
R. G. Ahmed
Laboratory of Cognitive and Brain Research, Kunmmg Institute of Zoology,
The Chinese Academy of Sciences, Kunming, Yunnan 650223, Peoples Republic of China
Yuan Ye Ma
Laboratory of Cognitive and Brain Research, Kunmmg Institute of Zoology,
The Chinese Academy of Sciences, Kunming, Yunnan 650223, Peoples Republic of China
S. H. Lee
Department of Reproductive Animals, Korea University, South Korea
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 226-241







ABSTRACT
Peroxiredoxins (Prxs) are a family of novel antioxidant proteins that are found in a variety of species and participate in a number of vital biological processes such as proliferation, differentiation, response to oxidative stress and intracellular signaling. It has been proposed that they might participate in these cellular processes by playing a role in eliminating or regulating the intracellular concentration of peroxides produced during metabolism as well as in the signaling cascades of growth factors and cytokines. Mammalian cells express six isoforms of Prx (Prx I to VI), which are classified into three subgroups (typical 2-Cys, atypical 2-Cys and 1-Cys) based on the number and position of cysteine (Cys) residues that participate in catalysis and on amino acid sequences and the immunological reactivity. Members of the typical 2-Cys subgroup include Prx I through Prx IV and contain an additional conserved cysteine in the carboxyl-terminal region, whereas Prx V and Prx VI, members of the atypical 2-Cys and 1-Cys subgroups, respectively, do not contain this second conserved Cys. On the other hand, Prxs activity can be regulated by phosphorylation and proteolysis processes in addition to overoxidation. Taken together, this study suggest that the generation of the oxidative stress which caused neurodegeneration may couple with produced Prxs and the reverse is true. However, this argument is still unclear on account of the difficulties of the direct observation of the reactive oxygen species due to their biological lifetime is short. Thus, experiments will be required to solve these problems and to comprehend the actual role of Prxs in neurodegeneration.
PDF Abstract XML References
How to cite this article
R. G. Ahmed, Yuan Ye Ma and S. H. Lee, 2006. Peroxiredoxins and Neurodegeneration. International Journal of Zoological Research, 2: 226-241.
DOI: 10.3923/ijzr.2006.226.241
URL: https://scialert.net/abstract/?doi=ijzr.2006.226.241
DOI: 10.3923/ijzr.2006.226.241
URL: https://scialert.net/abstract/?doi=ijzr.2006.226.241
INTRODUCTION
Oxidative stress is caused as the result of a variety of physiological and pathophysiological conditions (Matsumoto et al., 1999). Oxidative modifications of biological molecules such as proteins, lipids and nucleic acids are potentially harmful to living organisms and may contribute to aging (Stadtman, 1992). Therefore, thiol groups are highly sensitive to oxidation, proteins in which thiol groups play a key role represent an important target of oxidative stress. Certain antioxidative enzymes such as the peroxiredoxins (Prxs), superoxide dismutases (SODs) and glutathione peroxidases (GPxs) and antioxidant molecules such as glutathione and thioredoxin serve to protect essential molecules against oxidative modification (Matsumoto et al., 1999). Prxs are distinct from other peroxidases in that they have no cofactors, such as metals or prosthetic groups (Hirotsu et al., 1999). Prxs generally have two conserved cysteines (Cys-52 and Cys-173 of Heme-binding protein 23 kDa (HBP23)) at the N- and C-terminal regions (Chae et al., 1994a) and their antioxidant effects are coupled with the physiological electron donor activity of the thioredoxin system (Chae et al., 1994a; Kang et al. 1998a). Moreover, Prxs are a family of novel antioxidant proteins that are found in a variety of species and which has been shown to participate in a number of vital biological processes (Chae et al., 1993; Ishii et al., 1993; Shau et al., 1993; Chae et al., 1994a; Watabe et al., 1994) such as antioxidant defense, redox signaling, apoptosis control and kinase modulation (Isermann et al., 2004).
