The question, however, remains, at what level are the above-mentioned features of the vertebrate ANS applicable to these invertebrates.
At the late 18th-early 19th centuries, the authors studied the innervation of visceral muscles in arthropods did not hesitate to term the revealed nervous structures as ‘sympathetic’ (Lemoine, 1868; Alexandrowicz, 1909 ); it may be noted, however, that their meaning of the ‘sympathetic’ nervous system probably differed strongly from the recent one. Special comparison between the ANS in vertebrates and arthropods was made by the Russian comparative neuroanatomists (Zavarzin, 1941; Orlov, 1926, 1927, 1929; Nevmyvaka, 1928).
Zavarzin (1941) described three subdivisions in the ANS of insects. He believed that its cranial part was represented by the stomatogastric system of the recurrent nerve consisting of the frontal ganglion (connected with the cerebral ganglia), the recurrent nerve and the oesophageal ganglion innervating the heart and colon (he analogized this with the anterior part of the parasympathetic system of vertebrates). In his opinion, there was a topographical correspondence between the sympathetic nervous system of vertebrates and the system of so-called unpaired nerves of insects (segmentary nerves that exit from the ventral cord ganglia and supply many internal organs). Zavarzin (1941) compared the unpaired nerve of the last abdominal ganglion innervating the hindgut and sex organs of insects with the caudal division of the vertebrate parasympathetic system. According to more recent data, the autonomic centers of higher order are located in the tritocerebrum in insects (Gundel and Penzlin, 1978; Plotnikova, 1979).
Zavarzin (1941) suggested that there was a principal similarity between the two main divisions of the CNS supplying the external (somatic) and internal (visceral) muscles. He also believed that the both subdivisions of the CNS developed together with their effectors; thus vertebrates, with their highly developed visceral muscles possess the highly differentiated ANS, whereas the ANS in invertebrates is differentiated in lesser degree. At the same time, he suggested that differences between the ANS in vertebrates and arthropods are minor. In his opinion, the main difference was in lack of the twofold (sympathetic and parasympathetic) innervation of the internal organs in arthropods. Zavarzin (1941) also thought that arthropods do not possess internally secreting organs (“internal glands”), which were considered very important for functioning of the ANS in vertebrates; this obviously was his mistake probably caused by the deficit of experimental data at that time.
There is no possibility to consider the problem of the putative ANS in insects in this review. We can refer, however, to some very interesting reviews concerning the control of circulation (Miller, 1997), the development of enteric nervous system in insects (Copenhaver and Taghert, 1989, 1991; Ganfornina et al., 1996; Hartenstein, 1997) and structure and function of the stomatogastric nervous system (Penzlin, 1985). Miller’s conclusion is that “the coordination between circulation and ventilation is so highly organized that an overall autonomic nervous system is proposed or suggested to exert hierarchical control over thermoregulation, respiration and circulation. This hypothetical system, the autonomic nervous system, will have to be described before the nervous control of the circulatory system can be fully understood”. As for the enteric nervous system that innervates the alimentary tract and other related structures it is composed of a group of interconnected ganglia on the surface of the foregut and a network of neurons organized as a plexus on the surface of the foregut and the midgut. In spite of the existence of several inputs from the CNS, the enteric nervous system is largely autonomous in its functions concerning the control of feeding, peristaltic movements of the gut, molting and metabolism (Penzlin, 1985). The embryonic development of this system was shown to present some parallels to the development of the vertebrate peripheral nervous system (Ganfornina et al., 1996; Hartenstein, 1997).
One of the first (or probably the first) paper (titled: “Zur Kenntnis des sympatischen Nervensystems der Crustaceen”) by the famous neuroanatomist, J.S. Alexandrowics, was devoted to the study of ’fine histological interrelations’ in the hindgut of several crustaceans, mostly, of the crayfish (Alexandrowics, 1909). Based on the histological and physiological evidence, he concluded that the macruran hindgut has its own nervous system capable to perform peristaltic movements after complete isolation from the ventral nerve cord. The nerve fibers entering the hindgut from the last abdominal ganglion, on his opinion, serve to regulate the “automatic” movements of its muscles.
Later, several anatomical works on the nervous control of the digestive system of the crayfish were performed by Orlov (1927, 1929). An absence of sensory nerve endings in the Stomatogastric Ganglion (STG) indicated, in his opinion, that this ganglion cannot work by itself, without the connection with the commissural ganglia. Thus Orlov (1927, 1929) treated the neurons of commissural ganglia as pre-synaptic and neurons of the STG, as post-synaptic. He suggested that there is principal similarity in organization of nervous control of the digestive system between not only crustaceans and insects but also between the arthropods and vertebrates (Fig. 1).
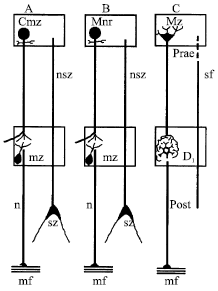 | |
| Fig. 1: | Scheme of neuronal interrelations in the stomatogastric system of the crayfish (A), beetle (B) and in vertebrate ANS (C) (Orlov, 1927) |
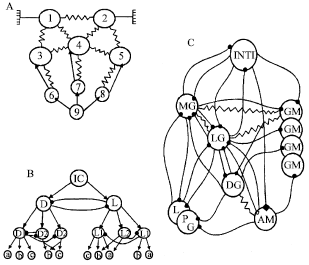 | |
| Fig. 2: | Scheme of central pattern generators for heart rhythm (A), ventilatory pattern (B) and gastric rhythm of the stomach (C) (modified from Taylor, 1982 (A, B) and Selverston, 1987 (C) Connections between neurons: filled circles, chemical excitatory; arrows (A, B) and filled squares (C), chemical excitatory; resistor symbol, electrotonic coupling; open square, functional excitation |
At the end of his study devoted to the STG of the crayfish he wrote: “Es muß.... konstatiert werden, daß die... Untersuchung der histologischen Struktur des Magenganglions des Astacus meine Voraussetzung... von der Möglichkeit eines vergleichend-histologischen Studiums des sympatischen Nervensystems der Wirbeltiere und Wirbellosen in vollem Masse bestätigt...” (Orlov, 1927, 1993).
In contrast to insects and in spite to the great advantages in many areas related to the crustacean neurophysiology, the problem of the specialized nervous control of the internal organs in crustaceans has not been investigated during the last 50-60 years of the 20th century. Probably the last paper where the term “sympathetic” as related to the crustaceans was used is that by Chaudonneret (1956) who described in Cambarus the “chaine sympatique laterale” that goes along the ganglion (SOG) and makes contacts with the nerve elements of different neuromeres. Later, even the analogies in nervous control of the crustacean visceral systems with that of the vertebrates were rigorously criticized (Selverston and Mulloney, 1974) and only some Russian neuroanatomists maintained Zavarzin’s views (Fomichev, 1986).
The situation is better with the cardiovascular system and the closely related system-the ventilatory system. Here one may refer to the reviews by Taylor (1982), McMahon (1995, 2001) and Wilkens (1999). As McMahon concluded, “Examination of responses to activity and environmental disturbance reveal complex responses in both respiratory and cardiovascular systems which are integrated into an overall compensatory response... The system revealed is functionally equivalent to the autonomic control observed in the vertebrates but differs markedly in anatomical and pharmacological detail” (McMahon, 1995).
The aim of this review is to consider the anatomical and physiological data on the nervous control of the main internal organs in decapods, in order to draw attention to an obvious but often omitted fact that normal adaptive behavior of these higher invertebrates is impossible without the elaborated nervous control of their internal organs adjusted to the internal demands of the animal and coordinated with current environmental conditions.
Direct Neural Control of Main Internal Organs in Crustaceans
Cardio-vascular System
In decapods, the heart is a single-chambered organ, which is suspended in the pericardial cavity by the elastic ligaments. In the dorsal wall of the heart is a ganglion (Alexandrowicz, 1932) composed of a small number of neurons varying from 5 (in the shrimp Alpheus) up to 16 (in the crayfish Procambarus) (Kuramoto and Yamagishi, 1990). The physiological and morphological properties of this ganglion have been relatively well studied, especially since 1951 when J. Welsh introduced it as an isolated preparation that exhibits remarkable reliability and stability in generating a rhythmic pattern (Cooke, 1988) (Fig. 2A). There are numerous studies and reviews devoted to the mechanisms of burst formation in this small assembly of neurons which often was considered as a model of the simple “brain” (Maynard, 1961; Hartline, 1979; Wiens, 1982; Selverston and Moulins, 1985; Cooke, 1988, 2002).
Crustaceans possess an open circulatory system that lacks veins. The heart gives rise to seven arteries which lead the haemolymph to all organs and appendages. After blood leaves the arteries and lacunar ‘capillary’ spaces, it is channeled through defined sinuses and returns via the gills to the pericardial sinus. In lobsters, blood returns to the ventricle through three pairs of ostia. The valves in the ostia, like the myocardium, contain cross-striated muscle fibers (Yazawa et al., 1999), which are innervated by the cardiac ganglion.
In decapods, cardiac neurons are differentiated into large motoneurons and small interneurons which interact mutually via electrical and chemical synaptic connections to produce synchronous burst discharges. In lobsters, axons from the four small cells are confined to the ganglion while those from the five large cells exit the ganglion and innervate heart muscle. These neurons have 1-3 independent axons, small axon collaterals and neuropilar dendrites and dendritic arbor embedded in the heart muscle. The motoneurones of the cardiac ganglion branch extensively and each muscle fiber receives multineuronal input. The heart rate can be modulated by an intrinsic, probably “asynaptic,” stretch reflex which involves activation of dendrites in the heart wall. In addition, the cardiac rhythm is modulated by the inhibitory and acceleratory “extrinsic” nerve fibers. In the crayfish, there are two pairs of cardioregulatory nerves, known as Superior Nerves II and III (SN II, SN III). It was well documented that the SN II has inhibitory function whereas the SN III is acceleratory (Taylor, 1970; Field and Larimer, 1975a, b). These nerves are known to consist of several fibers but there is common opinion that the crayfish possess only one pair of inhibitory and one pair of acceleratory neurons.
Many other decapods (such as crabs and lobsters) have one pair of inhibitors and two pairs of accelerators. However, the exact location of neurons giving rise to these axons remains unclear. In fact, there are no recent investigations of this particular question. Only in a large isopod, Bathynomus doederleni, the cardioregulatory neurons were identified by current neuroanatomical and electrophysiological techniques (Tanaka and Kuwasawa, 1991a, b). One inhibitory and two excitatory neurons are located in the first, second and third thoracic ganglia, respectively. In the crayfish, the cardioregulatory nerves arise in the subooesophageal ganglion, near the mixed nerves going to the second maxilla (Wiersma and Novitski, 1942; Taylor, 1970; Field and Larimer, 1975a). They run to the pericardial sinus where some axons supply a pair of neurohaemal, pericardial organs, others supply a plexus on the suspensory ligaments and one fibre from each trunk is combined into a dorsal nerve, which enters the heart. Their terminals are in contact with the pacemaker neurons of the CG.
It is widely accepted that the GABA is neurotransmitter released by the inhibitory nerves in several crustacean species (Florey, 1960; Maynard, 1961; Yazawa, Kuwasawa, 1984; Wilkens and Walker, 1992) whereas the nature of neurotransmitter released by the acceleratory nerves remains controversial.
The intrinsic generator of heart rhythm is probably regulated by input from neurons located at different levels of the central nervous system. The above-mentioned cardioregulatory nerves are known to be tonically active (Taylor, 1970; Field and Larimer, 1975a; Young, 1978), therefore it was suggested that they might exert a tonic control over heart rate similar to vagal tone operating on the dogfish heart (Taylor, 1982). Besides, there are interneurones whose axons go in the circumoesophageal commissures that control the activity of the cardioinhibitor and cardioaccelerator neurons in the SOG (Field and Larimer, 1975b). These interneurons (‘cardiac command fibers’) fell into three classes: strong inhibitors, which caused cardiac arrest; weak inhibitors, which caused bradycardia and accelerators, which caused tachycardia. In the discussion, the authors noted: “Although we characterized a surprisingly large number of ‘cardiac command fibers’, more exist in other parts of the Central Nervous System (CNS)...”. They agreed with the idea of McMahon and Wilkens (1972) who postulated a central integrating circuit which receives all the appropriate afferent input and activates cardioinhibitors or cardioaccelerators to act suitably.
