
A.H. Khatab Eman
Department of the Virus and Phytoplasma, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
N. Zein Salwa
Department of the Virus and Phytoplasma, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
A. Ahmed Amal
Department of the Virus and Phytoplasma, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 2 | Page No.: 224-233







ABSTRACT
Broad bean true mosaic virus was purified from infected faba bean plants. The UV absorption spectrum had a maximum at 260 nm and a minimum at 240 nm. The ratios of Amax /A min and A 260/A 280 were 1.48 and 1.71, respectively. The yield of purified BBTMV preparation was about 1.57 mg kg-1 infected leaves. Production of specific antiserum against BBTMV was obtained by rabbit immunization using three different methods of injections. The titer of the prepared antiserum as determined by indirect ELISA with dilution of tissues 1/5 was 1/1024. Purification of the immunogamaglobulin G and conjugate of IgG with alkaline phosphatase were carried out to be used for ELISA detection. The concentrations of IgG and IgG conjugated with alkaline phosphatase were 1.0 mg mL-1 and 1:1000, respectively. Antiserum produced specific to BBTMV was used for virus detection by using different serological diagnostic methods. The percentages of virus detection in leaves, stems were higher than that in roots; on the other hand the percentages of virus detection in anthers and pistils were higher than that in sepals.
PDF Abstract XML References Citation
Received: September 20, 2011;
Accepted: October 24, 2011;
Published: December 02, 2011
How to cite this article
A.H. Khatab Eman, N. Zein Salwa and A. Ahmed Amal, 2012. Purification, Serology and Prevalence of Broad Bean True Mosaic Comovirus (BBTMV). International Journal of Virology, 8: 224-233.
DOI: 10.3923/ijv.2012.224.233
URL: https://scialert.net/abstract/?doi=ijv.2012.224.233
DOI: 10.3923/ijv.2012.224.233
URL: https://scialert.net/abstract/?doi=ijv.2012.224.233
INTRODUCTION
Faba bean is widely cultivated in Egypt for its value as food, forage and green manure. Broad bean true mosaic virus (BBTMV) is one of a seed borne viruses, which infect temperate pulse crops, It was first describe in Vicia faba crops in Germany and was then found in England in 1959 and 1960 (Quantz, 1953).
BBTMV is transmitted by mechanical inoculation and by seeds. Its host range is very narrow and limited to the Fabaceae and it is only seed borne in Vicia species. It appears most likely to survive between cropping seasons in infected Vicia seed (Cockbain et al., 1976). Vorra-Urai and Cockbain (1977) reported experimental seed transmission rates of up to 5.3 % in broad bean (Vicia faba var. major) when 4 week old seedlings were inoculated in the glasshouse. Mali et al. (2003) found that BBTMV was seed transmitted in a range of faba bean and Vetch cultivars, at rates of up to 28% of seed. It occurs mainly in Europe and has been reported as the cause of significant virus disease problem in faba bean production in Great Britain and Western and Eastern European countries. It has also been reported in virus surveys in North Africa.
Thus, this investigation aimed to prepare of purified BBTMV for production of specific antiserum and purification of IgG which conjugated with alkaline phosphatase for application of serological diagnostic methods such as direct and indirect-ELISA, TBIA and DBIA.
MATERIALS AND METHODS
Source of virus isolate: Faba bean seeds supplied from Field Crops Research Institute were tested for virus detection. Samples of seedlings emerged from collected seeds which gave positive reaction with antiserum were used as virus source in this study.
Purification of the virus isolate: BBTMV was purified according to the method described by Jefferson and Makkouk (1986) with some modifications. Two hundred grams of faba bean cv. Giza 3 leaves showing distinct symptoms, 21 days after virus inoculation were collected and pulverized in liquid nitrogen.
