
Manee Archawaranon
Department of Biology, Faculty of Science, Ramkhamhaeng University, Bangkok 10240, Thailand
International Journal of Zoological Research
Year: 2005 | Volume: 1 | Issue: 1 | Page No.: 26-32







ABSTRACT
To study speech mimicry in hill mynahs Gracula religiosa is one of the channels to understand vocal learning in birds. Experiments were designed in order to study the critical period and social interaction affecting this learning behaviour. The variations of learning ability depended upon subspecies, sexes and sex hormone levels were also investigated. The studies were divided into five experiments. The results showed that the hand-reared nestlings in individual cages, after learning Thai words and phrases before six months of age for a year, were the most effective group to show vocal imitation (18 times/h). Meanwhile, wild birds older than six months, though reared and trained in individual cages, did not produce vocal imitation. The variations of vocal imitation between subspecies and sexes were not found. However, male birds with high vocal imitation efficiency (15 times/h) had higher testosterone levels than those with lower vocal imitation efficiency. Estradiol levels were high in female birds with high vocal imitation. The critical period of vocal imitation was the first six months of age and social contact with a trainer was required, without interacting with other hill mynahs. It seems likely that endocrine states, social interactions and vocal imitation output are correlated.
PDF Abstract XML References
How to cite this article
Manee Archawaranon, 2005. Vocal Imitation in Hill Mynahs Gracula religiosa: Factors Affecting Competency. International Journal of Zoological Research, 1: 26-32.
DOI: 10.3923/ijzr.2005.26.32
URL: https://scialert.net/abstract/?doi=ijzr.2005.26.32
DOI: 10.3923/ijzr.2005.26.32
URL: https://scialert.net/abstract/?doi=ijzr.2005.26.32
INTRODUCTION
Singing in passerine birds is a species specific communication which serves both territorial and sexual behaviour. Song learning is very crucial to how birds can survive in their lifetime fitness. They need to learn and develop species specific songs in which result their behaviours[1-4]. The evolutionary advantage of this behaviour is given. Territorial defence, mate attraction and individual recognition are suggested[5-8]. However, some birds mimic songs of other species in the wild[6,9]. Avian mimicry of the human voice is limited to a few species such as parrots and hill mynah[10]. Among these, a mynah produces vowel sounds most similar to those of humans[11].
Hill mynah Gracula religiosa is one of the most popular cage pets in Thailand and also in other countries. Their ability to imitate numerous diversity of sounds, particularly human speech in any languages is impressive. Most of the studies on hill mynah vocal imitation were done in captivity and involved conditioned learning together with vocal imitation responding[12-14], neuroanatomy as dendritic spine or neural coding of the speech control system in learned hill mynahs[15,16], neurophysiology[17,18]. None was reported on factors affecting vocal imitation ability of hill mynahs. Thus in this study, critical period of sound learning in hill mynahs was first dealt with, the age at which young were hand-reared and taught. Inter- and intra-specific social integration, the differences between subspecies, sexes and sex hormone levels were also examined.
MATERIALS AND METHODS
Two subspecies of hill mynahs found in Thailand, the northern race Gracula religiosa intermedia and the southern race G. r. religiosa[19] were studied of their vocal imitation ability, from 1996 to 2000. Birds of different ages and both sexes were studied, in specific experiments. All birds were donated by people who got them from poachers for cage pets. Age of birds was received from the owners and determined by characters of plumage, tarsus and wattle colour[20,21]. Adult birds (>one year), birds older than six months and nestlings less than four weeks old were used. Hand-rearing for nestlings was conducted until they were able to feed themselves, about forty-five days old. Therefore, except nestlings, birds had already been in flocks and interacted with the other members in the wild. The age of birds in flock was at least six months old because they were fledglings of that year and were caught in their first non-breeding season of the same year. Sexes were determined by sex chromosomes derived from the feather pulp[22].