In whole animals, stimulus-induced changes in heart rate appear to be caused by this mechanism. These reactions were shown by many authors on different crustaceans (Larimer, 1964; Mislin, 1966; Cuadras, 1979; Tanaka, 1990; Shuranova et al., 1993; Bojsen et al., 1998). Thus the well established fact is that many sudden changes in the environment result in rapidly arising and short-lasting bradycadia whereas motor activation correlated with tachycardia. In fact this reaction is a component of more general changes of several functional systems, first of all, the ventilatory one.
Routinely, measures of an animal’s Heart Rate (HR) are used to assess the degree of excitability and ability to respond to stimuli. Thus, the crayfish demonstrate a rapid increase in HR during defense posturing (Listerman et al., 2000). The HR increases after a short time (<1 min) with a visual stimulus and associated defense posturing (Fig. 3).
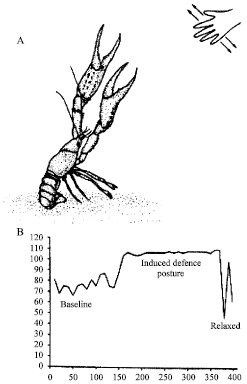 | |
| Fig. 3: | (A) A defense posture can readily be induced in crayfish by providing a moving shadow across their eyes. The rapid response of a meral spread, tail flexion and raising antennae coincides with an increase in heart rate (B). The elevated heart rate lasts for a few minutes in the absence of the stimulation. (Modified from Listerman et al., 2000) |
The rate returns to baseline, in most cases, within 5 min after a disturbance. Social posturing of a dominant crayfish (Procambarus clarkii) with a meral spread produced an increase in HR both within this animal and also in the submissive individual, which demonstrated no bodily movements (Listerman et al., 2000; Schapker et al., 2002).
However, only the initial part of these reactions is caused probably by direct nervous mechanisms. Comparison of intact lobsters with those that had both dorsal nerves cut showed that cardioinhibitor nerves are responsible for bradycardia induced by sudden external stimuli and the cardioaccelerator nerves cause the rapid-onset tachycardia associated with walking. In contrast, the continued gradually increasing heart rate during long walks on the treadmill is accounted for by hemolymph-borne factors including neurohormones (Guirguis and Wilkens, 1995). The same is true probably for long-lasting decrease in cardiac rate correlated with periods of inactivity (Gribble, 1994; Gribble and Broom, 1996).
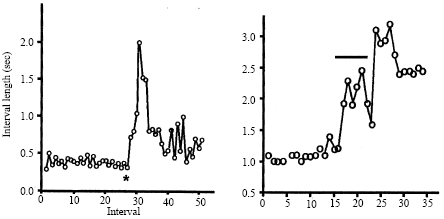
The fluctuations of heartbeat interval were investigated to assess cardio-regulatory nervous function in spiny lobsters (Yazawa and Katsuyama, 2001). The heart rate of freely moving animals exhibited the so-called on/off switching, i.e., an elevated and maintained rate was repetitively interrupted (with a period about 15 min) by periods of decreased rate. Each period was initiated by a sudden decrease in rate and was terminated by an exponential return to normal activity. Conversely, the steady state heart rate of the restrained animals was significantly higher and did not reveal such dynamics. This means that the acceleratory nerves are really highly active during times of stress. At the same time, an analysis of the return maps showed that in restrained animals only white-noise like fluctuations of heart rate are evident; this indicates that the brain may not strongly regulate the heart when the animal is kept under stressful conditions. A sudden marked decrease in heart rate followed by exponential recovery observed in freely moving lobsters is explained by a model based on the release and reuptake of the inhibitory transmitter, the GABA. Interestingly, this model can be also useful for explaining some parameters of the human heart activity.
The control of cardiac activity is not restricted by its effects on heart rate. It was established by several works of McMahon's (1995, 2001) and his collaborators on whole animals that heart rate and stroke volume can be controlled independently, thus measurements of both are needed to assess cardiac output accurately. In addition, the regional distribution of cardiac output is tightly controlled via cardioarterial valves at the base of each artery. These valves contain innervated muscle and differential contraction serves to regulate the efflux of oxygenated haemolymph into a particular system.
During treadmill walking activity, haemolymph flow through the sternal and anterolateral arteries of Cancer magister increased, while flow through the hepatic arterial system decreased. This indicated that oxygen supply was shifted away from the digestive system to the muscles of the walking legs and the respiratory system (Wachter and McMahon, 1996).
Arterial haemolymph flow patterns measured in restrained crayfish (Procambarus clarkii) and lobsters (Homarus americanus) remained constant during quiet conditions. Tail movements increased flow in both animals, although the general wave form of haemolymph flow and pressure did not change. In lobsters, periodic respiratory pauses were observed during which all arteries received hemolymph, despite the low heart rate (Reiber et al., 1997).
Changes in cardiac function and arterial haemolymph flow associated with 6 h of emersion were studied in the crab Cancer magister. The frequency of the heart beat began to decline sharply almost immediately after the start of the emersion period. Cardiac stroke volume fell more gradually. Haemolymph flow through all the arteries, with the exception of the anterior aorta, also declined markedly during emersion. As the water level in the experimental chamber fell below the inhalant branchial openings, a dramatic increase in haemolymph flow through the anterior aorta began and this continued for the duration of the emersion period. It was suggested that these responses are adaptations, suitable to conserve energy by reducing the minute volume of haemolymph pumped by the heart and to protect the supply of haemolymph to cephalic elements of the CNS (Airriess and McMahon, 1996).
In the above described experiments on the whole animals it is difficult to reveal the exact mechanisms acting on the cardiovascular system. Most probable, in this situation to the direct neural control a less direct influence of the CNS, exerted by neurosecretion of various active substances from the neurohemal organs, is added.
Ventilatory System
Oxygen uptake in decapod crustaceans occurs across the gills that are situated in a narrow branchial chamber. For renewal of the water near gills, these animals use a very efficient pumping system consisting of two specialized appendages, Scaphognathites (SG), one on each side, at the anterior ends of the branchial chambers. The SG which represents strongly modified second maxilla, is a blade like structure which moves from a fully levated (dorsal) to a fully depressed (ventral) position, drawing water across the gills by its rhythmic movements. The muscles responsible for SG beating (11 in the crayfish; Pasztor, 1968) are divisible to depressors and levators, each innervated by a separate nerve trunk (Pasztor, 1968; Pilkington and Simmers, 1973; Naynert, 1975; Young, 1975). The two nerves going in the crayfish from the hemiganglion are carrying impulses to these muscles alternately activating the depressors and levators.
The central pattern generator responsible for the production of ventilatory pumping (Ventilatory Pattern Generator, VPG) in crustaceans was originally thought to consist of a single endogenously oscillating non-spiking interneuron (Mendelson, 1971), but has later been shown to be the result of a network consisting of many neurons (Fig. 2B). In crabs where the VPG lies within the fused thoracic ganglion eight non-spiking interneurons were identified that exhibit large amplitude oscillations in their membrane potentials, which are in phase with the ventilatory motor pattern. These interneurons restricted to a single hemiganglion were suggested to be the primary components of the VPG in the crab (DiCaprio, 1989). There is also evidence that the ventilatory motoneurons have excitatory feedback to the premotor elements. Therefore it was suggested that these motoneurons also are components of the VPG (Simmers and Bush, 1983a, b). Three distinct interneurons of higher order were also demonstrated in crab’s thoracic ganglion. They can modulate the frequency of the ventilatory rhythm (DiCaprio and Fourtner, 1988) exerting their effect on both the right and left VPGs. It was suggested that these interneurons might be a link between the spiking ‘command’ neurons and the non-spiking interneurons which constitute a part of the VPG.
The so-called command neurons that can alter ventilatory rhythm were described earlier in the circumoesophageal connectives of the crab by Wilkens et al. (1974). These fibers (their maximal number within the whole connective is approximately 15) may carry impulses from higher centers in the central nervous system to exert their control (in about 70% of the cases) on both the ventilatory and circulatory systems. Unfortunately, the location of the nerve cells inside the cerebral ganglia was not established at the time when this study was performed. It should be noted too that the data about the VPG structure as well as about its central modulation were only obtained in crabs (Brachyura) whose VPG lies in the fused thoracic ganglion. There is no data on the crayfish (Macrura) whose VPG is located in the SOG.
The number of physiological studies devoted to VPG activity in crustaceans is greater than the anatomical reports. The activity of the VPG like that of the heart ganglion (or even to a greater degree) can be modulated by many factors. First of all, it is well known that the ventilatory activity is correlated with oxygenation of the ambient medium. Two mechanisms for its detection were demonstrated in different decapods: peripheral and central O2 chemoreception. In the crayfish, oxygen-sensitive activity was recorded from the branchial nerve which terminates in the branchio-cardiac vein wall and its surroundings. Impulse frequency in this nerve increased in response to hypoxia and decreased in hyperoxia. Baroreceptor activity was also observed (Ishii et al., 1989). In the crab, a population of peripheral O2-sensitive chemoreceptors is shown within the arterial system in the ventral anterior region, probably around the scaphognathites. These receptors are reversibly stimulated by agents known to be excitatory for vertebrate peripheral O2 chemoreceptors. This indicates that basic mechanisms of O2 chemosensitivity may be the same throughout the animal kingdom (Zinebi et al., 1990). There is no less importance in crustaceans, as well in vertebrates, for the VPG to continually receive feedback signals from chemoreceptors which adjust their activity in response to the environmental conditions. The isolated thoracic ganglion also responds to changes in oxygen tension (Wilkens et al., 1989).
Also, the activity of the VPG is known to depend on the animal’s functional state. In the experiments with the crayfish (Procambarus cubensis) reared in the laboratory, we recorded non-vasively the succession of potential waves, which represent the summed electrical fields produced by the SG-moving muscles, the so-called Electroscaphognathite-gram (ESG) (Burmistrov and Shuranova, 1996; Shuranova et al., 2003a). The crayfish, which could move freely, was in a vessel similar to its “home” tank and in the same ambient conditions that were “natural” to it. A regular ventilatory pattern with rather high (3-5 Hz) frequency is observed many minutes after fastening the electrodes for ESG recording (Fig. 4A). Similar tonic ESG was typical for a very high excitation level caused by various strong influences (such as shifting the animal into a new tank, putting a shelter in the tank etc.). However, observable behavior was rare. Regular high-frequency ESG was also observed in feeding and sexual behavior. It is remarkable that in the latter such ventilatory patterns were typical not only for running but also for motionless animals (Fig. 4B). Thus, there was no correlation between the ESG and the presence or absence of skeletal motor activity. A regular ESG pattern of low frequency and low amplitude are observed in the crayfish after they have been housed for two days or more in the same experimental tank. This coincided with its long-term immobility and non-reactivity to usual external stimuli. If the animal is awakened in some manner, then a high frequency and high-amplitude ESG is measured (Shuranova and Burmistrov, 2003a).
These studies suggest that ventilatory activity in the crayfish is regular when the animal is in one of the two extreme states (stressed or sleep-like) and when it is inactive behaviorally. In the intermediate state when the crayfish seems wakeful and behaviorally active its ESG is mostly highly irregular. The variability concerns both the parameters of single waves and general structure of the ESG in successive time periods. The spontaneous short-term variability of the ESG patterns is often correlated with different phases of walking: “bursts” of ESG waves are observed during “through”-forward walking and inhibitory changes in the ESG, during motor pauses, all of which are typical for crayfish walking. It should be emphasized however that the ESG may be highly irregular even in the totally immobile crayfish. This indicates that the variability depends on some uncontrolled afferent stimuli to the various sensory systems of the animal. A wide variety of optical, mechanical, chemical and osmotic stimuli are known to alter SG rhythms (Larimer, 1964; Wilkens and McMahon, 1972). Such studies were carried out however on immobilized animals, or ones restricted in their movements which are incapable on this account to have normal behaviors.