The pulverized tissues were homogenized with a blender in 400 mL of 0.5 M potassium phosphate buffer, containing 0.05 M ethylenediamine tetra acetic acid (EDTA), 0.5 % 0.5 % sodium sulfite (Na2CO3) and 0.01 M diethyldithiocarbamate (DIECA), pH 8. The homogenate was filtered through two layers of cheesecloth and subjected to LSC at 8000 rpm for 10 min. The supernatant obtained was stirred with a mixture of 25% chloroform and 25% carbon tetrachloride for 30 min and the aqueous phase was separated by LSC at 8000 rpm for 10 min at 4°C in a Beckman J-21 C centrifuge using JA-20 rotor. The virus was precipitated from the supernatant with 6% polyethylene glycol (PEG 6000) and 0.3 M sodium chloride (NaCl) stirring overnight at 4°C. The pellets were collected by 20 min centrifugation at 10000 rpm at 4°C and suspended in 100 mL of 0.01 M borate buffer, pH 7.5, containing 0.1% 2- mercabtoethanol (2-ME) and left overnight at 4°C with slowly stirring then centrifuged for 10 min at 8000 rpm to eliminate any- soluble materials followed by centrifugation for 90 min at 35.000 centrifuged for 2 h at 28,000 rpm in a Beckman SW 60 rotor at 4°C. Gradient columns were stored overnight at 8°C prior to use. The virus zones were collected with a bent tip hypodermal needle and syringe, diluted 1:1 with 0.001 M borate buffer, pH 7.5, without additives, and then concentrated by centrifugation for 90 min at 36.000 rpm. Final pellet were suspended in 1 mL of 0.001 M borate buffer and clarified using LSC at 6000 rpm for 10 min. Infectivity was tested on leaves of broad bean seedlings.
Absorption of the purified virus and determination of virus concentration: The UV-absorption spectra of BBTMV purified preparation was determined with spectronic 2000 spectrophotometer. The concentration was calculated using the absorbance value at 260 nm and extension coefficient of 7.7 (Gibbs et al., 1968). The values of a minimum and a maximum for the purified virus as well as the ratios rpm at 4°C in a Beckman L8-80 M of A260/A280 and Amax/min were ultracentrifuge using rotor 80 Ti. The pellets were resuspended in 0.0.01 M borate buffer, pH 7.5, and then layered on top of 10-40% sucrose gradients prepared in 0.01 M borate buffer, pH 7.8 and calculated.
Antiserum production of BBTMV: Rabbit immunization: Two NewZealand white rabbits, each one weighted 3 kg were used for antiserum raised against BBTMV. A total of 8 mg purified BBTMV was used for injection as described in Table 1. One mL (1 mg mL-1) of purified virus was emulsified with an equal volume of freund's complete adjuvant for subcutaneous or intramuscular injections at weekly intervals. Intravenous injection without adjuvant was given.
Intramuscular injection was performed in the right and the left band thighs, respectively using a 5 mL disposable syringe. Virus was diluted with phosphate buffer, pH 7.2, (V/V). Intravenous injection was made in the left ear at the marginal vein using 1 mL insulin disposable syringes.
| Table 1: | The rabbit immunization scheme used for antiserum production against BBTMV |
 | |
Rabbit bleeding: Rabbits were bleeding 10 days after the last injection from the right ear. The blood was collected, left to clot at 37°C in an incubator for 1-2 h and then kept at 4°C overnight.
Blood collection and separation of serum: Antiserum was separated through centrifugation at 4000 rpm for 20 min. The antiserum was collected and stored at 4°C until used for titer determination and other serological tests (Noordam, 1973).
Determination of antiserum titer: Antiserum titer was measured with indirect ELISA test. Clarified sap of virus infected and control broad bean leaves were diluted at 1/5 using phosphate buffer, pH 7.2, containing 0.85% NaCl. BBTMV antiserum preparation was diluted with the serum buffer, 1/1, 1/2, 1/4, 1/8, 1/16, 1/1/32, 1/64, 1/128, 1/256, 1/512, 1/1024 and 1/2048, respectively. The reaction was done between infected clarified extract and its induced antiserum by indirect ELISA test according to methods described by Lommel et al. (1982).
Purification of immunoglobulin (IgG): Immunoglobulins were purified from the antiserum using the method recorded by Steinbuch and Audran (1969). One mL of virus antiserum was added to 2 mL of 0.06 M Sodium acetate buffer, pH 4.8 (1:2), then dialyzed against this buffer (0.06 M Sodium acetate) for about 24 h (three times). While stirring vigorously add drop wise 0.082 mL actonic acid (caprylic acid) continuous stirring for 30 min at room temperature, then centrifuged at 8000 for 10 min. The supernatant was collected and dialyzed twice against 0.05 M phosphate buffer, pH 7.2, for 4 h each.
The resulting IgG was diluted with distilled water to make 4 mL and equal volume of saturated ammonium sulphate solution were added at room temperature while stirring and left at continuous stirring for 30 min. After centrifugation at 8000 rpm for 10 min, the pellet was collected and suspended in 1 mL distilled water followed by dialysis three times against 0.05 M phosphate buffer, pH 7.2 for 24-48 h (three times). If necessary the IgG was centrifuged clear 10 min at 8000 rpm. The IgG concentration was then adjusted to 1 mg mL-1 (A280 nm = 1.4) and stored at -20 until use.