Numerous studies have demonstrated that Prxs can protect cells against a variety of nitrosative stresses (Peterson and Luckhart, 2005). In addition, brain cells also display higher sensitivity to oxidative stress than cells of other tissues due to the abundance of iron and oxidizable lipids (Halliwell, 1992; Beal, 1996; Castagne et al., 1999). Reactive Oxygen Species (ROS) toxicity is a common underlying feature of neuropathological states, including Pick's Disease (PD), Alzheimer's Disease (AD) and Down Syndrome (DS) and proteins of the Prx gene and other antioxidant gene families have been documented to exert antioxidant and cytoprotective effects (Butterfield et al., 1999). Characterizing the patterns of Prx protein expression in normal and diseased brain could thus yield significant insight into the neurochemical mechanisms of defense and pathology. Furthermore, Prxs provide a valuable tool for elucidating the role of antioxidant pathways in the pathological processes. Therefore, it is imperative to describe the backgrounds and recent findings which provide specific suggestions for explore the relation between peroxiredoxins and neurodegeneration:
Peroxiredoxins
The peroxiredoxins (Prxs) are a recently discovered family of antioxidant peroxidases that can reduce hydrogen peroxide and alkyl hydroperoxides to water and the corresponding alcohols, respectively and can protect cells from widely divergent organisms against a variety of nitrosative stress challenges (Bryk et al., 2000; Wong et al., 2002; Dubuisson et al., 2004; Trujillo et al., 2004). Prxs have also been implicated in oxidative signaling mechanisms regulating apoptosis (Kim et al., 2000), cell differentiation (Rabilloud et al., 1995; Jin et al., 1997; Butterfield et al., 1999). The peroxiredoxins define an emerging family of peroxidases able to reduce hydrogen peroxide and alkyl hydroperoxides with the use of reducing equivalents derived from thiol-containing donor molecules such as thioredoxin, glutathione and trypanothione (Declercq et al., 2001). Prxs are a family of peroxidases widely distributed in eukaryotes and prokaryotes (Plaisant et al., 2003). In higher plants they are involved in balancing hydroperoxide production during photosynthesis, while in higher animals peroxiredoxins appear to be involved in the redox-regulation of cellular signaling and differentiation, displaying in part opposite effects (Hofmann et al., 2002). Indeed, these enzymes are truly ubiquitous having been identified in yeast, plant and animal cells, including both protozoan and helminth parasites and most, if not all, eubacteria and archaea. Although located primarily in the cytosol, Prxs are also found within mitochondria, chloroplasts and peroxisomes, associated with nuclei and membranes and, in at least one case, exported (Jin and Jeang, 2000; Hofmann et al., 2002). Prxs are produced at high levels in cells; in addition, proteins in Escherichia coli (Link et al., 1997), the second or third most abundant protein in erythrocytes (Moore et al., 1991) and compose 0.1-0.8% of the soluble protein in other mammalian cells (Chae et al., 1999). Many organisms produce more than one isoform of Prx, including at least six Prxs identified in mammalian cells (Prx I-Prx VI). Furthermore, the Prxs are an emerging family of multifunctional enzymes that exhibit peroxidase activity in vitro and in vivo participate in a range of cellular processes known to be sensitive to reactive oxygen species (Schroder et al., 2000). An immediate physiological electron donor for the peroxidase catalysis for five Prx proteins (Prx I-V) has been identified as thioredoxin (Trx), but that for Prx VI (1-Cys Prx) is still unclear (Lee et al., 2001). They appear to be fairly promiscuous with respect to the hydroperoxide substrate; the specificities for the donor substrate vary considerably between the subfamilies, comprising GSH, thioredoxin and tryparedoxin (Hofmann et al., 2002).
Crystal structures of Prx I, II, V and VI have revealed that Prxs are novel members of the double-stranded RNA production and conditions for RNA interference (RNAi) (Peterson and Luckhart, 2005). 2-Cys (2-cysteine) peroxiredoxins are a large and diverse family of peroxidases which, in addition to their antioxidant functions, regulate cell signaling pathways, apoptosis and differentiation (Wood et al., 2002). These enzymes are obligate homodimers (alpha(2)), utilizing a unique intermolecular redox-active disulfide center for the reduction of peroxides and are known to form two oligomeric states: individual alpha(2) dimers or doughnut-shaped (alpha(2))(5) decamers. Wood et al. (2002) suggest that the enzymatic and signaling activities of all 2-Cys Prxs are regulated by a redox-sensitive dimer to decamer transition. Plaisant et al. (2003) found that Prx V is a recently discovered mammalian member of this family of antioxidant enzymes that is able to reduce hydrogen peroxide and alkyl hydroperoxides (Plaisant et al., 2003). In doing so, a reactive cysteine in the peroxiredoxin active site is weakly oxidized (disulfide or sulfenic acid) by the destroyed peroxides and the cellular thiols (e.g., thioredoxin) are used to regenerate the peroxiredoxins to their active state (Rabilloud et al., 2002).
Also, Peroxiredoxins constitute a novel family of antioxidant proteins, which specifically prevent enzymes from metal-catalyzed oxidation (Chuchalin et al., 2003). Prx family members in the presence of some thiols can specifically prevent biopolymers (proteins, nucleic acids, lipids) from damage by reactive oxygen species (Chae et al., 1994b). Recently, a novel family of proteins which function as antioxidants, in the protection of thiol groups in proteins, has been reported (Chae and Rhee, 1994). They also exhibit peroxidase activity in a thioredoxin (Trx)-dependent manner, they are referred to as Prxs and constitute a system which is similar to the glutathione/glutathione-peroxidase system (Chae et al., 1994b). Peroxiredoxin exhibits thioredoxin-dependent peroxidase activity and constitutes a family of proteins (Matsumoto et al., 1999). Thioredoxin peroxidase (TPx), one of the Prx gene superfamily, was first reported from Saccharo-myces cerevisiae (Kim et al., 1988 and 1989). TPx functions as an antioxidant to remove the reactive oxygen species (ROS) O2¯ and H2O2 (Docampo, 1995) derived from normal cellular metabolism (Gutteridge and Halliwell, 1989) using thioredoxin as the electron donor (Henkle-Duhrsen and Kampkotter, 2001). High concentrations of ROS can have serious deleterious effects on membrane lipids, nucleic acids and proteins.
Prxs exert their protective antioxidant role in cells through their peroxidase activity (Wood et al., 2003):
whereby hydrogen peroxide, peroxynitrite and a wide range of organic hydroperoxides (ROOH) are reduced and detoxified (Jacobson et al., 1989; Poole and Ellis, 1996; Peshenko and Shichi, 2001; Hofmann et al., 2002).