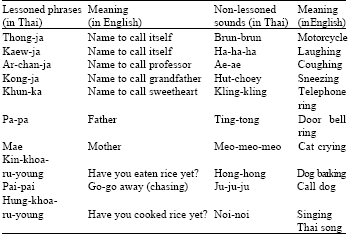
This study was divided into five experiments carried out at the Zoological Research Station, Ramkhamhaeng University, Bangna Campus and also at the Department of Biology, Ramkhamhaeng University, Main Campus. Birds were taught ten phrases in Thai language (lessoned phrases, Table 1) both by keepers and by tape cassettes twice a day in the morning at 0700-0800 h during period of cleaning, changing food and water and in the evening at 1500-1600 h. All of them had equal chances to hear any sound from outside the cages (non-lessoned sound). When the lesson was finished in each experiment, the number of times birds produced the lessoned phrases was recorded everyday during 0700-0800 h and 1500-1600 h for thirty days. One-way Analysis of Variance was used to test the differences of the times which learned birds produced the lessoned phrases. The non-lessoned sound was inevitable and not considered in this study.
Experiment I
Critical period: The experiment was designed to study the critical period at which hill mynahs learned and mimicked sound in captivity. Twenty-four northern birds, G. r. intermedia, were divided into seven groups (Table 2). Group 1 to Group 6 were composed of three to four nestlings in each group, Group 7 was four adult birds. They were kept in individual cages, 50x60x60 cm in size.
Birds of Groups 1 and 7 were taught the lesson immediately. Birds of Group 2 to 6 were first kept in the laboratory at the Department of Biology, Ramkhamhaeng University, Main Campus, without hearing the lesson until they were 3, 6, 7, 8, 9 months old respectively and then transferred to Bangna Campus. All seven groups were taught the lesson at the Zoological Research Station, Ramkhamhaeng University, Bangna Campus (Table 2). Vocal imitation ability was recorded at once after the lesson was finished for thirty days. The result of this experiment would show the ability to mimic sound at different ages.
Experiment II
Social interaction: This experiment was conducted in order to study the effect of inter- and intra-specific social interaction to vocal imitation. Twelve northern birds, G. r. intermedia, were equally divided into two groups. Both groups were nestlings, less than four weeks old. Group 1 were six birds kept in six individual cages, 50x60x60 cm while Group 2 were kept in two big aviaries size 4x8x3 m, three birds in each aviary with four adult birds. The lessoned phrases were taught immediately by keepers in three individual cages and one aviary and by tape cassettes in other three individual cages and another aviary for a year. The imitated phrases were recorded at once after the lesson was finished and the recording proceeded for thirty days. The results would indicate the difference of the ability of vocal imitation when birds were kept in different conditions and taught by different tools.
| Table 1: | Phrases in Thai language for teaching birds (lessoned phrases) and sounds from outside the cages (non-lessoned sounds) |
 | |
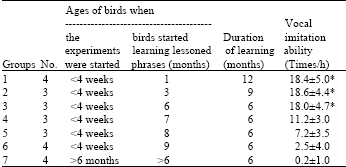
| Table 2: | Treatments of birds in experiment I and the ability to produce vocal imitation (M±SD) of birds which started to learn at different ages (* =p< 0.01) |
 | |
Experiment III
Subspecies difference: Northern birds, G. r. intermedia (n=6) and southern birds, G. r. religiosa (n=6) were taught the lessoned phrases for a year when they were less than four weeks old and kept in twelve individual cages. The vocal imitation was recorded in the same way as the previous experiments for thirty days. This would show the difference of ability between subspecies.
Experiment IV
Sex difference: The sexes of twelve northern birds, G. r. intermedia, were identified by inspecting sex chromosomes (ZW in female and ZZ in male)[22]. Male birds (n=6) and female birds (n=6) were taught the lessoned phrases for a year since they were one month old in twelve individual cages. The vocal imitation was recorded as in the previous experiments for thirty days. The results would indicate the difference of the ability of vocal imitation between sexes.
Experiment V
Sex hormone levels: Northern birds, G. r. intermedia, from Experiment I to IV were divided into four groups (Group 2-5). Group 1 (n=6) was the young at ages less than three months old and never produced any imitated sound. Group 2 (n=8) was adult birds and could produce only natural hill mynah sound. Group 3 (n=8) was birds with no imitation ability. Group 4 (n=8) was the ones which could make vocal imitation only twice in an hour. Group 5 (n=8) was the birds which could produce imitation 15 times/h. Blood was drawn twice, at three-month interval from all birds for measuring testosterone and estradiol level by using Enzyme-Linked Immunosorbent Assay (ELISA) [23]. The results would show the correlation between vocal imitation ability and sex hormone levels.