In experiments on free moving crayfish (Astacus astacus and Procambarus cubensis) the ventilatory reactions to various weak (behaviorally neutral) external stimuli unexpected by the animal were investigated (Shuranova et al., 1993). Such stimuli as a single drop of water falling on the surface of the tank, light tap outside the experimental chamber, turning light on and off, or a shadow, given with long intervals (not less than 30 min) to a quiescent animal, for the most part did not produce a obvious behavioral reaction. At the same time, these stimuli evoked marked changes in the ESG pattern. An initial component of ventilatory reaction for different stimuli reaching the crayfish through short perturbations of the water perceived mainly by crayfish’ mechanoreceptors consisted in a rapid and strong increase of the intervals between the successive ESG-waves (Fig. 5, left). Similar reactions were evoked by stimuli perceived by visual receptors, such as a moving shadow (Fig. 5, right) or switching the light off (Fig. 6A and B1). Switching from no light to light usually elicited less marked inhibitory changes in the ESG because this effect was often masked by excitatory changes in ventilation overlapping it and correlating with gradual development of motor manifestations. It should be noted however that the excitatory changes in the ESG arose later than the initial inhibition (Fig. 6A and B2). Thus stimuli of different modalities presented to alert, immobile crayfish elicited similar ventilatory reactions caused probably by the inhibitory commands from the brain.
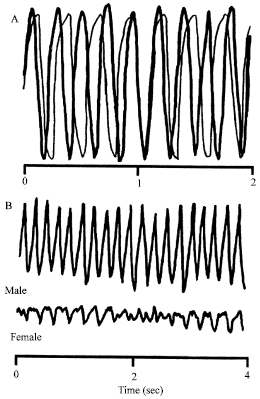 | |
| Fig. 4: | The regular high-frequency Electroscaphognathite-gram, (ESG) pattern typical for a highly excited crayfish can be seen (A) fifteen minutes after positioning a rod on the carapace (thick line) and two hours later (thin line). (B) A short fragment of the ESGs recorded in the motionless male (thick line) and female (thin line) crayfish placed in the same tank |
 | |
| Fig. 5: | Inhibitory reactions in the ESG caused by a single drop of water (left) and by a moving shadow (right). Abscissa, successive intervals; ordinate, number of intervals before, during and after sensory stimulus. (*a drop of water; thick line at right, “shadow” stimulus) |
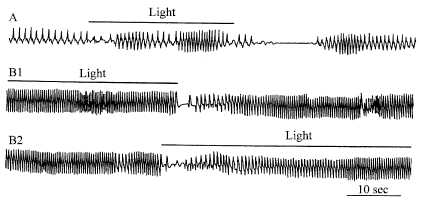 | |
| Fig. 6: | ESG reactions to illumination of the dark-adapted crayfish (A, B-different animals, B1-light off, B2-light on) |
It was suggested that even being immobile, the crayfish is continuously monitoring its surrounding and is revealing its slightest changes. Marked ventilatory responses (in fact they were usually coincident with similar changes in heart rate, Shuranova et al., 1993) of the crayfish to different external stimuli which strongly depend on its functional state but much less depend on the modality of the stimulus appear to be similar with those characteristic for the so-called “orienting response” of higher mammals (Graham, 1979; Pavlov, 1923; Zernicki, 1987). Thus, it was suggested that relatively “neutral” (“indifferent”) unexpected external stimuli might trigger in a crayfish’s brain some processing of the information about the “novel” stimulus and its possible consequences (Shuranova and Burmistrov, 1995; Burmistrov and Shuranova, 1996).
Coordination of Heart and Ventilatory Activity
The prominent feature of many ventilatory reactions in the crayfish is their obvious coordination with the cardiac reactions (Taylor, 1982). For instance, similar reactions in both systems to unexpected external stimuli were found in the experiments with simultaneous recording heart rate and SG beats rate (Larimer, 1964; McMahon and Wilkens, 1972; Wilkens et al., 1974; Young, 1978; Young and Coyer, 1979; Schapker et al., 2001; Shuranova et al., 1993). Wilkens et al. (1974) investigated the central control of this coordination in Cancer magister by stimulating command interneurons in the circumoesophageal connectives. They found that 68% of the effective fibers influenced both heart and SG systems, that cardiac arrest was always accompanied by SG responses in intact crabs and that most responses were inhibitory. Correlated activities in the cardioregulatory nerves and ventilatory system was demonstrated in the lobster (Young, 1978) and the activities were shown to be coordinated between the left and right SG’s and the heart and that the coordination was not altered by cautery of a major sensory organ in the SG, the oval organ (Young and Coyer, 1979). The authors concluded that the cardioregulatory neurons provide at least one route for coordinating cardiac and ventilatory activity. They also noted that the question of how the ventilatory and cardioregulatory systems are coupled in the ganglion (SOG) is an interesting one which remains to be answered. Unfortunately these observations were made about two decades ago and appear to be without further development.
At the same time, there is a great number of papers performed on different decapods indicating the correlated reactions of both systems during various exercises and changes in the environment (Taylor, 1982; Airries and McMahon, 1994; Wachter and McMahon, 1996; Gribble and Broom, 1996; Reiber et al., 1997; Rose et al., 1998). Recent data (Rose et al., 2001) show that the lobsters with normal heart function respond to the increased delivery requirements during walking with an increase in heart and ventilation rate. During exercise at two speeds, the ventilation rate increased by the same amount but ventilation volume was significantly greater at the fastest walking speed. Similarly, heart rate was elevated to the same level at different walking speeds, but cardiac stroke volume was greater at the faster walking. Noteworthy, that after impairment of cardiac function, ventilatory rate was significantly elevated. When cardiac output was more severely impaired, ventilation rate was greater. Thus the performance of one system changed to compensate for impaired function in the other.
Thus, the following points need to be emphasized: (1) the Ventilatory Pattern Generator (VPG) in crustaceans is under constant control of higher nervous centers; (2) there are many reports on the physiologically measured coordination of ventilatory and heart reactions provided probably by common brain structures but these structures have not been determined yet and (3) there also is no direct evidence about mutual interactions of the neurons forming the VPG and neurons whose axons are known as cardioregulatory ones, though they may be located near each other.
The Digestive System
Decapods are scavenger-predators, with ability to handle and masticate large pieces of food (the latter process is performed, however, in their stomach!). In normal eating movements, the mandibles act in coordination with accessory feeding appendages, especially the second and third maxillipeds. Their digestive system includes the mouthparts (mouth opening, mandibles, labrum and paragnatha), the foregut, midgut and hindgut with an anal opening.
The Foregut
At present, various components of the digestive system have been disproportionately studied. The doubtless favorite in this field is the foregut: nervous mechanisms underlying its function have recently been studied by a great number of researchers and many reviews are available (Harris-Warrick et al., 1989, 1992; Hartline and Maynard, 1975; Hartline et al., 1988; Marder and Hooper, 1985; Maynard and Selverston, 1975; Moulins and Nagy, 1985; Selverston, 1988; Selverston et al., 1976, 1998; Simmers et al., 1995; Wales, 1982).
Unlike the mouthparts, which are innervated by the SOG and unlike the hindgut and anus, innervated by the last abdominal ganglion, the foregut is largely innervated by a discrete extension of the CNS, the so-called Stomatogastric Nervous System (STNS). This system consists of paired commissural ganglia (CoGs), the Oesophageal Ganglion (OG) and the Stomatogastric Ganglion (STG), plus their associated nerve trunks.
The foregut of decapods consists of four regions: the esophagus, the cardiac sac, the gastric mill and the pylorus. The food enters the esophagus that moves it along by alternate contractions of the constrictor and dilator muscles. From the cardiac sac the food moves to the gastric region where it is shredded by rhythmic movements of three gastric-mill “teeth”. Finally, the small particles of the food pass into the pylorus and its peristaltic movements carry the pulp to the midgut where it is digested.
Primary control of the foregut musculature (about 40 muscles; Maynard and Dando, 1974) lies in the STG. The anatomical picture of the STG was studied by a light microscopy technique (Orlov, 1927). Like other decapod ganglions, the cell bodies lie at the periphery and the central part is occupied by the neuropil. Orlov claimed that the STG has a very simple structure: motor neurons, descending axons and some cell bodies of sensory neurons, which do not terminate in the STG but send their axons to the more anterior ganglia.
The more recent investigations of the STG in the lobster (King, 1976 a, b) and crayfish (Vladimirova and Fomichev, 1980) which used the electron microscopic technique confirmed these findings but also revealed that the motoneurons (their number is about 30) send processes into the underlying neuropil mass. The neuropil is differentiated into two regions: a core consisting of large heavily ensheathed processes and a surrounding region of fine-textured neuropil. The latter contains about one million synapses that are easily recognizable under the electron microscope by presynaptic vesicles and pre-and post-synaptic membrane specializations. Neuron counts in the STG of the crab showed that it contains 25-26 neurons (Kilman and Marder, 1996). In crab’s neuropil, not only usual synaptic contacts but also neurohemal-like profiles are seen, similar to those in neurosecretory regions of decapods. In addition, the junctions are described that may be real candidate for electrical junctions which are so important in the STG’s function (see below). Thus, despite the limited number of neurons, the STG appears to possess rather complex internal organization.
The motor output leaves the ganglion along a number of motor nerves which branch widely upon the wall of the stomach. The sole input to the STG is provided by the stomatogastrc nerve, which connects it with the CoGs and OG.
The neuronal structure of these ganglia was studied in detail by light microscopy 300 OGs and 600 CoGs were used in this study as mentioned by the Orlov (1929). The OG is not a typical crustacean ganglion. It contains a set of 15-16 nerve cells which lie in the junction of the inferior oesophageal nerves with the oesophageal nerve and do not surround the neuropil. In addition to cell bodies, in the OG are many kinds of nerve fibers. Some of them go through the OG without any contacts whereas the other have contacts with its elements. In comparison to the OG, the structure of the CoG is much more complex and stable. Each of the CoGs contains several hundreds of neuronal cell bodies (Rezer, 1987; Tierney et al., 1999). Orlov described many kinds of nerve fibers and nerve cells in the CoG. The most prominent feature of this ganglion is that it contains neurons whose axon enter the circumooesophageal commissure and direct towards the SOG, or neurons whose axons go towards the cerebral ganglia. Besides, there are nerve cells whose axon bifurcates: one branch goes towards the SOG and the other-towards the cerebral ganglia.
The CoGs and the OG receive their major inputs from higher centers and from foregut sensory receptors via a variety of foregut sensory nerves. Each of the paired CoGs gives rise to the superior and inferior oesophageal nerves, which travel medially to the OG. At this ganglion, these nerves are joined with the small unpaired inferior ventricular nerve that connects the OG with the cerebral ganglia. In the crayfish, this nerve consists of eight axons with diameters between 3.5 and 8 μm. Two of these axons terminate with their cell bodies within the inferior ventricular nerve; the other six axons spread inside the brain, mainly within the protocerebrum and the neuropils of the first and second antennae. Several fibers also send neuritis via the circumooesophageal connectives toward the CoGs and down to the ventral nerve cord (Boem et al., 1997, 2002; Heinzel et al., 2002).
The STG, like the CG, has been shown to function after its complete isolation from the body. This allowed the identification of all its neurons and their function so that “wiring diagrams” can be constructed. In decapod crustaceans, the STG is usually divided into two CPG circuits: the gastric mill CPG, which controls rhythmic movements of medial and lateral “teeth”, breaking the food and the pyloric CPG, which controls rhythmic pumping and filtering movements of the posterior part of the stomach.
In lobster, the pyloric muscles consist of the dorsal dilator muscles innervated by two PD motor neurons, the constrictor 1 muscles innervated by the single LP motor neuron and a set of constrictor 2 muscles innervated by 8 PY motor neurons. The 14 pyloric neurons, their synaptic connections and fast transmitters have all been identified and characterized (Johnson and Hooper, 1992; Miller, 1987; Mulloney, 1987). All the chemical connections within the network are inhibitory, using either Ach or Glu as transmitters; all other connections are via rectifying or nonrectifying electrical junctions. Although all the neurons are conditional bursters in the presence of appropriate neuromodulatrs, the rapidly oscillating Anterior Burster (AB) and the two electrically connected PD neurons normally function as the major pacemaker for the network, which produces a characteristic pyloric patterns (Fig. 2C).
Though for a long time the isolated STG has been a beloved model for studying problems related to the concept of the CPG, it should be stressed that its performance and reliability are greatly affected by some extrinsic factors. Without modulatory input to the STG no rhythmic patterns can be produced and the neurons fire tonically or they are silent (Russell, 1979). On the contrary, “when neuromodulators are released into the ganglion from specific neurons or are delivered as hormones, the properties of the neurons and synapses change dramatically and modulator-specific gastric mill and pyloric patterns are produced” (Selverston et al., 1998).