Mammalian Prxs and Their Reaction Mechanisms
The Prx antioxidant protein family is found in a wide variety of species and plays important roles in protecting cells against oxidants and in regulating signaling by hydrogen peroxide (Yanagawa et al., 2005). Also, the role of Prx in protection from hydroxyl radicals (Lim et al., 1993), thiyl radicals and oxidized thiyl radical anions (Yim et al., 1994) in the in vitro system is due at least in part to its peroxidase activity (Netto et al., 1996). The peroxiredoxin protein is expressed widely in animal tissues and serves an antioxidant function associated with removal of cellular peroxides (Sarafian et al., 1999). At least six Prx enzymes are present in mammals (Prx I-VI) (Rhee et al., 2001). However, Chae et al. (1999) recorded that twelve mammalian Prx members have been previously identified in association with various cellular functions apparently unrelated to peroxidase activity. Also, Prxs are a recently described superfamily of nonseleno-peroxidases that catalyze the reduction of a broad spectrum of peroxides (Wang et al., 2004). These Prx iso-forms are found in the cytosol, mitochondria, peroxisomes and plasma, all of which are potential sites of ROS production (Kang et al., 1998b; Matsumoto et al., 1999; Mizusawa et al., 2000; Oberley et al., 2001; Fujii and Ikeda, 2002).
The mammalian Prxs family contain six different isoforms that are divided into three subtypes according; 1) The number of cysteine residues that are directly involved in catalysis and peroxidase reactions: 2-Cys (Prx I-IV), atypical 2-cys (Prx V) and 1-Cys (Prx VI) (Wood et al., 2003); 2) The amino acid sequences (Jin et al., 1997); and 3.) The immunological reactivity (Jin et al., 1997; Kang et al., 1998a). Some Prxs possess reductase activity for hydrogen peroxide and alkyl hydroperoxides depend on; 1) Their ability to reduce these substances to H2O2 or alcohols, respectively (Bruchhaus et al., 1997) 2) Their involvement in protection against phospholipid peroxidation (Manevich et al., 2002). Moreover, Prxs act as peroxynitrite reductases and participate in the detoxification of reactive nitrogen species (Bryk et al., 2000). The Prx isoforms share a common reactive cysteine residue in their N-terminal region, which is oxidized to either cysteine sulfinic acid or disulfide, both of which are then readily reduced back to the sulfhydryl state by various cellular reductants (Rhee et al., 2005b). Wood et al. (2003) reported that the peroxidase reaction is composed of two steps centered around a redox-active cysteine called the peroxidatic cysteine. All Prxs to date conserve an active-site Arg, which would lower the pKa of the peroxidatic cysteine somewhat by stabilizing its thiolate form. The second step of the peroxidase reaction, the resolution of the cysteine sulfenic acid, distinguishes the three Prx classes (Fig. 1). Thus, these Prxs use redox-active cysteines to reduce peroxides (Chae et al., 1994a).
| Mammalian cells express six Prx isoforms (Prx I-VI), which can be divided into three subgroups according the number and position of the active Cys-Prxs residues as a following: |
 |
 |
 |
Regulation of Prxs Activity
Prxs have received a great deal of attention recently owing to their role in regulating levels of hydrogen peroxide, an intracellular signaling molecule common to many cytokine-induced signal-transduction pathways (Jin et al., 2000; Hofmann et al., 2002). Indeed, regulation of redox signaling through cysteine modification by peroxides and peroxynitrite has been reported for a growing number of enzymes and transcriptional regulators (Claiborne et al., 1999). It was recently shown that the overoxidation of Prx II is likely to be physiologically relevant, in that its peroxidatic cysteine is oxidized to sulfinic (-SO3H) or sulfonic (-SO2H) acid forms in vivo upon exposure of Leydig cells to tumor necrosis factor (Rabilloud et al., 2002). It has been proposed that Prxs in mammalian cells act as a dam against oxidative stress and that the ratio of active to inactive enzyme might play a role in whether cells are susceptible to cytokine-induced apoptosis (Rabilloud et al., 2002). In addition to overoxidation, Prx activity has also been shown to be regulated by phosphorylation and proteolysis (Schroder et al., 1998; Cha et al., 2000; Chang et al., 2002; Koo et al., 2002).
Phosphorylation Process
Recently, phosphorylation of mammalian Prx I, Prx II, Prx III and Prx IV at the conserved residue Thr89 (PrxTT numbering) by cyclin-dependent kinases was shown to decrease the peroxidase activity of the Prxs (Chang et al., 2002). In the case of Prx I, this phosphorylation was observed to occur in vivo during mitosis. The authors concluded that the phosphorylated Thr89 had an unfavorable electrostatic effect on the peroxidatic active site. An examination of the structure of the Prx II decamer reveals that a phosphorylated Thr89 would introduce unfavorable electrostatic interactions within the dimer-dimer interface by placing two negatively charged phosphates in close proximity (Wood et al., 2003). In fact, a reasonable alternative interpretation is that phosphorylation of Thr89 attenuates the enzyme activity by disrupting the decameric structure (Wood et al., 2003).