RESULTS
Experiment I
Critical period: It was shown that birds which were taken from the nest at less than four weeks old, kept individual cages and practiced the lessoned phrases before six months of age were the most effective groups to learn and imitate fluently. As in Table 2, birds which started lessoned learning when they were one, three and six months old could produce the vocal imitation averaging 18.3 times/h whereas birds which started learning at seven, eight and nine months of age could produce less imitation than the former three groups (p<0.01). The learning ability of the birds in Group 4, 5 and 6 decreased gradually when they got older as evidenced in Group 6 which started learning when they were nine months old and produced vocal imitation only 2.5 times/h. Wild birds in Group 7 which were over six months old when they were caught, produced no imitation and could only make natural hill mynah sound.
Experiment II
Social interaction: The results indicated that three birds in Group 1 which were in three individual cages and taught lesson by keepers produced most vocal imitation (18.2 times/h). While birds kept in individual cages and learned lesson from tape cassettes produced less imitation than the former but was not significantly different. Birds kept with adult birds in two aviaries could barely produce vocal imitation (Table 3).
Experiment III
Subspecies difference: No significant difference was found between two subspecies of hill mynahs in Thailand in producing vocal imitation (Table 4). Southern birds seemed to produce vocal imitation more frequently than the northern ones, though not statistically significant.
Experiment IV
Sex difference: There was no significant difference between sexes in the ability to learn and to produce vocal imitation (Table 5).
| Table 3: | The comparison of the ability to produce vocal imitation (M±SD)(n=3) between birds kept in individual cage and in aviary which were taught by keepers or by tape cassettes |
 | |
| Table 4: | The comparison of the ability to produce vocal imitation (M±SD) between northern and southern birds |
| Table 5: | The comparison of the ability to produce vocal imitation (M±SD) between males and females (n=6) |
Although females were likely to produce more vocal imitation (17 times/h) but were not significantly different from males (14.5 times/h).
Experiment V
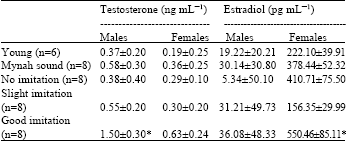
Sex hormone levels: From the comparison of testosterone levels among five groups (Table 6) of birds which had five levels of efficiency in vocal imitation, it was shown that males of Group 5 which produced the most vocal imitation (15 times/h) had the highest testosterone level (1.5 ng mL-1) and was significantly different from males of Group 4 (0.55 ng mL-1) (p<0.01) which produced vocal imitation 2 times/h. Apart from this, the difference of the testosterone levels among Group 1-4 were not significant.
Estradiol levels in females revealed that the differences of vocal imitation ability were related to the differences of hormone levels. Female birds of Group 5 had significantly higher estradiol levels (550 pg mL-1) than the other groups (p<0.01).
DISCUSSION
In avian species, the most common communication signal is auditory. Birds use sound as special designed signal, so called songs, to activate or modify behaviour of the receivers. Species specific patterns of songs or repertoires are important for various functions. Most males sing songs in breeding season for territorial defense and mate attraction. Using sound also plays a role in individual and species recognition.
Vocal learning in birds occur in the critical period which depends upon each species but they would sing when the suitable time comes later in their life times. In the temperate zone, Chaffinches Fringilla coelebs which hatch in spring from May to July learn songs during June to August and sing in the next spring. Learning in the critical period is very influential for living the rest of their lives.
| Table 6: | The comparison of testosterone and estradiol levels (M±SD) among five groups of birds which produced different vocal imitation abilities in experiment V (* = p < 0.01) |
 | |
Parents and young birds form adhesive social bond in order to send species specific songs to the next generation correctly[8]. There were experiments revealed that when young Chaffinches which were less than one year old were kept separately from adult birds and never heard species songs of the latter, their songs were as simple as those of young birds kept together in flock[24,25]. Birds which sang their own species songs accurately were the ones that heard adult songs either from living birds or tape cassettes.