Modulatory inputs can alter every aspect of the CPG network, including (1) the neuronal composition of the active circuit, (2) the relative phasing of neuronal activity, (3), the functional synaptic wiring diagram and (4), the intrinsic properties of the cells. Furthermore, a single modulatory input can have different effects on the different cells in the circuit. “This extensive ‘sculpting’ of the CPG circuit by different modulatory inputs results in the formation of many alternative functional circuits from a single anatomically defined network, each capable of generating its own motor program” (Harris-Warrick and Flamm, 1986).
The main neuronal influences are descending. They are established by the neurons of the anterior ganglia, CoGs and OG, through their axons in the stomatogastric nerve. There is probably a single exception, namely the recently discovered sensory neurons named the gastro-pyloric receptors (Katz and Harris-Warrick, 1989, 1991).
At present, many nerve cells that modulate the activity of the STG networks were identified and the ways by which they influence the neurons of the STG were investigated in detail. Within them are the anterior pyloric modulator, whose cell body is located in the OG (Nagy et al., 1981; Nagy and Dickinson, 1983); the pyloric oscillator neuron in the CoGs (Nagy and Dickinson, 1983); modulatory proctolin neuron (Nusbaum and Marder, 1989); pyloric suppressor neurons with somata in the inferior ventricular nerve (Cazalets et al., 1990). Specific input from two identified interneurons, one excitatory and one inhibitory, that project from each CoG, is shown in the lobster’s gastric mill CPG (Combes et al., 1999).
It is impossible to describe here all the identified modulatory neurons at the anterior ganglia whose influence on the STG neurons is highly specific and complicated. As an example, it would be interesting to mention three proctolin-containing neurons in the crab Cancer borealis called the modulatory proctolin neuron (MPN; a symmetrical pair of somata in the OG) and modulatory commissural neurons 1 and 7 (MCN1, MCN7; each has its soma in the CoG) (Marder et al., 2002). All these projection neurons contain proctolin, but the MPN has GABA as co-transmitter and the MCN1 contains proctoline, GABA and CabTRP. Therefore, they influence the STG activity differently. For example, the MPN modulates the pyloric rhythm directly (by proctolin) and besides, it suppresses the gastric mill patterns releasing GABA to some CoG’s neurons. The MPN also seems to use GABA within the STG.
The Hindgut
There are only a limited number of papers devoted to the structure and function of the hindgut in decapods. These papers are sparsely distributed in about hundred-year long period from the first publication by Alexandrowicz (1909) and Shuranova et al. (2003b).
Alexandrowicz demonstrated through histological techniques that there is a nerve plexus on the outer surface of the hindgut (‘Grundplexus’) formed by the nerve fibers leaving the terminal abdominal ganglion. The nerve fibres from this plexus reach single muscle bundles where they are seen as ‘Endplexuses’. In addition, he revealed many (3-4 thousands) bipolar nerve cells that were distributed uniformly over the length of the hindgut. One process went to the hindgut’s channel and ended there between the epithelial cells whereas the other was directed to its outer wall and ‘on its way it formed contacts with the processes of other nerve cells’. These processes formed a net on the surface of the hindgut, which had close connections with the fibers of the ‘Grundplexus’ and ‘most likely, the fibers of both went to the muscles together’.
The second part of the same paper was devoted to physiological experiments on the isolated hindgut of the crayfish. He appeared to be the first who did such experiments. It was shown that the hindgut isolated from the abdominal nerve cord and placed into the Ringer solution contracted spontaneously for a long time (up to 36 h). The contractions existed not only in the whole hindgut but also in its fragments.
The results of the physiological experiments were very important for Alexandrowicz because they confirmed his anatomical data. He concluded that ‘the nervous system of the crayfish hindgut consists of autonomic nervous devices, which are connected with the central nervous system... The nerve leaving the last abdominal ganglion... has to control the automatic movements’ of the hindgut”.
Later, the anatomical findings of Alexandrowicz were mainly confirmed in the investigation of a Russian neuroanatomist, (Orlov, 1926). The discovery of the spontaneous motility of the hindgut made by Alexandrowicz was confirmed later in several papers (Ebara, 1969; Winlow and Laverack, 1972). It was shown that the hindgut has many forms of spontaneous activity, which does not depend probably on the presence of the terminal abdominal ganglion.
Thus up-to-date opinion is that the spontaneous contractions of the hindgut are myogenic what contradicts obviously Alexandrowicz’s view. A recent investigation sheds some light on the peculiarities of the contractile physiology of the intestinal muscles in the crayfish (Brenner and Wilkens, 2001). The main conclusion from this study is that the muscles of the hindgut differ of the skeletal muscles in the crustaceans. These muscles cannot be tetanized either by repetitive stimulation or by elevated potassium saline. They can work in trace amounts of external Ca2+ and they have some differences in the E-C coupling as compared to crustacean skeletal muscles. This may be a pre-requisite for the intestinal muscles to be capable of generating peristaltic contractions via endogenous pacemakers without neural input. The existing data are, however, insufficient in order to determine strictly the source of the spontaneous peristaltic activity of the intestinal muscles in crustaceans and a view about neural contribution cannot be excluded (Prosser et al., 1965; Wrong et al., 2003).
The structure of the hindgut efferent neurons in the last abdominal ganglion of the crayfish (their number is 35-48) differ markedly from that of somatic motoneurons. Their axonal projections consist of a thick primary neurite with lateral projections and limited arborization of varicose fibers in the ganglionic neuropil. They also send axonal processes to the ganglionic neural sheath, where they arborize profusely. A peculiar feature of the hindgut efferents is their extensive axonal arborization in the intestinal nerve root. All hindgut efferent terminals contain agranular vesicles (50-60 nm) and a few large granular vesicles (100-200 nm). It is suggested that the hindgut efferent neurons are involved in a broader range of regulatory processes, such as innervation of the intestinal muscles, intraganglionic integration, 'en route' synaptic modulation of efferent and afferent information and neurohumoral control of peripheral targets (Elekes et al., 1988).
The main features of nervous supply of the hindgut revealed by Alexandrowicz were confirmed in the EM-study (Elofsson et al., 1978). Large nerve bundles were found outside the circular muscle layer and smaller ones-between the two muscle layers. Close to the muscles, only thin fibres were seen. The neuromuscular junctions in the longitudinal and circular muscles of the crayfish took two forms: small terminals embedded in the cytoplasm of the muscle cell and terminals contacting protrusions from the muscle cell. Two types of vesicle-containing terminals were found. One type contains dense-core vesicles (about 80-110 nm), as well as synaptic vesicles (approx. 40 nm); the other type contains only synaptic vesicles. The nerve-muscle contact is not only multiterminal but also multisynaptic. Several synaptic sites have been identified between a single terminal and one muscle cell. It is suggested that in crayfish intestine there is a morphologic basis for at least a double innervation one of them being catecholamiergic.
The main source for catecholamines is however not in the last but in more anterior ganglia (Mercier et al., 1991). Though a small number of catecholaminergic neurons is present in the sixth abdominal ganglion, none of these appears to innervate the hindgut. Two neurons immunoreactive to tyrosine hydroxylase whose cell bodies are located in the third and fourth abdominal ganglia were found to contribute to nerve terminals which contain catecholamines. Each of them sends a single axon posteriorly into the intestinal nerve, where the axon bifurcates to supply the anterior and posterior hindgut regions. `The data are consistent with the hypothesis that DA is released as a neurotransmitter by neurons innervating the hindgut. However, as yet there has been no definitive demonstration that DA is localized within conventional synaptic terminals. It is suggested that the DA-releasing neurons act as motor neurons, or that they augment the effects of motor neurons releasing the DA at the peripheral level.
Extracts of crustacean hindguts have been found to contain DA (Mercier et al., 1991), orcocinin (Bungart et al., 1994), proctolin (Mercier et al., 1997) and a FMRFamide-like peptide (Mercier et al., 1997). All of them excite hindgut muscles, but their roles in controlling peristalsis have not been firmly established. It has shown recently that L-glutamate reliably increased tonus in isolated hindgut of the crayfish acting on both circular and longitudinal muscles (Wrong et al., 2003). Evidence is presented that the crayfish hindgut contains glutamate receptors which are very similar to those associated with arthropod skeletal muscles. Three effects of glutamate on hindgut contractions were observed: an increase in tonus, a transient increase in contraction frequency and suppression of spontaneous rapid contractions. Thus the main effect of glutamate is excitatory. The suppression of spontaneous phasic contractions may be caused by receptor desensitization.
Some other factors are shown to influence the peristaltic activity of the hindgut and among them are not only excitatory (e.g., CCAP; Audehm et al., 1993) but also inhibitory (allatostatines; Dircksen et al., 1999) agents acting probably by neurohumoral way.
Neurohumoral Mechanisms of ANS Regulation
In mammalian systems the release of adrenaline (i.e., epinephrine), from the adrenal gland, is associated with the sympathetic division of the ANS and is characterized by the ‘fight or flight’ reactions. The parasympathetic is more direct in neural regulation and does not appear to be associated with neuroendocrine factors such as with the sympathetic system. Adrenaline in vertebrates, has a wide range of actions on various target tissues. Primarily, it increases heart and ventilatory rates as well as systemically increasing blood pressure.
In crustaceans, an analogous peripheral ANS structure has not been found. However, they are known to possess a highly elaborated system of the so-called neuroendocrine organs (specialized structures in which nerve terminals come into contact with the circulatory system). They include the X-organ-sinus gland system, the pericardial organs, prothoracic gland, green gland and neural endocrine cells within the nerve cord.
The ‘X-organ’ is a tightly clustered group of neurons on the ventral margin of the medulla terminalis whose axons run to the ‘sinus gland’-a structure that is very well exposed to hemolymph. This is the major neuroendocrine control center in crustaceans probably homologous with protocephalic neurosecretory pathways in other arthropod groups. These pathways are involved in processes of molting, growth, sexual maturation and regulation of metabolism. The crustacean sinus gland is also a source of hormones that mediate adaptations of metabolic functions to environmental conditions. At least 90% of the terminals in the sinus gland represent terminations of neurosecretory somata of the X-organ. Functionally, this system seems to be similar to the main autonomic center in vertebrate’s brain, the hypothalamus, connected directly with the anterior lobe of the hypophysis (Fingerman, 1997). A neuroendocrine or modulatory function is supposed also for a division of the protocerebrum, the “central body” (Sandeman, 1982; Schürmann et al., 1991).
The so-called “pericardial organs” lie in the venous cavity surrounding the crustacean heart. Their anatomy is very complex; in brief, they consist of the ligamental nerve plexuses formed by the neurosecretory axons of the “dorsal nerve” apparatus. Neuronal somata known to contribute their processes to the pericardial organs are incompletely studied. They include C and B cells located in the brachyuran thoracic mass and probably presented in crayfish ventral ganglia, intrinsic PO cells and the “root cells” found at the first branch point off each thoracic segmental nerve in Homarus. A prominent input to the pericardial organs comes from paired large dopaminergic cells of the commissural ganglia (Cooke and Sullivan, 1982). Pericardial organs are specialized for the production and storage of octopamine, dopamine and serotonin, as well as some peptides (proctolin, cardioexcitatory peptide, etc.) for release into general circulation.
These structures are designed for rapid release of substances which lead to alterations in sensory and motor function; thus altering the behavior of the animal. Rapid whole organism effects can be initiated in this hormonal manner while more direct effects can be insured with direct innervation. The integration of the various inputs on a target tissue can be complex when considering modulation of both pre-and postsynaptic sites.
Considering that the endocrine compounds and hormones released into the hemolymph cause acute responses (Fingerman, 1995, 1997), crustaceans are similar to insects which release numerous hormonal compounds from neural secretory structures to alter behavior of the whole animal. Evidence suggests that exposure of 5-HT to X-organ/sinus gland complex can result in a rapid release of the crustacean hyperglycemic hormone which maybe related to acute and chronic stress-induced hyperglycemia (Keller and Beyer, 1968; Lee et al., 2000, 2001).