Conjugation of alkaline phosphatase with IgG: IgG was conjugated with alkaline phosphatase according to protocol given by Clark and Adams (1977). Two mg alkaline phosphatase was dissolved in 1 mL of the above IgG preparation and dialyzed extensively against PBS (0.02 M phosphate buffer saline, pH 7.4). Glutaraldehyde was added to 0.05% final concentration and the mixture was incubated at 22°C for 4 h. Glutaraldehyde was then removed by dialysis five times against PBS buffer. Sodium azied 0.02% was added as preservative. The volume obtained was measured and bovine albumin (Sigma A-4503) at the rate of 5 mg bovine albumin to 1 mL solution was added and stored at 4°C. The concentration of IgG and IgG conjugate was determined using direct ELISA. IgG conjugate was diluted to 1/250, 1/500, 1/1000 and 1/2000 with the conjugate buffer, while the IgG was diluted with the coated buffer to concentrations of 0.5, 1.0,2 and 4 μg mL-1, respectively. Controls of healthy and infected broad bean sap were used. The reaction was done between IgG and IgG conjugate, by DAS = ELISA as follows. One hundred μL of purified gamaglobulin diluted in coating buffer (0.5, 1.0, 2.0 and 4.0 μg mL-1) were added to each tested well of micro titer plate. The plate was incubated at 37°C for 4 h or overnight at 4°C, then the plate was washed as above. Two hundred μL IgG conjugate diluted (1/250, 1/500, 1/1000 and 1/2000) with conjugate buffer were added to each well. The plate was incubated at 37°C for 4 h or overnight at 4°C and washed 4 times. Two hundred μL aliquots of freshly prepared substrate (5.0 mg nitrophenyl phosphate (Sigma 104-105) dissolved in 10 mL substrate buffer) were added to each well and then incubated at room temperature for 30-60 min. The reaction was stopped by adding 50 μL 3.0 M NaOH to each well. The absorbance was measured at 405 nm by ELISA reader. Reading twice those of healthy plant control average was considered positive.
Immunological detection of BBTMV by tissue blot immunoassay (TBIA) and dot blot immunoassay (DBIA) on nitrocellulose membranes: The technique of TBIA described by Lin et al. (1990) with some modification on nitrocellulose membranes were used for BBTMV detection. Nitrocelluose membrane, 0.45 mM pore size were marked with a lead pencil into squares of 1x1 cm. Rolled up fresh leaf tissues and/or stems were cut with razor blade for each sample. Exposed cut edges of tested tissues were pressed steadily, but not firmly into MCM. The membranes were washed three times with PBST at 5 min interval, incubated in 1% bovine serum albumin+2% nonfat dried milk in PBST for one hour at room temperature and washed after blocking with PBS-BSA (bovine serum albumin). The MCM were then incubated with virus specific antibodies diluted in PBS 1:2000 for two hours at room temperature. Following three successive washing in PBST, 5 min in interval the MCM were incubated with alkaline phosphatase labeled with goat antirabbit immunoglobulins (dilution 1:1000) and incubated for one hour at room temperature followed by three times washing in PBST. The membrane was incubated in substrate solution, NitroBlue Tetrazolium (NPT) and 5- bromo 4 chloro-3- indolylphosphate (BCIP) for 5 min. After color has appeared membrane was rinsed, membranes were rinsed quickly with H2O and then air-dried.
Dot blot immunoassay (DBIA): The technique of DBIA described by Lin et al. (1990) with some modifications on nitrocellulose membranes was used for BBTMV detection. Nitocellulose membrane, 0.45 mM pore size were marked with a lead pencil into squares of 1x1 cm. Healthy and infected faba bean plant samples were ground in phosphate buffer pH 8.5 (1:6 w/v). Five microliters from each healthy and infected samples were placed on the membrane. The membranes were washed three times with PBST at 5 min interval, placed in the PBS blocking solution for one hour at room temperature, washed tree times with PBST at 5 min interval and incubated with specific BBTMV antibodies separately in dilutions of 1:1000 in) BS for one hour at room temperature with gently shaking. The membranes were then washed three times in PBST and incubated with antirabbit conjugate (dilution 1:1000 in conjugate buffer) for one hour at room temperature. The membranes were washed three times in PBST and incubated 5 min in a substrate solution. After color has appeared, membranes were rinsed quickly with H2O and then air-dried.