The critical period to learn the speech or sounds of hill mynahs was evidently within the first six months of age. The birds had been kept individually since they were less than four weeks old and listened to lessoned phrases solely. Therefore, during their critical period, they learned only what we forced them to learn. Without any contact with the other birds, they formed social contact with the keepers. As for birds from the wild, when they were older than six months old, they have already learned sounds from the other birds in the flock and since they already passed the critical period, they produced no vocal imitation. Although most of birds learn only species songs, there is 15-20% of passerine birds which imitate other species[5,6,8,26,27]. However, trained birds are more likely to learn from living specimen than from tape cassettes (African grey parrots Psittacus erithacus[27-29], starlings Sturnus vulgaris[30], zebra finches Poephila guttata[31], cowbird Molothrus ater[32]). This also occurred in hill mynahs, they liked to learn from the keepers. Social contact is a prominent way from which birds choose to learn during their critical periods.
The difference of subspecies did not make any difference in vocal imitation ability of hill mynahs. The factor of geographic variation normally results in conspecific songs as dialects. Song learning of different subspecies of white-crowned Sparrows Zonotrichia leucophrys oriantha, migratory population and Z.l. nuttalli, sedentary birds depended on reproductive behaviour[33]. However, the vocal imitation in hill mynahs was not a song and both subspecies listened to the same lessoned phrases during their critical period, they did not learn sounds from the nature which may represent dialects. It is interesting to conduct further study on vocalization of these birds in the forest of Thailand. In the study on the geographic variation of northern birds G. r. intermedia the differences of chip-calls between birds from different regions were greater than the differences between the individuals in each region[34]. This study showed the results concerning birds of the same subspecies but captured from different regions. It seems likely that birds in the nature either in the same or different subspecies inhabiting in different regions generally have dialects.
The ability to learn songs may not be much different, the sets of chromosomes of both races of hill mynahs are not different[35] which consequently reflect no genetic difference in vocal learning. However, the variability of mtDNA was not related to song dialects of the rufous-collared sparrow Zonotrichia capensis, studied in northwestern Argentina[36]. This is in contrast with white- throated sparrows Z. albicollis which have two colour morphs. There were relationships between the volume of nuclei in the avian song system and song production. White-striped birds which sang more frequently had larger song nuclei than those of the tan- striped ones[37]. In hill mynahs, there has been no study on neurophysiology of vocal imitation comparisons between subspecies. Therefore, it is quite difficult to tell the correlation between subspecies and imitative vocalization by means of neuroanatomy.
Although it has been accepted for years that most of male rather than female birds sang, in the Experiment IV, female hill mynahs had talent in vocal imitation at the same levels as their male counterparts. Hormones have effects on vocal learning and imitation in this bird. Because the results in Experiment V showed that both males and females which were able to imitate the lessoned phrases quite well (15 times/h) had higher levels of testosterone and estradiol than the other groups which had lower frequency of vocal imitation. However, the question of how and when the effects of hormones initiate remains to be answered. In sparrows, the critical period is about first three months old and they produce song when they are seven to eight months old[38]. The experiment which stimulated them to sing earlier than expected became successful by administering three months old birds with testosterone which made them sing simple songs. Normally, after birds learn songs during their critical periods and pass the memorization phases they start to sing whenever testosterone circulate in their bodies. Normally female passerine birds rarely sing. But that hormone administration in female birds stimulates them to sing is well evidenced (white-crowned sparrow[26], European starlings[39], budgerigars Melopsittacus undulatus[40] ). Although white-throated sparrows males hardly sang in winter, testosterone implanted birds sang during non-breeding season[41]. It is possible that testosterone has an effect on increasing the amount and volume of high vocal centers which are song nuclei in the pathway of vocal learning in brain (canaries[42], European starlings[43], song sparrows Melospiza melodia[44] ). Female wrens which did not sing as much as males had song nuclei less sensitive to steroids than males[45]. But testosterone administration in female canaries increased high vocal center and enhanced females to sing more[46]. Exogenous administration of sex steroids may increase vocal imitation ability in hill mynahs as well.