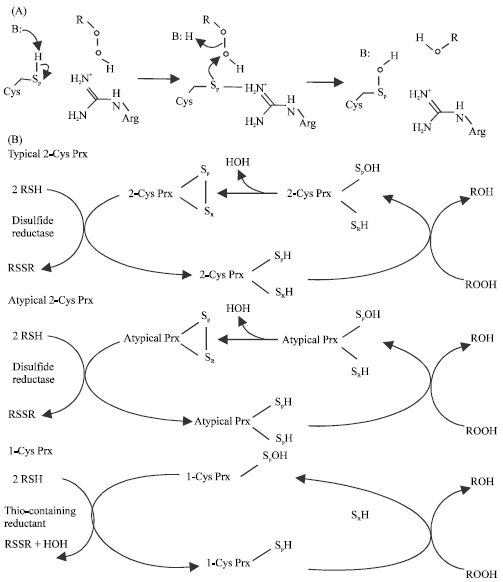 | |
| Fig. 1: | Peroxiredoxin (Prx) mechanism (Wood et al., 2003). (a) The common first step of peroxide reduction involving nucleophilic attack by the peroxidatic cysteine (Sp) and formation of the cysteine sulfenic acid intermediate (SpOH), probably shared by all Prxs. Both the catalytic base that deprotonates the peroxidatic cysteine and catalytic acid that protonates the RO¯ leaving group are labeled B, although this does not imply that they are the necessarily same entity. The guanidino group of the conserved arginine is presumed to stabilize the ionized peroxidatic cysteine. (b) The three mechanisms distinguishing the Prx classes, with peroxidatic cysteine and resolving cysteines in the reduced (SpH and SRH, respectively), sulfenic acid (SpOH) or disulfide (Sp and SR, connected) state. In the case of dimeric 2-Cys Prxs, the peroxidatic cysteine (black Sp) and resolving cysteine (pink SR) originate from different subunits and condense to form an intersubunit disulfide bond (black and pink striped bar). Reduction of typical and atypical 2-Cys Prxs involves one flavoprotein disulfide reductase and at least one additional protein or domain containing a CXXC motif, which is oxidized from a dithiol (2 RSH) to a disulfide (RSSR) state during Prx reduction (e.g., thioredoxin reductase and thioredoxin, AhpF, trypanothione reductase, trypanothione and tryparedoxin, or lipoamide dehydrogenase, SucB and AhpD (Nogoceke et al., 1997; Poole et al., 2000 and Bryk et al., 2002). Reductants of 1-Cys Prxs include low molecular weight thiols, but physiological partners are as-yet unidentified. |
Several researchers have reported that dimeric forms of Prxs exhibit less activity than decameric forms (Nogoceke et al., 1997; Chauhan and Mande, 2001). This observation is supported by the crystal structures, which show that the active sites of the typical 2-Cys Prxs are adjacent to and stabilized by the dimmer-dimer interface of the decamer. It is notable that the phosphorylation mechanism probably favors different oligomeric states (dimer and decamer, respectively).
Like many enzymes involved in the metabolism of intracellular messengers, the peroxidase activity of Prx I and Prx II is regulated by phosphorylation, which in this instance is mediated by cyclin-dependent kinases (Cdks) (Chang et al., 2002). Phosphorylation at Thr90 by several Cdks, including Cdc2, in vitro, results in inhibition of its peroxidase activity. Experiments with HeLa cells arrested at various stages of the cell cycle showed that phosphorylation of Prx I on Thr90 occurs in parallel with the activation of Cdc2 (Rhee et al., 2005a). Prx I phosphorylation was observed in cells in the M phase but not in interphase cells, despite the fact that Prx I can be phosphorylated by other Cdks in vitro. This specificity likely reflects the facts that Prx I is a cytosolic protein and therefore only encounters activated Cdks after the breakdown of the nuclear envelope during mitosis and that Cdc2 is the Cdk that is activated in the M phase (Rhee et al., 2005a). These results suggest that Cdc2-dependent phosphorylation of Prx I/Prx II and the resulting accumulation of H2O2 might be a critical step in the progression of normal cell division.
Proteolysis Process
Another mechanism proposed to regulate peroxidase activity in vivo entails specific proteolysis of the C termini of Prxs, preventing peroxide-mediated inactivation in response to rising levels of peroxide (Koo et al., 2002). In studies of a typical 2-Cys Prx from yeast, a portion of the enzyme was found to have a truncated C-terminal following purification (Koo et al., 2002). In follow-up mutagenesis studies, C-terminally truncated forms of the enzyme were found to be more resistant to peroxide overoxidation and inactivation than the sensitive wild-type enzyme (Koo et al., 2002). A similar truncation of Prx II that removed the C-terminal 13 residues (including the last a helix) has also been observed during the isolation of the enzyme from erythrocytes (Cha et al., 2000). Interestingly, the regulatory protease calpain is present in erythrocytes and will specifically cleave this region of Prx II in vitro (Schroder et al., 1998). Proteolysis would make the enzyme resistant to overoxidation but leave it susceptible to inactivation by phosphorylation.