The neural secretory cells within the ventral nerve cord, like those in the pericardial organ, are known to release 5-HT and OA (octopamine) rapidly into the hemolymph as well as within the neuropile within the ventral nerve cord to have immediate actions on neural tissue (Livingstone et al., 1981; Langworthy et al., 1997). For this reason, both 5-HT and OA may be considered “quick-response” compounds. The speed of their release from neural secretory cells and effect upon crustacean and insect behaviors have attracted most of the attention over the years. Therefore, we have highlighted the actions of these two factors on internal organs that are tightly regulated by the ANS in higher organisms.
The neurosecretatory cells containing 5-HT and OA within the VNC have their cell bodies and processes well characterized via immunocytochemistry staining in adults and throughout development in lobsters (Beltz et al., 1992; Schneider et al., 1993). A large number of varicose endings of neurons are located within the 2nd nerve roots of the ventral nerve cord ganglia, thus suggesting release sites directly into the hemolymph (Beltz et al., 1992; Evans et al., 1976a, b). Ultrastructural analysis of these terminals show many granular filled vesicles (Livingston et al., 1981). At least a 100 cell bodies have been shown to have 5-HT immunoreactivity in the lobster nervous system. Many of these cells are located within the SOG and it appears that each ganglion in the ventral nerve cord has at least one cell body that contains 5-HT (Langworthy et al., 1997). There are 2 extraordinary large pairs of 5-HT containing cells that exist in the 5th thoracic and 1st abdominal ganglia (Beltz and Kravitz, 1983, 1987). It is known that if the animals are stressed during a sacrifice and dissection the staining profiles are not as robust as compared to animals chilled slowly and then sacrificed. This suggests again that a rapid release of 5-HT occurs due to stress of the animal. Curiously, recent findings in crayfish by Musolf and Edwards (1999) and Beltz et al. (2001) as well as in frogs by Ten Eyck (1999) have demonstrated that 5-HT can be taken up into cells which may not synthesize 5-HT but can store the compound and then release it upon excitation of the cell.
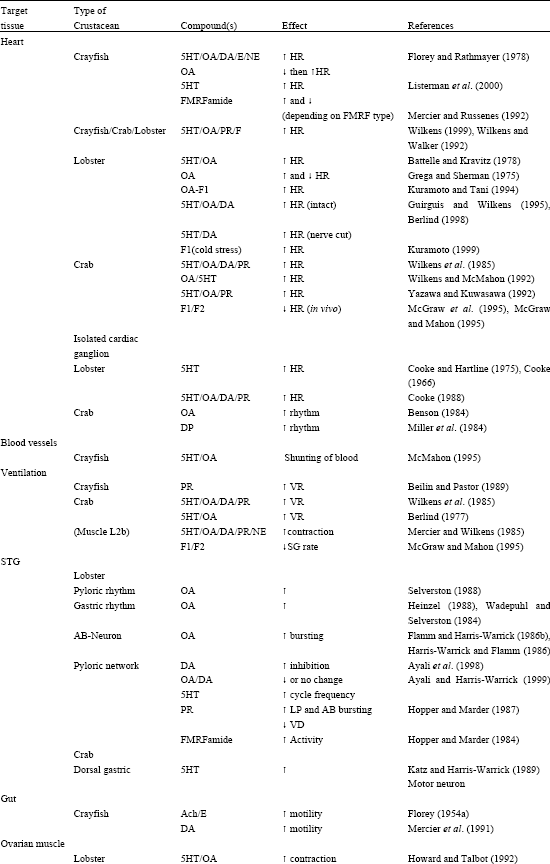
The direct effects of various neuroendocrine substances have been investigated in crustaceans (Table 1). Florey began in the 1950's assessing the effects of ACh and adrenalin on gut motility and skeletal muscle contraction with the assumption that these compounds were hormonal (Florey, 1954 a,b; Florey and Florey, 1954). Some of the same neurosecretatory cells release substances within a neuropile or to particular targeted neurons in a ganglia serving a more direct role in neuromodulation (Beltz and Kravitz, 1983, 1987; Siwicki et al., 1987; Livingston et al., 1981; Evans et al., 1976 a, b). Studies from Kravitz’s group (Livingston et al., 1980) with lobsters and crayfish demonstrated that injections of serotonin and octopamine into the circulatory system can cause the animals to assume various postures similar to those observed during social interactions. These studies led to further investigations of the anatomical location and physiological functions of the neurons that packaged and released such substances (Beltz and Kravitz, 1983). In addition, more effort was put into understanding the role of the compounds on sets of known neurons in lobsters (Harris-Warrick and Kravitz, 1984) and in crayfish (Breen and Atwood, 1983).
Peripheral actions are broad for 5-HT and OA in crustaceans. Both monoamines have been shown to alter HR, ventilatory rate, presynaptic release at neuromuscular junctions, postsynaptic input resistance of muscle cells, tactile and proprioceptive sensory neurons responsiveness to stimuli, visual response and properties associated with digestive processes (Table 1). Other neuromodulators such as proctolin, dopamine, adrenalin and noradrenaline as well as various forms of FMRFamides have been examined to their effects on target tissue in crustaceans and in insects (Table 1). Reviews of octopamine (Roeder, 1999) and 5-HT (Weiger, 1997) discussing the localization and function suggests homologous actions in a variety of arthropods.
Heart Rate
Alexandrowicz (1932, 1953) suggested that the pericardial organs in crustaceans represented neuroendocrine organs releasing substances to directly act on the heart. It is now known that the CGs of various crustaceans are influenced by this and other neurohemal inputs (Cooke and Sullivan, 1982; Grega and Sherman, 1975; McGaw et al., 1995; Wilkens, 1999; Wilkens and McMahon, 1992; Yamagishi et al., 1997).
The excitatory actions of 5-HT on the cardiac rate in crustaceans have been shown in both in situ and in vivo preparations (Florey and Rathmayer, 1978; Wilkens, 1995, 1999; Wilkens and Mercier, 1993; Wilkens and Walker, 1992).
| Table 1: | Direct effects of various neuroendocrine substances have been investigated in crustaceans |
 | |
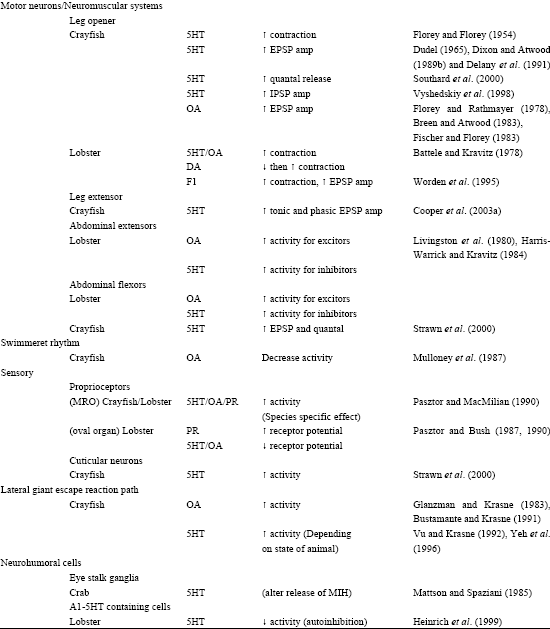 | |
| Abbreviations: 5HT-Serotonin; OA-Octopamine; DA-Dopamine; E-Epinephrine; NE-Norepinephrine; PR-Proctolin; HR-Heart Rate; VR-Ventilatory Rate | |
Octopamine also has an excitatory effect on HR if innervation to the cardiac ganglion remains intact (Guirguis and Wilkens, 1995). Pronounced differences were observed in responses to FMRFamide-related peptides for semi-isolated and intact hearts as is the case for the actions of 5-HT and OA (McGaw et al., 1995). Such differences in effects of neuromodulators in intact and isolated systems emphasizes the need to consider responses in the whole animal to relate neuromodulator effects to behavior.
In whole animals (Procambarus clarkii), it has recently been shown that HR can quickly change, within a minute, during injections of 5-HT into the animal’s hemolymph, at a circulating concentration of 100 nM (Listerman et al., 2000). An elevated HR for more than 10 h in adults can occur with concentrations in the range of 1 μM. The results of this recent studies of HR measures and effects of 5-HT, indicate that HR measures may be a good bioassay of endogenously released 5-HT since HR is rapidly altered by exogenous application or by systemic injections. The results of their study do not support the finding that 5-HT is elevated during social interactions, or chronically in aggressive individuals since HR can rapidly decrease during an intense social interaction and that a elevated HR is not observed for dominate crayfish within a dyiad when the animals are not interacting (Li et al., 2000; Listerman et al., 2000).
Ventilatory Pattern Generator
The evidence to date suggests that the cardiac and ventilatory responses to stress are likely due to neuroendocrine modulation (Wilkens et al., 1985). There are, however, only a few studies about the effect of amines and peptides on the VPG in crustaceans. Berlind (1976) discovered that extracts of the pericardial organs of crabs injected into intact crabs increased the beating frequency of the ventilatory appendages. He suggested that the active factor was not Dopamine (DA) or 5-HT, but probably it was a peptide. Later the peptide was identified as proctolin. There is now ample evidence for the presence of proctolin in the ventilatory system of Macrura. In lobster and crayfish a large cluster of somata with proctolin immunoreactivity was found close to the ventilatory neuropile (Siwicki and Bishop, 1986). Double-staining experiments using lucifer-yellow backfilling of the SG nerve followed by proctolin antibody binding and visualization with rhodamine-labeled secondary antibody (Pasztor and Marder, unpublished observations; Beilin and Pasztor, 1989) confirmed that some of SG motoneurons are proctolinergic. It is known that OA, 5-HT and proctolin are present in the pericardial organs, the anterior ramifications of which distribute prolific neuroendocrine endings across the membrane bounding the articular cavity of the SG. The effect of proctolin was obvious both in whole-animal experiments and in studies of single afferents from the ‘oval organ’ of the SG (Pasztor and MacMillan, 1990) and the ventilatory muscles (Beilin and Pasztor, 1989).
The effects of OA and 5-HT on the VPG are, however, not so clear. Berlind (1977) found that 5-HT decreased the frequency of bursting in the nerve trunks containing motor neuron fibers to the ventilatory muscles and increased the duration of pauses in motor output. No effect of DA, NA, OA and GABA were shown in these experiments. Different results were obtained however by Rajashekar and Wilkens (1992). They found that DA caused a dose-dependent increase in the frequency of motor neuron bursts produced by deafferented crab’s VPG. The effect of nicotine is tri-phasic, beginning with an initial reversal of burst pattern followed by a period of depression and a long period of elevated burst rate. On the contrary, 5-HT had no direct effect on the VPG. These observations may support the hypothesis that dopaminergic pathways are involved in the maintenance of the VPG rhythm and that the acceleratory effects of nicotine may involve release of DA at intraganglionic sites. Thus, it appears that 5-HT may have indirect actions and may even alter DA release. Probably the question about involvement of various amines in control of ventilatory function in the crayfish needs to be clarified in the future.
Digestion
Some of the earliest studies established that without modulatory input to the STG no rhythmic patterns can be produced and the neurons fire tonically or they are silent. “When neuromodulators are released into the ganglion from specific neurons or are delivered as hormones, the properties of the neurons and synapses change dramatically and modulator-specific gastric mill and pyloric patterns are produced” (Selverston et al., 1998).
Modulatory inputs can alter every aspect of the CPG network, including (1) the neuronal composition of the active circuit, (2) the relative phasing of neuronal activity, (3), the functional synaptic wiring diagram and (4), the intrinsic properties of the cells. Furthermore, a single modulatory input can have different effects on the different cells in the circuit. “This extensive ‘sculpting’ of the CPG circuit by different modulatory inputs results in the formation of many alternative functional circuits from a single anatomically defined network, each capable of generating its own motor program” (Harris-Warrick and Flamm, 1986). For several decades, the STG of decapod crustaceans has been used extensively as a model system for studying of modulatory effects of several monoamines and neuropeptides on the activity of neural circuits (Ayali et al., 1998; Ayali and Harris-Warrick, 1999; Flamm and Harris-Warrick, 1986a, b; Heinzel, 1988; Katz and Harris-Warrick, 1989; Nusbaum et al., 2001; Selverston, 1988; Wadepuhl and Selverston, 1984; Swensen and Marder, 2000). Modulator distribution has been studied using techniques such as immunocytochemistry, high performance liquid chromatography and radioimmunoassay. These studies have shown that many modulatory compounds are contained within neuronal somata situated in the CoGs and the OG, all of which send axons to the STG (as previously indicated). In addition, the STG circuits are modulated by some sensory cells which release neuromodulators in the STG in response to chemosensory cues or stretch in specific stomach muscles. DA and OA were the first of the neurotransmitters present in fibers of the stomatogastric nerve to be intensively studied. In addition to the large DA-containing neuron in the CoG, Kushner and Maynard (1977) described fluorescence, attributed to the DA, in several small somata in the CoGs, in fibers in supraoesophageal and stomatogastric nerves and in neuropilar processes in the STG. The action of DA on the STG is complex and varies during the time of application. The same is true for OA.