There is no exact explanation why some birds imitate other species’ songs in the wild. There are a lot of hypotheses and experiments concerning this inquiry. Vocal imitation in birds occurred when two different species lived in the same environment and had the same predators[5]. Other hypotheses included no specific patterns of songs to learn, to enlarge the repertoire size of their own songs to make it more complicated, the purpose of individual recognition, mate recognition, threatening and most likely to deceive predators or parasite young birds luring parent host birds to feed them. Hill mynah was said not to sing in the wild[47] and it was scarcely territorial[34]. Their utterances were elaborate contact notes for keeping mates or flocks together.
There were findings that hill mynah could imitate other species’ sound in the nature[48] such as alarm call of Kloss’ s Gibbon and spacing call of male Pig-tailed Langurs[49]. In contrast, there were many reports revealed that hill mynah could not do so especially in the wild but they could do well in captivity[34,47,50]. In the wild they may not have other species’ song to imitate[34,51]. Male hill mynahs imitated neighboring males and that females imitated neighboring females[34]. If birds were allowed to learn solely from their own species’ sound and interact within the flock during their critical period, they would not attend to learn anything else. This occurred in Experiment I, where birds of group 7 were captured from the wild when they were over six months old. In captivity, when birds were kept individually, the deprivation of normal social opportunities made them attend to and accept a wide ranges of sounds[50,52].
It seems likely that birds are more concerned with immediate responses from companions than with their utterances. The reinforcement for the vocal imitation comes mainly from their interest in companions or keepers and form social contact[53] as called imprinting[34].
Conditioned learning was also studied in hill mynahs. The study on stimulus control over speech in hill mynahs showed that associate learning on vocalization occurred in this bird[13]. There are very few studies on neurophysiology of vocal imitation in hill mynah. An example is the neural coding of speech sound in the telencephalic auditory area of hill mynah[16]. Furthermore, it was found that speech-trained hill mynah had a reduction of dendritic spine density and an enlargement of the remaining spine of neurons in the telencephalic nucleus hyperstriatum ventrale pars caudale. This finding was interpreted that hill mynah’s potential to imitate new sounds shrank with age. Results of Experiment I indicated that not only they passed their critical periods when they were older than six months old, the ability of vocal learning decreased with age.
The experiments in this study emphasized factors affecting competency and were not concerned with neurophysiology of vocal imitation. It is quite interesting to investigate the differences of the anatomy and physiology of neurons in the brain which control vocal learning between intact and manipulated mimic birds. However, although the study of neurophysiology on vocal imitation will disclose the knowledge that why some birds such as hill mynah or parrots can do imitation while the others can not, the sacrifice of well-trained mimic birds in order to study their neurons in this aspect is involved with ethic.
ACKNOWLEDGMENTS
I am grateful to R. Haven Wiley and H. Mueller, Department of Biology, The University of North Carolina at Chapel Hill, North Carolina, USA. for inspiring me in working on Ornithology. I thank Pornchai Wongwasana for his patient in collecting data of these long-term experiments. This study was supported by The Thailand Research Fund.
REFERENCES
- Archawaranon, M., 2002. Zoogeography of various hill mynah phenotypes in Thailand. J. Biol. Sci., 2: 645-647.
CrossRefDirect Link - Archawaranon, M., 2003. The impact of human interference on Hill Mynahs Gracula religiosa breeding in Thailand. Bird Conserv. Int., 13: 139-149.
CrossRefDirect Link - Archawaranon, M., 2005. Captive Hill Mynah Gracula religiosa breeding success: Potential for bird conservation in Thailand? Bird Conserv. Int., 4: 679-682.
Direct Link - Archawaranon, M., 2004. Rapid sexing hill Mynah Gracula religiosa by sex chromosomes. Biotechnology, 3: 160-164.
CrossRefDirect Link - Archawaranon, M. and U. Mevatee, 2002. Karyotypes among various morphological populations of Hill Mynah, Gracula religiosa, from different areas in Thailand. J. Biol. Sci., 2: 386-389.
CrossRefDirect Link