Neurodegeneration
The inherent biochemical and physiological characteristics of the brain, with its high unsaturated phospholipid content and energy requirements, make it particularly susceptible to free radical mediated injury (Krapfenbauer et al., 2003). Furthermore, the brain has the ability to readily accumulate iron, which promotes the production of the damaging hydroxyl radical and is relatively poorly endowed with protective antioxidant enzymes or antioxidant compounds (Halliwell and Gutteridge, 1999). The reactive oxygen intermediates (including superoxide and hydroxyl radicals as well as hydrogen peroxide) can cause direct cellular injury by including lipid and protein peroxidation and damage to nucleic acid (Takeda et al., 1984; Richard et al., 1990). Moreover, Betteridge (2000) reported that the free radicals can be produced by several different biochemical processes within the body including: (1) reduction of the molecular oxygen during aerobic respiration yielding superoxide and hydroxyl radicals; (2) by products of chemical reactions such as oxidation of catecholamme and activation of the arachidonic acid cascade product electrons, which can reduce molecular oxygen to superoxide; (3) production of superoxide and hypochlorous acid (HOC1), a powerful oxidant, by activated phagocytes and (4) nitric oxide production by vascular endothelium and other cells. ROS can cause widespread damage to biological macromolecules and are believed to play a causal role in many degenerative diseases (Demple and Amabile-Cuevas, 1991; Neumann et al., 2003). Lipid peroxidation was previously reported in other neurodegenerative diseases, including Alzheimer's disease and epilepsy (Halliwell and Gutteridge, 1999). Also, Oxidative stress induced by Reactive Oxygen Species (ROS) such as hydrogen peroxide has been implicated in the pathogenesis of several neurodegenerative diseases (Kim et al., 2001a). Thus, the oxidative stress is one of the most important factors in the pathogenesis of idiopathic Parkinson’s disease (Beal, 2002; Mouradian, 2002).
On the other hand, Sarafian et al. (1999) revealed a clear segregation of expression of Prx I and Prx II gene products in different brain cell types. In the cerebral cortex, cerebellum, basal ganglia, substantia nigra and spinal cord, Prx I was expressed primarily in astrocytes, while Prx II was expressed exclusively in neurons. Prx I was also prominently expressed in ependymal cells and subependymal matrix of substantia nigra and basal ganglia. Prx II was not expressed at uniform density in all neurons. In general, small neurons such as cerebellar granule neurons displayed little or no staining, while large neurons, such as hippocampal pyramidal and Purkinje neurons were heavily stained (Sarafian et al., 1999). The absence of expression of Prx I in neurons and the selective expression of Prx II in large neurons suggest that these antioxidant enzymes serve distinct functional roles that may reflect the different functions and biochemical activities of these cell types. Restricted expression of these genes may also contribute to the selective vulnerability of these cells to a wide variety of neuropathologic conditions. In addition, increased protein levels of Prx I and Prx II could provide protection against neuronal cell death induced by hydrogen peroxide, while decreased protein levels of Prx III could be caused by mitochondrial damage shown in Alzheimer's disease (AD) and Down syndrome (DS) (Kim et al., 2001a). Thus, upregulated Prx protein levels provides evidence for the involvement of ROS in the pathogenesis of AD and DS. Hence, increased expression of Prx II in DS, AD and PD frontal cortex could represent a cellular response initiated against apoptosis, as enhanced apoptosis is demonstrated in these disorders (Engidawork et al., 2001; Gulesserian et al., 2001b). In addition, Prx I reduces hydrogen peroxide, the widespread glial expression of Prx I indicates that it may play an important protective role against oxidative damage in the nervous system (Mizusawa et al., 2000).
On the other word, in human brain, Prx I was primarily expressed in astrocytes, whereas Prx II was expressed exclusively in neurons (Sarafian et al., 1999). Prx I, II and VI are localized predominantly to the cytosol, while Prx III and V are largely detected in the organdies (Seo et al., 2000) and Prx IV is secreted outside of the cells (Okado-Matsumoto et al., 2000). Normal aging is associated with increased oxidative stress, which in turn increases the probability of age-dependent neurodegenerative brain pathology (Squire, 2001). Oxidative stress-associated cell damage has also been described in age-related neurodegenerative disorders, including Pick's Disease (PD), Alzheimer's disease (AD) and Down syndrome (DS), as DS has a neurodegenerative component later in life (Lubec, 1996; Multhaup et al., 1997). The region-dependent differences in expression of antioxidant proteins between DS and AD brain were reported previously (Gulesserian et al., 2001c), explaining, at least in part, the variation observed in morphology and distribution of lesions (Mann, 1997). Moreover, abnormal phosphorylation of proteins, a shared characteristic feature of DS, AD and PD, is viewed as a consequence of oxidative stress (Gerst et al., 1999). It appears therefore likely that oxidative stress would contribute to the development of neurodegenerative pathologies in these disorders.
Previously, Prx II is exclusively expressed in neurons (Sarafian et al., 1999) and neuronal loss, in which apoptosis is implicated as a likely mechanism (Caims, 1999; Dickson, 2001), is one characteristic feature of DS, AD and PD. Taking this into account, one would expect decreased expression of Prx II. One possible explanation for this paradox could be that a compensatory mechanism might be initiated by surviving neurons to protect themselves from apoptosis. Oxidative stress in fetal DS is proposed to be linked to decreased expression of Prx proteins (Gulesserian et al., 2001a). The switch in direction of change in adult life with disease progression most likely supports that such compensatory response is operative. This indeed can be taken as evidence for the possible existence of a neuronal apoptosis decision cascade in neurodegenerative disorders in which a competitive decision making process balances cell death mechanisms against protective and reparative processes (Cotman, 1998). Prx II may also be attributed to difference in regional sensitivity to apoptosis. Indeed, the previous studies in DS and AD (Engidawork et al., 2001; Gulesserian et al., 2001b) suggest that cerebellum is less vulnerable to apoptosis than frontal cortex. Although over-expression of Prx I is also shown to inhibit apoptosis, lipid peroxidation and the release of cytochrome c from mitochondria in different cells (Kim et al., 2000), no detectable changes in expression of Prx I were observed in both regions between the experimental subjects. This lends support for the notion that compensatory response initiated by neurons indeed accounts for the change in Prx II, as Prx I is predominantly expressed in glial cells (Sarafian et al., 1999).