It is known that DA, 5-HT, proctolin and OA have different abilities to modulate activity within the STG (Ayali and Harris-Warrick, 1999; Johnson and Harris-Warrick, 1997; Flamm and Harris-Warrick, 1986a, b). This may have a significance within the intact animal since the distribution of the neurons containing these neuromodulators within the ganglion are not equal, thus some sites are more strongly modulated by one particular modulator or a mix of modulators of different strengths more so than at other sites in the neural circuit.
Each species may even have a varied distribution of occurrence of the modulators, for example 5-HT is found throughout many regions of the STG but it’s occurrence is species dependent (Beltz et al., 1984). In addition, depending the receptor subtypes and the circuitry, modulators may have excitatory as well as inhibitory influences. For example, in the pyloric network DA is one of the main substances to modulate the activity and it is able to excite as well as inhibit depending on the site of action and activity within the circuit (Harris-Warrick et al., 1998). Selectivity of neuronal action by proctolin on key neurons can also change the entire dynamics of the network’s activity profile (Blitz and Nusbaum, 1999; Hooper and Marder, 1987). Anatomical evidence demonstrates that in some cells 5-HT and DA are synthesized together. One particular neuron containing 5-HT and DA affects the STG activity and is known to arise from the CoG while also sending information of the rhythms to the brain or thoracic nervous system (Johnson et al., 1994). Other neuromodulators, for example FMRFamides have been shown to increase the frequency of activity of the STG in lobsters (Hooper and Marder, 1984) and are known to be present in the STG and inputs to the hindgut of crayfish that can regulate circulation and defecation (Mercier et al., 1991; Tierney et al., 1997). 5-HT immunoreactivity in the stomatogastric nervous system has been investigated in seven species of crayfish and there are notably differences among species (Tierney et al., 1999), which may translate into differences in experimental results in investigating the influences of neuromodulators in STG activity.
The evidence mentioned shows that under neuromodulatory instruction, individual neurons can participate in different oscillatory motor circuits and hence more than one rhythmic behavior and even more profoundly, preexisting networks can be dismantled to specify dynamically a new circuit for an entirely different behavior. This de novo network construction is achieved again by neuromodulatory-induced alterations in the oscillatory and synaptic properties of individual target neurons. On this basis, therefore, a functional CPG network must be seen in a more dynamic context than previously thought since it may exist only in a particular behavioral situation dictated by modulatory influences.
Effects on the Skeletal Muscles and Sensory Systems
The systemic release of neuromodulators effects all the neuromuscular tissues that are bathed by the hemolymph, much like the release of adrenalin by a sympathetic response in mammals. Exogenous application of 5-HT or octopamine on neuromuscular junctions has been shown to enhance the Excitatory Postsynaptic Potential (EPSP) amplitude of many preparations examined within crustaceans (Worden, 1998) such as the abdominal flexor and extensor muscles (Table 1) and muscles within the limbs (Table 1). Little is known about the direct mechanisms of action on the neurons, with the exception of a few studies that have shown that 5-HT might work through an inositol triphosphate (IP3) and cyclic AMP (cAMP) second-messenger system (Dixon and Atwood, 1989a, b; Dropic et al., 2005). Octopamine was shown to increase cAMP levels in skeletal muscle (Batelle and Kravitz, 1978). Interestingly, dopamine initially will decrease muscle contraction followed by enhanced contraction upon nerve stimulation (Batelle and Kravitz, 1978). Recent evidence also demonstrated inhibitory as well as excitatory responses for octopamine on transmitter release from motor neurons which may be dose dependent or depends on the behavioral state of the animal (Djokaj et al., 2001). The influence of the modulators are known to mainly act at the sites of release among presynaptic neurons (Cooper et al., 2003a; Southard et al., 2000; Strawn et al., 2000).
As stated by Bicker and Menzel (1989), “Although behavioral responses to neuromodulators are becoming better understood much remains to be explored in sensory systems and effects on circuitry”. Investigations have been underway to understand the effects of neuromodulators on individual target cells (sensory, CNS, motor command and muscles) as well as their roles in the integration of these systems, but many aspects of neuromodulation on circuits remain unknown. The roles are complicated by the fact that neuromodulators work in concert with other neuromodulators (Christie et al., 1995; Djokaj et al., 2001). Cocktails of neuromodulators and their mixed action on sensory systems is an avenue for future research. The complexity of the vertebrate nervous system makes it difficult to address the effects of neuromodulators on entire pathways which can regulate a specific behavior whereas in crustaceans some the complexity is manageable. Strawn et al. (2000) examined the effects of 5-HT on a complete neural circuit within the crayfish to address the role of a neuromodulator within an entire circuit. The results of this study demonstrate that 5-HT increases: (1) activity of the sensory neurons which drive the motor neurons; (2) the intrinsic activity of superficial flexor motor neurons; (3) vesicular release properties of the superficial motor nerve terminals and (4) input resistance of the muscle directly. The influence of sensory neuron activity on the drive of the 3rd root has also been shown to be enhanced by 5-HT. The neurohemal release of 5-HT can modulate not only the motor neurons but also sensory input and its drive within a pathway.
A number of studies have demonstrated that 5-HT heightens the responsiveness of various sensory systems within crustaceans. Specifically, studies of modulator effects on crustacean joint proprioceptors and muscle-tension receptors (Cooper et al., 2003b; Pasztor and Bush, 1987), as well as on the abdominal MRO’s have been used to investigate 5-HT and other potential hormones (Pasztor and Macmillan, 1990). As for the neuromuscular preparations, the sensory systems are exposed to compounds that are contained within the circulating hemolymph.
The visual system in crustaceans is also thought to be directly influenced by hormonal components. Visual acuity within the compound eyes of crustaceans has been correlated with the fluctuations of hormones in the animal’s hemolymph and neural tissue (Aréchiga et al., 1993; Aréchiga and Huberman, 1980; Castaéon-Cerantes et al., 1999; Kulkarni and Fingerman, 1992). It is thought that serotonergic innervation from the cerebral ganglion may be responsible for the diurnal patterns and visual sensitivity (Castaéon-Cerantes et al., 1999; Sandeman and Sandeman, 1990).
Similarly, in mammals, the regulation of the iris of the eye is by autonomic control, which effects the visual acuity of the animal. Regulation of visual ability in crustaceans by 5-HT as well as by direct innervation therefore suggests, at least correlatively, that one might infer a comparable ANS. In crustaceans the regulation may be mainly via hormonal factors since crustaceans have fewer nerve cells in their CNS.
Behavioral Responses
Survival of an animal in an ecological arena requires the use of many predation-avoiding behaviors, one of which is an escape behavior. Field and laboratory observations indicate that crayfish exhibit a wide range of rapid behaviors to combat threatening stimuli as well as changes in the environment, thus indicating an ability to assess and respond for the circumstance. Since various degrees of a response can be initiated by an individual under differing physiological and environmental conditions, we speculate that motivational drive or the internal state of the animal may have a role in how an animal responds. Crayfish also demonstrate ‘fear or fight or flight-type’ responses during social interactions with other crayfish as well as to environmental stimuli.
Even the earliest behavioral studies in crustaceans have noted the existence of rapid defense postures in crayfish (Bethe, 1897; Huxley, 1880; Wiersma, 1961). The activity of the ANS may well set the behavioral outcome in these instances. It is known that in socially interacting crickets (Hofmann and Stevenson, 2000) a submissive individual can regain its aggressiveness with the same opponent after it is given physical disturbance of a few seconds. It is thought that the animal is ‘reset’ by the stress of a strong disturbance.
Experiments by Kellie et al. (2001) with adult Procambarus clarkii showed that there is a substantial increase in tail flipping behavior of the intermediate sized animals that were induced to autotomize their chelipeds, as compared to a control group with both chelipeds intact. This study indicated that there is a rapid alteration as well as a long-term alteration in the telson-induced response which enhances the ability of the animal to tail flip (Fig. 7). Because of the rapidity of this change in responsiveness, it is likely that an analogous vertebrate sympathetic system is functioning in the crayfish to alter the behavioral state of the animal causing it to be more receptive to neural activity and enhancing motor command. Perhaps the long-term effects are due to a chronic stress effect on the animal mediated through hormonal factors as can also occur in mammals (Pagé and Cooper, 2004).
Many above-mentioned studies indicate participation of the heart and a tightly connected ventilatory system in various kinds of behavior in decapod’s. They can be supplemented with recent evidence devoted to studying cardiovascular component of the context signal memory in the crab, Chasmagnathus granulatus (Hermitte and Maldonado, 2005). The sudden presentation of a rectangular screen overhead elicited crab’s escape reaction and heart arrest followed by bradycardia. After training, there was a decrease of escape reaction, suppression of heart arrest and attenuation of bradycardia. In authors’ opinion, the results of this study “strongly suggest that decapods are altogether capable of displaying a response in essence similar to that of vertebrates mediated by the autonomic nervous system”.
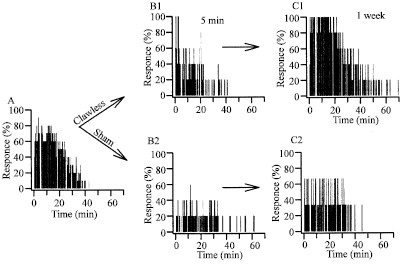 | |
| Fig. 7: | The rapid effects of claw removal on habituation rates in intermediate sized crayfish demonstrates the neural plasticity of the circuit. Ten crayfish were tested (A). Immediately following the test, 5 of the crayfish had their chelipeds removed and were retested (B, clawless). The other 5 crayfish were held as if their chelipeds were removed (B, sham). One week later, the same groups were reexamined (C). Relative cumulative frequency plots indicate that the crayfish with both chelipeds removed continued to tail flip longer than the sham control group. (Modified from Kellie et al., 2001) |
Less is known about involving other internal organs in behavioral reactions but it is well documented that the release of urine in the crayfish, Astacus leptodactylus, is greater during fights than during inactivity or non-social activities (Breithaup and Eger, 2002). Thus, the process of urine release coupled to offensive behavior is probably under the control of the ANS. In this experiments, the crayfish, Procambarus cubensis, put in a novel experimental chamber, often had a higher level of defecation than in its home tank (Shuranova et al., 2005). This phenomenon is well known in the experiments on rats where it is considered as indication of the high level of emotionality (Silverman, 1978). Thus, similar forms of environmental stress in vertebrates and the associated ANS responses are analogous in crayfish.
CONCLUSIONS
The main conclusions drawn from the above described data are listed below and presented schematically in Fig. 8.