Prx III is mainly found in the mitochondria (Krapfenbauer et al., 2003). Since there is extensive damage to mitochondria in neurodegenerative disorders (Kim et al., 2001b), leakage could be an important factor in the disease groups but not in controls. Decreased Prx III levels in the cytosol of DS and PD as well as AD (though insignificant) patients despite leakage suggest that all the mitochondrial Prx III is used up and what was detected is nascent Prx III (Krapfenbauer et al., 2003). Alternatively, Prx III detected in the cytosol could represent both nascent and leaked Prx III. In both cases, decreased Prx III could indicate that there is impaired synthesis of Prx III and/or enhanced Prx III instability in neurodegenerative diseases, this being more severe in DS and PD.
Prx VI has several unique characteristics, including the absence of a second cysteine residue that is conserved in all other Prxs, the presence of a unique carboxy-terminal domain and a demonstrated phospholipase activity (Krapfenbauer et al., 2003). Furthermore, Prx VI shows conservation of several amino acids important in dimer formation and active site configuration that are not found in other family members (Phelan, 1999). Recently, definitive evidence has been provided for the presence of both peroxidase and phospholipase activities in Prx VI (Chen et al., 2000). Accordingly, the enzyme can reduce hydrogen peroxide and short chain organic, fatty acid and phospholipid hydroperoxides. Prx VI expression did not show any apparent change in DS and AD, reflecting the protein may not have significant contribution to cellular defense against oxidative stress in the context of DS and AD. However, its elevation in PD may suggest that Prx VI could be useful in discriminating PD from DS/AD. Oxidative signals are shown to stimulate expression of regulatory factors that bind to stress response element of the Prx VI gene, thereby activating its expression (Fatma, et al., 2001). Increased expression of Prx VI in PD might therefore be caused by certain transcription factor and/or activating factor that is selectively upregulated in PD. Prx VI protects from ROS-mediated DNA fragmentation (Fatma, et al., 2001), which results in neuronal death by apoptosis or other mechanism (Gleckman et al., 1999). Thus, upregulation of Prx VI might be linked to greater degree of neuronal loss in PD (Dickson, 2001) although the technique of proteomics does not distinguish the cell (neuron/glia) that elaborates Prx VI. Generally, the oxidative stress and damage play a role in the pathogenesis of a number of diseases associated with neurodegeneration and Prxs play an important role against this damage.
REFERENCES
- Alphey, M.S., C.S. Bond, E. Tetaud, A.H. Fairlamb and W.N. Hunter, 2000. The structure of reduced tryparedoxin peroxidase reveals a decamer and insight into reactivity of 2Cys-peroxiredoxins. J. Mol. Biol., 300: 903-916.
Direct Link - Beal, M.F., 1996. Mitochondria free radicals and neurodegeneration. Curr. Opin. Neurobiol., 6: 661-666.
CrossRef - Beal, M.F., 2002. Oxidatively modified proteins in aging and disease. Free Radic. Biol. Med., 32: 797-803.
CrossRefDirect Link - Bruchhaus, I., S. Richter and E. Tannich, 1997. Removal of hydrogen peroxide by the 29 kDa protein of Entamoeba histolytica. Biochem. J., 326: 785-789.
Direct Link - Bryk, R., P. Griffin and C. Nathan, 2000. Peroxynitrite reductase activity of bacterial peroxiredoxins. Nature, 407: 211-215.
CrossRefDirect Link - Butterfield, L.H., A. Menno, S.H. Golub and H. Shau, 1999. . From cytoprotection to tumor suppression the multifactorial role of peroxiredoxins. Antioxid. Redox Signal, 1: 385-402.
Direct Link - Chae, H.Z., I.H. Kim, K. Kim and S.G. Rhee, 1993. Cloning sequencing and mutation of thiol-specific antioxidant gene of Saccharomyces cerevisae. J. Biol. Chem., 268: 16815-16821.
Direct Link - Chae, H.Z. and S.G. Rhee, 1994. A thiol-specific antioxidant and sequence homology to various proteins of unknown function. Biofactors, 4: 177-180.
Direct Link - Chae, H.Z., K. Robison, L.B. Poole, G. Church, G. Storz and S.G. Rhee, 1994. Cloning and sequencing of thiol-specific antioxidant from mammalian brain: Alkyl hydroperoxide reductase and thiol-specific antioxidant define a large family of antioxidant enzymes. Proc. Natl. Acad. Sci. USA., 91: 7017-7021.
PubMedDirect Link - Chae, H.Z., S.J. Chung and S.G. Rhee, 1994. Thioredoxin-dependent peroxide reductase from yeast. J. Biol. Chem., 269: 27670-27678.
PubMedDirect Link - Chae, H.Z., H.J. Kim, S.W. Kang and S.G. Rhee, 1999. Characterization of three isoforms of mammalian peroxiredoxin that reduce peroxides in the presence of thioredoxin. Diabetes Res. Clin. Pract., 45: 101-112.