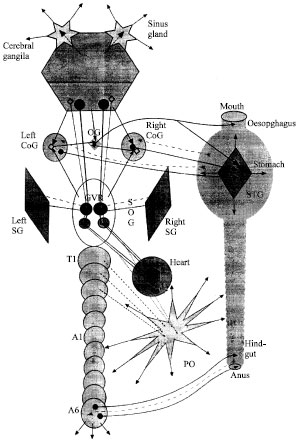 | |
| Fig. 8: | Schematic drawing of nervous regulation of main internal organs in Decapods The targets considered include heart muscle (Heart), muscles moving the symmetric ventilatory appendages, scaphognathites (Left SG, Right SG) and the muscles of the digestive tract (Mouth, Oesophagus, Stomach, Hindgut and Anus). They are triggered by an ‘intrinsic’ nerve nets: Cardiac Ganglion (CG), Generator of Ventilatory Rhythm (GVR), the Stomatogastric Ganglion (STG), Oesophageal Ganglion (OG) and neurons of the last abdominal ganglion (A6) (motor output signals are noted by solid arrows, backward input signals-by dotted arrows). The cardioregulatory neurons are within the Subesophageal Ganglion (SOG, near the VPG; they exert direct inhibitory (blue arrows) or acceleratory (red arrows) influence of the neurons comprising the CG. There also are many ‘command’ neurons of higher order issuing from the Cerebral ganglia; they effect both on the cardiac and ventilatory functions, with the inhibitory effect (blue arrows) dominating over the acceleratory (red arrows). The STG is under the control of paired commissural ganglia (Left CoG, Right CoG) and the OG. Their also is a direct pathway from the cerebral ganglia to the stomatogastric nervous system (the inferior ventricular nerve, a thin black arrow to the OG). A region within the cerebral ganglia that is assumed to regulate some metabolic functions is shown (Central Body, CB). Some neurohemal structures that exert widespread and strong modulatory influence (solid red arrows) on the activity of the internal organs are presented: the X-organ-Sinus gland system (Sinus gland), Pericardial Organs (PO) and neurons of the A6 (dotted red arrows to the PO indicate some known inputs to it) |
| • | In decapods, the cardiac ganglion, located inside the myocardium is unique in its structure and may be considered analogous to the intrinsic AV node in the vertebrate though the later has a non-neural origin. |
| • | The cardiac ganglion has inhibitory and acceleratory external control that may be analogized with vertebrates' parasympathetic and sympathetic systems. In both neurons, tonic spontaneous activity is similar to that in vertebrate parasympathetic and sympathetic neurons and their direct synaptic action on the neurons of the cardiac ganglion is well documented. At rest, the cardiac ganglion is predominantly under the influence of the inhibitory neuron (similar to the parasympathetic tonus in the mammalian heart under basal conditions). |
| • | Cardioregulatory neurons and ventilatory pattern generators are proximately linked like the respiratory centers and nuclei ensuring cardiac control in vertebrates. |
| • | There is common control of the cardioregulatory neurons and neurons comprising the ventilatory pattern generator by probably the same neurons of the cerebral ganglia. |
| • | Crustacean cardiac and ventilatory functions are well coordinated by direct synaptic and indirect neuromodulatory and/or neuroendocrine systems (from central neurons) like in vertebrates. |
| • | The stomatogastric ganglion governing the foregut muscles may be analogous with the enteric nervous system of vertebrates; as distinct from vertebrates, it is located outside the gut and it is known to innervate not the smooth but the striated muscles. These muscles are, however, slow and tonic, functionally similar to muscles of the alimentary tract in vertebrates. |
| • | The remarkable feature of the STG is that it contains very few (about 30) neurons and has no direct afferent input. At the same time, there is approximately one million synapses in its neuropil (King, 1976 a,b) which indicates a very sophisticated external control of its activity. This is in contrast with the enteric nervous system in mammalians that contains the enormous number of neurons (e.g., in humans about 108) but has few extrinsic nerve fibers (Gershon, 1987). Thus the STG is crustaceans seems to be more dependent of the extrinsic neural control than the enteric nervous system of mammalians. |
| • | The main input to the STG goes from the CoGs and OG and there is the great dependence of the ability to generate rhythmic pattern in the STG on the inputs from these ganglia that may be considered as the high-order centers in relation to the STG. |
| • | At present, many modulatory neurons in these ganglia are identified and their effects on the STG are studied in detail. The effects caused by each of these identified neurons are usually highly specific and widespread, concerning many neurons in the STG and resulting in deep transformation of the existing rhythmic pattern. It is obvious, however, that the effects caused by “modulatory” CoG neurons differ markedly from those caused by the cardioregulatory neurons on the CG: in the strict sense, this is not modulation but triggering some new CPG circuits. |
| • | The same is true for the effects of different amines (5-HT, DA, OA) and peptides (proctolin, FMRFamide, etc.). Thus, the role of some “higher” neurons in the control of the “intrinsic” nervous system of the decapod gut is obvious; it is unclear, however, in what degree this control system is similar to the sympathetic and parasympathetic systems governing the digestive tract in vertebrates. |
| • | Taking into account all the above-mentioned difficulties and uncertainties there is still plenty of evidence to suggest that the crayfish and other decapods have a sophisticated nervous control of their internal organs similar in principle to that of the ANS in vertebrates. In addition to “intrinsic” nerve nets triggering movements of the muscles of the internal organs, there are neural centers of higher order that strongly modulate these movements and influence other aspects of activity of the internal organs. The very important role in this modulation appears to belong both to the direct synaptic and to the more distributed neuroendocrine mechanisms, the latter may be especially important in crustaceans, with their surprisingly parvocellular nervous system. |
Future studies in the crustacean autonomic nervous system
There are numerous future studies which would greatly aid in helping to delineate aspects of the ANS in crustaceans. Here we mention some topics of immediate need in this area. Attention to anatomical studies will enhance our general understanding. For example, microanatomical investigation within the SOG is needed for identification of the cardioregulatory neurons and to aid further description of the VPG. Mapping the anatomical and physiological interrelations between both the cardiac and ventilatory networks is needed since they both are obviously involved in functional regulation of internal organs. In addition, more precise description in the paths and destination points of all the nerves going in and out the SOG would greatly enhance the knowledge in the function and architecture of the ganglion.
If the anatomical paths are mapped than one might be able to better determine which functional paths to examine physiologically. Identification within the cerebral ganglia of the origin for 'command' fibers to the cardioregulatory center and to the VPG would provide information to what might drive these systems. In addition, much would be gained by knowing which regions within the cerebral ganglia are connected with the control of visceral functions including the digestive system.
In general, more detailed investigations on the neural regulation of the digestive system are needed. It is suggested that there is a stomatogastric nervous system (STNS) which includes the STG+CoG+OG, however the interrelationships are not yet fully described.
There also remain many unanswered questions concerning the connections between neural centers and neuroendocrine organs, especially the pericardial organs. A variety of substances are able to be released into the hemolymph but what drives the release under various conditions are only beginning to be investigated. The neuroendocrine organs likely receive commands from nerve centers but it is not known if only one or more than one center is involved. There is likely a very sophisticated control of the neuroendocrine organs which may well be the most important function of the putative ANS in crustaceans.
Preliminary versions of the paper were presented at meeting conferences (Cooper et al., 2002; Shuranova et al., 2000, 2001)
ACKNOWLEDGMENTS
The work was partly sponsored by the RFBR grant No. 05-04-48610 (Z.S.) and NSF-IBN-0131459 (RLC).
List of Abbreviations| • | ANS - Autonomic nervous system |
| • | CG - Cardiac ganglion |
| • | CNS - Central nervous system |
| • | CoG - Commissural ganglion |
| • | CPG - Central pattern generator |
| • | DA - Dopamine |
| • | ESG - Electroscaphognathite-gram |
| • | GABA - Gamma-aminobutyric acid |
| • | HR - Heart rate |
| • | 5-HT - 5-hydroxytryptamine (serotonin) |
| • | NA - Noradrenalin |
| • | OA - Octopamine |
| • | OG - Oesophageal ganglion |
| • | SG - Scaphognathite |
| • | STG - Stomatogastric ganglion |
| • | SOG - Suboesophageal ganglion |
| • | VPG - Ventilatory pattern generator |
REFERENCES
- Airriess, C. and B. Mcmahon, 1996. Short-term emersion affects cardiac function and regional haemolymph distribution in the crab cancer magister. J. Exp. Biol., 199: 569-578.
Direct Link - Ayali, A. and R.M. Harris-Warrick, 1999. Monoamine control of the pacemaker kernel and cycle frequency in the lobster pyloric network. J. Neurosci., 19: 6712-6722.
Direct Link - Beltz, B.S. and E.A. Kravitz, Beltz, B.S. and E.A. Kravitz, 1983. Mapping of serotonin-like immunoreactivity in the lobster nervous system. J. Neurosci., 3: 585-602.
Direct Link - Beltz, B.S., S.M. Helluy, M.L. Ruchhoeft and L.S. Gammill, 1992. Aspects of the embryology and neural development of the American lobster. J. Exp. Zool., 261: 288-297.
Direct Link - Blitz, D.M. and M.P. Nusbaum, 1999. Distinct functions for cotransmitters mediating motor pattern selection. J. Neurosci., 19: 6774-6783.
Direct Link - Bohm, H., E. Messai and H.G. Heinzel, 1997. Activity of command fibres in free-ranging crayfish, Orconectes limosus. Raf. Naturwissenschaften, 84: 408-410.
CrossRef - Birikundavyi, S., R. Labib, H.T. Trung and J. Rousselle, 2002. Performance of neural networks in daily stream flow forecasting. J. Hydrol. Eng., 7: 392-398.
Direct Link - Bojsen, B.H., H. Witthofft, B. Styrishave and O. Andersen, 1998. In situ studies on heart rate and locomotor activity in the freshwater crayfish Astacus astacus (L.) in relation to natural fluctuations in temperature and light intensity. Freshwater Biol., 39: 455-465.
Direct Link - Breen, C.A. and H.L. Atwood, 1983. Octopamine a neurohormone with presynaptic activity-dependent effects at crayfish neuromuscular junctions. Nature, 303: 716-718.
CrossRef - Bustamante, J. and F.B. Krasne, 1991. Effects of octopamine on transmission at the first synapse of the crayfish lateral giant escape reaction pathway. J. Comp. Physiol. A, 169: 369-377.
Direct Link - Castan!on-Cervantes, O., B.A. Batelle and M.L. Fanjul-Moles, 1999. Rhythmic changes in the serotonin content of the brain and eyestalk of the crayfish during development. J. Exp. Biol., 202: 2823-2830.
Direct Link - Christie, A.E., P. Skiebe and E. Marder, 1995. Matrix of neuromodulators in neurosecretory structures of the crab Cancer borealis. J. Exp. Biol., 198: 2431-2439.
Direct Link - Combes, D., P. Meyrand and J. Simmers, 1999. Motor pattern specification by dual descending pathways to a lobster rhythm-generating network. J. Neurosci., 19: 3610-3619.
Direct Link - Cooke, I.M., 1966. The sites of action of pericardial organ extract and 5-hydroxytryptamine in the decapod crustacean heart. Am. Zool., 6: 107-121.
CrossRef - Cooke, I.M. and D.K. Hartline, 1975. Neurohormonal alteration of integrative properties of the cardiac ganglion of the lobster Homarus americanus. J. Exp. Biol., 63: 33-52.
Direct Link - Cooke, I.M., 1988. Studies on the crustacean cardiac ganglion. Comp. Biochem. Physiol., 91: 205-218.
Direct Link - Cooper, R.L., A. Donmezer and J. Shearer, 2003. Intrinsic differences in sensitivity to 5-HT between high- and low-output terminals innervating the same target. Neurosci. Res., 45: 163-172.
CrossRefDirect Link - Cooper, R.L., E. Ward, R. Braxton, H. Li and W.M. Warren, 2003. The effects of serotonin and ecdysone on primary sensory neurons in crayfish. Microsc. Res. Tech., 60: 336-345.
Direct Link - Copenhaver, P.F. and P.H. Taghert, 1989. Development of the enteric nervous system in the moth. II. Stereotyped cell migration precedes the differentiation of embryonic neurons. Dev. Biol., 131: 85-101.
Direct Link - Cuadras, J., 1979. Heart rate and agonistic behavior in unrestrained crabs. Mar. Behav. Physiol., 6: 189-196.
Direct Link - Delaney, K., D.W. Tank and R.S. Zucker, 1991. Presynaptic calcium and serotonin-mediated enhancement of transmitter release at crayfish neuromuscular junction. J. Neurosci., 11: 2631-2643.
Direct Link - DiCaprio, R.A., 1989. Nonspiking interneurons in the ventilatory central pattern generator of the shore crab Carcinus maenas. J. Comp. Neurol., 285: 83-106.
Direct Link - Dircksen, H., P. Skiebe, B. Abel, H. Agricola, K. Buchner, J.E. Muren and D.R. Nassel, 1999. Structure distribution and biological activity of novel members of the allatostatin family in the crayfish Orconectes limosus. Peptides, 20: 695-712.
Direct Link - Dixon, D. and H.L. Atwood, 1989. Adenyate cyclase system is essential for long-term facilitation at the crayfish neuromuscular junction. J. Neurosci., 9: 4246-4252.