Direct Link - Chauhan, R. and S.C. Mande, 2001. Characterization of the Mycobacterium tuberculosis H37Rv alkyl hydroperoxidase AhpC points to the importance of ionic interactions in oligomerization and activity. Biochem. J., 354: 209-215.
PubMed - Chen, J.W., C. Dodia, S.I. Feinstein, M.K. Jain and A.B. Fisher, 2000. 1-Cys peroxiredoxins a bifunctional enzyme with glutathione peroxidase and phopholipase A2 activities. J. Biol. Chem., 275: 28421-28427.
Direct Link - Choi, H.J., S.W. Kang, C.H. Yang, S.G. Rhee and S.E. Ryu, 1998. Crystal structure of a novel human peroxidase enzyme at 20A resolution. Natl. Struct. Biol., 5: 400-406.
CrossRef - Chuchalin, A.G., V.I. Novoselov, O.N. Shifrina, S.K. Soodaeva and L.M. Barishnikova, 2003. PeroxiredoxinVI in human respirator system. Respiratory Medicine, 97: 47-151.
Direct Link - Claiborne, A., J.I. Yeh, T.C. Mallett, J.Luba and E.J. Crane, V. Charrier and D. Parsonage, 1999. Protein-sulfenic acids: Diverse roles for an unlikely player in enzyme catalysis and redox regulation. Biochemistry, 38: 15407-15416.
Direct Link - Dubuisson, M., D.V. Stricht, A. Clippe, F. Etienne and T. Nauser et al ., 2004. Human peroxiredoxin 5 is a peroxynitrite reductase. Febs Lett., 571: 161-165.
Direct Link - Fatma, N., D.P. Singh, T. Shinohara and J.LT. Chylack, 2001. Transcriptional regulation of the antioxidant protein 2 gene a thiol-specific antioxidant, by lens epithelium-derived growth factor to protect cells from oxidative stress. J. Biol. Chem., 276: 48899-48907.
Direct Link - Fisher, A.B., C. Dodia, Y. Manevich, J.W. Chen and S.I. Feinstein, 1999. Phospholipid hydroperoxides are substrates for non-selenium glutathione peroxidase. J. Biol. Chem., 274: 21326-21334.
Direct Link - Gommel, D.U., E. Nogoceke, M. Morr, M. Kiess, H.M. Kalisz and L. Flohe, 1997. Catalytic characteristics of tryparedoxin. Eur: Biochem. J., 248: 913-918.
Direct Link - Gulesserian, T., E. Engidawork, B.C. Yoo, N. Caims and G. Lubec, 2001. Alteration of caspases and their regulatory proteins in brains of down syndrome. J. Neural Trans., 61: 163-179.
Direct Link - Halliwell, B., 1992. Reactive oxygen species and the central nervous system. J. Neurochem., 59: 1609-1623.
CrossRefPubMedDirect Link - Jacobson, F.S., R.W. Morgan, M.F. Christman and B.N. Ames, 1989. An alkyl hydroperoxide reductase from Salmonella typhimurium involved in the defense of DNA against oxidative damage purification and properties. J. Biol. Chem., 264: 1488-1496.
Direct Link - Kim, J.A., S. Park, K. Kim, S.G. Rhee and S.W. Kang, 2005. Activity assay of mammalian 2-cys peroxiredoxins using yeast thioredoxin reductase system. Anal. Biochem., 338: 216-223.
Direct Link - Kim, K., I.H. Kim, K.Y. Lee, S.G. Rhee and E.R. Stadtman, 1988. The isolation and purification of a specific protector protein which inhibits enzyme inactivation by a thiol/Fe(III)/O2 mixed-function oxidation system. J. Biol. Chem., 263: 4704-4711.
Direct Link - Kim, S.H., R. Vikolinsky, N. Caims, M. Fountoulakis and G. Lubec, 2001. The reduction of NADH: Ubiquinone oxidoreductase 24- and 75-kDa subunits in brains of patients with down syndrome and Alzheimer's disease. Life Sci., 68: 2741-2750.
Direct Link - Koo, K.H., S. Lee, S.Y. Jeong, E.T. Kim and H.J. Kim et al., 2002. Regulation of thioredoxin peroxidase activity by C-terminal truncation. Arch. Biochem. Biophys., 397: 312-318.
CrossRef - Krapfenbauer, K., E. Engidawork, N. Cairns, M. Fountoulakis and G. Lubec, 2003. Aberrant expression of peroxiredoxin subtypes in neurodegenerative disorders. Brain Res., 967: 152-160.
Direct Link - Lee, S.P., Y.S. Hwang, Y.J. Kim, K.S. Kwon, H.J. Kim, K. Kim and H.Z. Chae, 2001. Cyclophilin A binds to peroxiredoxins and activates its peroxidase activity. J. Biol. Chem., 276: 29826-29832.
Direct Link - Lim, Y.S., M.K. Cha, H.K. Kim, T.B. Uhm, J.W. Park, K. Kim and I.H. Kim, 1993. Removals of hydrogen peroxide and hydroxyl radical by thiol-specific antioxidant protein as a possible role in vivo. Biochem. Biophys. Res. Commun., 192: 273-280.