Direct Link - Dropic, A.J., E. Brailoiu, and R.L. Cooper, 2005. Presynaptic mechanism of action induced by 5-HT in nerve terminals possible involvement of ryanodine and IP3 sensitive Ca2+ stores. Comp. Biochem. Physiol. A, 142: 355-361.
Direct Link - Elekes, K., E. Florey and M.A. Cahill, 1988. Morphology and central synaptic connections of the efferent neurons innervating the crayfish hindgut. Cell Tissue Res., 254: 369-379.
CrossRef - Evans, P.D., E.A. Kravtiz and B.R. Talamo, 1976. Octopamine release at two points along lobster nerve trunks. J. Physiol., 262: 71-89.
Direct Link - Field, L.H. and J.L. Larimer, 1975. The cardioregulatory system of crayfish neuroanatomy and physiology. J. Exp. Biol., 62: 519-530.
Direct Link - Flamm, R.E. and R.M. Harris-Warrick, 1986. Aminergic modulation in lobster stomatogastric ganglion. 2. Target neurons of dopamine, octopamine and serotonin within the pyloric. J. Neurophysiol., 55: 866-881.
Direct Link - Florey, R.E., 1954. An inhibitory and an excitatory factor of mammalian central nervous system and their action on a single sensory neuron. Arch. Intern. Physiol., 62: 33-53.
CrossRef - Florey, R.E., 1960. Studies on the nervous regulation of the heart beat in decapod crustacea. J. Gen. Physiol., 43: 1061-1081.
PubMed - Ganfornina, M.D., D. Sanchez and M.J. Bastiani, 1996. Embryonic development of the enteric nervous system of the grasshopper Schistocerca americana. J. Comp. Neurol., 372: 581-596.
Direct Link - Glanzman, D.L. and F.B. Krasne, 1983. Serotonin and octopamine have opposite modulatory effects on the crayfish's lateral giant escape reaction. J. Neurosci., 3: 2263-2269.
Direct Link - Gribble, N.A., 1994. A conceptual model for the regulation of cardiac and ventilatory activity of the portunid crab Portunus pelagicus Linnaeus. J. Theor. Biol., 168: 19-29.
Direct Link - Guirguis, M.S. and J.L. Wilkens, 1995. The role of the cardioregulatory nerves in mediating heart rate responses to locomotion, reduced stroke volume and neurohormones in Homarus americanus. Biol. Bull., 188: 179-185.
Direct Link - Harris-Warrick, R.M., B.R. Johnson, J.H. Peck, P. Kloppenburg, A. Ayali and J. Skarbinski, 1998. Distributed effects of dopamine modulation in the crustacean pyloric network. Ann. N. Y. Acad. Sci., 860: 155-167.
Direct Link - Hartenstein, V., 1997. Development of the insect stomatogastric nervous system. Trends Neurosci., 20: 421-427.
Direct Link - Hartline, D.K. and D.M. Maynard, 1975. Motor patterns in the stomatogastric ganglion of the lobster Panulirus argus. J. Exp. Biol., 62: 405-420.
Direct Link - Winlow, W. and M.S. Laverack, 1972. The control of hindgut motility in the lobsterHomarus gammarus (L.). I. Analysis of hingut movements and receptor activity. Mar. Behav. Physiol., 1: 1-27.
PubMed - Howard, D.R. and P. Talbot, 1992. In vitro contraction of lobster Homarus ovarian muscle methods for assaying contraction and effects of biogenic amines. J. Exp. Zool., 263: 356-366.
Direct Link - Johnson, B.R. and R.M. Harris-Warrick, 1997. Amine modulation of glutamate responses from pyloric motor neurons in lobster stomatogastric ganglion. J. Neurophysiol., 78: 3210-3221.
Direct Link - Kellie, S., J. Greer and R.L. Cooper, 2001. Alterations in habituation of the tail flip response in epigean and troglobitic crayfish. J. Exp. Zool., 290: 163-176.
CrossRefDirect Link - King, D.G., 1976. Organization of crustacean neuropil. I. Patterns of synaptic connections in lobster stomatogastric ganglion. J. Neurocytol., 5: 207-237.
Direct Link - Kulkarni, G.K. and M. Fingerman, 1992. Quantitative analysis by reverse phase high performance liquid chromatography of 5-hydroxytryptamine in the central nervous system of the red swamp crayfish, Procambarus clarki. Biol. Bull., 182: 341-347.
Direct Link - Langworthy, K., S. Helluy, J. Benton and B. Beltz, 1997. Amines and peptides in the brain of the American lobster: Immunocytochemical localization patterns and implications for brain function. Cell Tissue Res., 288: 191-206.
CrossRef - Li, H., L.R. Listerman, D. Doshi and R.L. Cooper, 2000. Heart rate measures in blind cave crayfish during environmental disturbances and social interactions. Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 127: 55-70.
CrossRefDirect Link - Listerman, L.R., J. Deskins, H. Bradacs and R.L. Cooper, 2000. Heart rate within male crayfish social interactions and effects of 5-HT. Comp. Biochem. Physiol. A Mol. Integr. Physiol., 125: 255-263.
PubMed - McMahon, B.R., 2001. Control of cardiovascular function and its evolution in Crustacea. J. Exp. Biol., 204: 923-932.
Direct Link - Mercier, A.J., I. Orchard and V. TeBrugge, 1991. Fmrfamide-like immunoreactivity in the crayfish nervous system. J. Exp. Biol., 156: 519-538.
Direct Link - Page, M.P. and R.L. Cooper, 2004. Novelty stress and reproductive state alters responsiveness to sensory stimuli and 5-HT neuromodulation. Comp. Biochem. Physiol. A, 139: 149-158.
Direct Link - Prosser, C.L., R.A. Nystrom and T. Nagai, 1965. Electrical and mechanical activity in intestinal muscles of several invertebrate animals. Comp. Biochem. Physiol., 14: 53-70.
Direct Link - Rajashekar, K.P. and J.L. Wilkens, 1992. Dopamine and nicotine but not serotonin modulate the crustacean ventilatory pattern generator. J. Neurobiol., 23: 680-691.
Direct Link - Reiber, C., B. McMahon and W. Burggren, 1997. Cardiovascular functions in two macruran decapod crustaceans Procambarus clarkii and Homarus americanus during periods of inactivity, tail flexion and cardiorespiratory pauses. J. Exp. Biol., 200: 1103-1113.
Direct Link - Rose R.A., J.L. Wilkens and R.L. Walker, 1998. The effects of walking on heart rate ventilation rate and acid-base status in the lobster Homarus americanus. J. Exp. Biol., 201: 2601-2608.
Direct Link - Rose, R.A., K. MacDougall, A. Patel, J.L. Wilkens and R.L. Walker, 2001. Effects of walking on ventilatory and cardiac function in intact and cardiac impaired lobsters. Physiol. Biochem. Zool., 74: 102-110.
Direct Link - Russell, D.F., 1979. CNS control of pattern generators in the lobster stomatogastric ganglion. Brain Res., 177: 598-602.
Direct Link - Schapker, H., T. Breithaupt, Z. Shuranova, Y. Burmistrov and R.L. Cooper, 2002. Heart rate and ventilatory correlative measures in crayfish during environmental disturbances and social interactions. Comp. Biochem. Physiol. A, 131: 397-407.
CrossRef - Schneider, H., B.A. Trimmer, J. Rapus, M. Eckert, D.E. Valentine and E.A. Kravitz, 1993. Mapping of octopamine-immunoreactive neurons in the central nervous system of the lobster. J. Comp. Neurol., 329: 129-142.
Direct Link - Selverston, A.I., D.F. Russell and J.P. Miller, 1976. The stomatogastric nervous system structure and function of a small neural network. Prog. Neurobiol., 7: 215-290.
Direct Link - Selverston, A.I., R. Elson, M. Rabinovich, R. Huerta and H. Abarbanel, 1998. Basic principles for generating motor output in the stomatogastric ganglion. Ann. N. Y. Acad. Sci., 860: 35-50.
Direct Link - Shuranova, Z.P., Y.M. Burmistrov and R.L. Cooper, 2003. Bioelectric field potentials of the ventilatory muscles in the crayfish. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 134: 461-469.
CrossRefDirect Link - Simmers, A.J. and B.M.H. Bush, 1983. Central nervous mechanisms controlling rhythmic burst generation in the ventilatory motoneurones of Carcinus maenas. J. Comp. Physiol., 150: 1-21.
CrossRef - Simmers, J., P. Meyrand and M. Moulins, 1995. Modulation and dynamic specification of motor rhythm-generating circuits in Crustacea. J. Physiol. Paris, 89: 195-208.
CrossRef - Siwicki, K.K., B.S. Beltz and E.A. Kravitz, 1987. Proctolin in identified serotonergic dopaminergic and cholinergic neurons in the lobster Homarus americanus. J. Neurosci., 7: 522-532.
Direct Link - Southard, R.C., J. Haggard, M.E. Crider, S.W. Whitheart and R.L. Cooper, 2000. Influence of serotonin on the kinetics of vesicular release. Brain Res., 871: 16-28.
CrossRef - Strawn, J.R., W.S. Neckameyer and R.L. Cooper, 2000. The effects of 5-HT on sensory, central and motor neurons driving the abdominal superficial flexor muscles in the crayfish. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 127: 533-550.
PubMedDirect Link - Swensen, A.M. and E. Marder, 2000. Multiple peptides converge to activate the same voltage-dependent current in a central pattern-generating circuit. J. Neurosci., 20: 6752-6759.
Direct Link - Taylor, E.W., 1970. Spontaneous activity in the cardioaccelerator nerves of the crayfish Astacus pallipes Lereboullet. Comp. Biochem. Physiol., 33: 859-869.
Direct Link - Taylor, E.W., 1982. Control and co-ordination of ventilation and circulation in crustaceans responses to hypoxia and exercise. J. Exp. Biol., 100: 289-319.
Direct Link - Tierney, A.J., J. Blanck and A.J. Mercier, 1997. Fmrfamide-like peptides in the crayfish Procambarus clarkii stomatogastric nervous system distribution and effects on the pyloric motor pattern. J. Exp. Biol., 200: 3221-3233.
Direct Link - Vu, E.T. and F.B. Krasne, 1992. Evidence for a computational distinction between proximal and distal neuronal inhibition. Science, 255: 1710-1712.
Direct Link - Weiger, W.A., 1997. Serotonergic modulation of behaviour: A phylogenetic overview. Biol. Rev., 72: 61-95.
Direct Link - Wiersma, C.A. and E. Novitski, 1942. The mechanism of nervous regulation of the crayfish heart. J. Exp. Biol., 19: 255-265.
Direct Link - Wilkens, J.L., A.J. Mercier and J. Evans, 1985. Cardiac and ventilatory responses to stress and to neurohormonal modulators by the shore crab Carcinus maenas. Comp. Biochem. Physiol. C, 82: 337-343.
CrossRef - Wilkens, J.L., R.E. Young and R.A. DiCaprio, 1989. Responses of the isolated crab ventilatory central pattern generators to variations in oxygen tension. J. Comp. Physiol. B, 159: 29-36.
CrossRef - Wilkens, J.L., 1995. Regulation of the cardiovascular system in crayfish. Am. Zool. 35: 37-48.
CrossRef - Wilkens, J.L., 1999. The control of cardiac rhythmicity and of blood distribution in crustaceans. Comp. Biochem. Physiol. A, 124: 531-538.
CrossRef - Yamagishi, H., H. Ando and T. Makioka, 1997. Myogenic heartbeat in the primitive crustacean Triops longicaudatus. Biol. Bull., 193: 350-358.
Direct Link - Yazawa, T. and K. Kuwasawa, 1992. Intrinsic and extrinsic neural and neurohumoral control of the decapod heart. Cell. Mol. Life Sci., 48: 834-839.
CrossRef - Yeh, S.R., R.A. Fricke and D.H. Edwards, 1996. The effect of social experience on serotonergic modulation of the escape circuit of crayfish. Science, 271: 366-369.
Direct Link - Harris-Warrick, R.M., M. Nusbaum and F. Nagy, 1992. Modulation of Stomatogastric Neural Networks by Identified Transmitters and Neurons. The Stomatogastric Nervous System: A Model Biological Neural Network, Harris-Warrick, R.M., E. Marder, A.I. Selverston and M. Moulins (Eds.). MIT Press, Boston,.
CrossRef