Direct Link - Lim, Y.S., M.K. Cha, H.K. Kim, I.H. Kim, 1994. The thiol-specific antioxidant protein from human brain: gene cloning and analysis of conserved cysteine regions. Gene, 140: 279-284.
Direct Link - Lubec, G., 1996. The hydroxyl radical from chemistry to human disease. J. Invest. Med., 44: 324-346.
Direct Link - Manevich, Y., T. Sweitzer, J.H. Pak, S.I. Feinstein, V. Muzykantov and A.B. Fisher, 2002. 1-Cys peroxiredoxin overexpression protects cells against phospholipid peroxidation-mediated membrane damage. Proc. Natl. Acad. Sci. USA., 99: 11599-11604.
Direct Link - Manevich, Y. and A.B. Fisher, 2005. Peroxiredoxin 6, a 1-cys peroxiredoxin, functions in antioxidant defense and lung phopholipid metabolism. Free Rad. Biol. Med., 38: 1422-1432.
Direct Link - Matsumoto, A., A. Okado, T. Fujii, J. Fujii, M. Egashira, N. Niikawa and N. Taniguchi, 1999. Cloning of the peroxiredoxin gene family in rats and characterization of the fourth member. Febs Lett., 443: 246-250.
Direct Link - Oberley, T.D., E. Verwiebe, W. Zhong, S.W. Kang and S.G. Rhee, 2001. Localization of the thioredoxin system in normal rat kidney. Free Radic. Biol. Med., 30: 412-424.
CrossRefDirect Link - Phelan, S.A., 1991. AOP2 (antioxidant protein 2): Structure and function of a unique thiol-specific antioxidant. Antioxd. Redox. Signal, 1: 571-584.
PubMed - Seo, M.K., S.W. Kang, K. Kim, L.C. Baines, T.H. Lee and S.G. Rhee, 2000. Identification of a new type of mammalian peroxiredoxin that forms an intramolecular disulfide as a reaction intermediate. J. Biol. Chem., 275: 20346-20354.
Direct Link - Shau, H., R.K. Gupta and S.H. Golub, 1993. Identification of a natural killer enhancing factor (NKEF) from human erythroid cells. Cell Immunol., 147: 1-11.
Direct Link - Shau, H., L.H. Butterfield, R. Chiu and A. Kim, 1994. Cloning and sequence analysis of candidate human natural killer-enhancing factor genes. Immunogenetics, 40: 129-134.
CrossRef - Spyrou, G., E. Enmark, A. Miranda-Vizuete and J. Gustafsson, 1997. Cloning and expression of a novel mammalian thioredoxin. J. Biol. Chem., 272: 2936-2941.
Direct Link - Squire, T.C., 2001. Oxidative stress and protein aggregation. Exp. Gerontol., 26: 1539-1550.
CrossRef - Takeda, K., Y. Shimada, M. Amano, T. Sakal, T. Okada and I. Yoshiya, 1984. Plasma lipid peroxides and alpha-tocopherol in critically ill patients. J. Crit. Care. Med., 12: 957-959.
Direct Link - Uwayama, J., A. Hirayama, T. Yanagawa, E. Warabi and R. Sugimoto et al., 2006. Tissue Prx I in the protection against Fe-NTA and the reduction of nitroxyl radicals. Biochem. Biophy. Res. Commun., 339: 226-231.
Direct Link - Wang, V., Y. Manevich, S.I. Feinstein and A.B. Fisher, 2004. Adenovirus-mediated transfer of the 1-cys peroxiredoxin gene to mouse lung protects against hyperoxic injury. Am. J. Physiol. Lung Cell Mol. Physiol., 286: L1188-L1193.
CrossRef - Wong , C.M., Y. Zhou, R.W. Ng, H.F. Kung and D.Y. Jin, 2002. Cooperation of yeast peroxiredoxins Tsalp and Tsa2p in the cellular defense against oxidative and nitrosative stress. J. Biol. Chem., 277: 5385-5394.
Direct Link - Wood, Z.A., L.B. Poole, R.R. Hantgan and P.A. Karplus, 2002. Dimers to doughnuts redox-sensitive oligomerization of 2-cysteine peroxiredoxins. Biochem. J., 41: 5493-5504.
Direct Link - Wood, Z.A., E. Schroder, J.R. Harris and L.B. Poole, 2002. Structure mechanism and regulation of peroxiredoxins. Trends Biochem. Sci., 28: 32-40.
Direct Link - Yanagawa, T., K. Omura, H. Harada, T. Ishii, J. Uwayama, K. Nakaso and S.Y. Iwasa, 2005. Peroxiredoxin I expression in tongue squamous cell carcinomas as involved in tumor recurrence. Intl. J. Oral Maxillofac. Surg., 34: 915-920.
Direct Link - Sobh, A.M. and H.H. Mady, 2008. Peristaltic flow through a porous medium in a non-uniform channel. J. Applied Sci., 8: 1085-1090.
CrossRefDirect Link - Castagne, V., M. Gautschi, K. Lefevre, A. Posada and P.G.H. Clarke, 1999. Relationships between neuronal death and the cellular redox status focus on the developing nervous system. Prog. Neurobiol., 59: 397-423.
Direct Link - Cha, M.K., C.H. Yun and I.H. Kim, 2000. Interaction of human thiol-specific antioxidant protein 1 with erythrocyte plasma membrane. Biochem. J., 39: 6944-6950.
CrossRef