Prevalence of the virus in different plant organs: Leaves, flower, stem and roots from faba bean (cultivars Giza3) infected with BBTMV were aseptically separated to their morphological organs, pistils, sepals, petals and anther were collected; each organ was homogenized in phosphate buffer saline in polyethylene small bags. Each extract was used as antigen using direct ELISA technique.
Prevalence of the virus in seeds and seed parts for detection of seed-borne virus: Upper, medium and flower pods were taken from BBTMV infected faba bean plants. Seeds from pods of each stage were taken at random selection. The seeds aseptically dissected into three parts: seed coats, cotyledon and embryos.
The different parts of seeds were washed several times by rinsing in distilled water for 10 min. The dissected seed portions homogenized separately in Phosphate Buffer Saline (PBS) by pressure- crushing in polyethylene bags. The bags were then shaken within one hour at room temperature. Each extract was used as antigen using direct ELISA technique.
RESULTS
Purification of BBTMV: BBTMV was purified with a high degree of purity. Three zone were observed at 2.5-3 cm. and 3.5 below the meniscus of sucrose density-gradient columns. These zones when collected together were found infectious (when tested on the indicator host plants). The absorption spectrum of the purified virus was typical for nucleoprotein as determined with a Spectrometer 2000 Spectrophotometer with a maximum at 260 nm. and a minimum at 240 nm (Fig. 1). The UV-absorbance ratios of a max/min and A260/280 were 1.48 and 1.71, respectively (for the two lower zones, collected together). The yield of purified BBTMV preparation was 1.57 mg kg-1 infected faba bean leaves.
Production of antiserum specific to BBTMV: Purified BBTMV preparation was used for rabbit immunization. Determination of antiserum titer: The polyclonal antibodies were obtained from rabbit bleedings taken 10 days after the last injection. Data presented in Table 2 show that the titer of antiserum against BBTMV was 1/1024 as determined by indirect ELISA technique, when infected sap diluted to 1/5 was used in ELISA system. Antiserum produced was used in the subsequent experiments. ELISA reaction was considered positive when the A405 values were greater than twice of healthy controls.
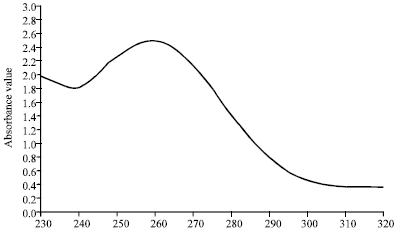 | |
| Fig. 1: | Ultraviolet absorption spectrum of purified BBTMV preparation |
| Table 2: | Determination of antiserum titer against BBTMV using indirect ELISA |
 | |
| * Reading after 30 min incubation with substrate | |
| Table 3: | Schematic diagram of checker broad arrangement for determination of approximate working dilution of IgG and IgG conjugate to BBTMV for ELISA test |
 | |
| Inf : Infected plant, H: Healthy plant | |
Purification of immunoglobulins (IgG): Purification of IgG was done using caprylic acid method as described before. Its concentration was adjusted to 1 mg mL-1 (A 280 nm = 1.4).
Conjugation of IgG with alkaline phosphatase: Positive reactions obtained when purified IgG and IgG conjugate of the virus were tested against BBTMV-infected plants. The concentration of IgG and IgG conjugated with alkaline phosphatase for BBTMV were 1.0 mg mL-1 and 1:1000, respectively (Table 3).
Immunological detection of BBTMV by tissue blot immunoassay (TBIA) and dot blot immunoassay (DBIA) on nitrocellulose membranes using antiserum production: Tissue blot immunoassay and Dot blot immunoassay were found to be sensitive to detect BBTMV in infected faba bean plants (Fig. 2).
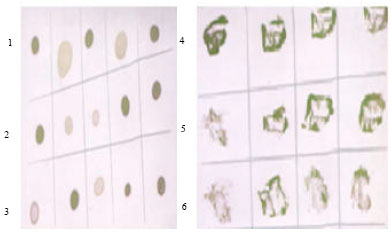 | |
| Fig. 2: | BBTMV was identified by DBIA technique (a) and TBIA (b) using polyclonal antibodies. Healthy and infected faba bean cv. Giza 3 plant samples [lane 1 (five leaves samples) and lane 4 (four stems samples)] from Agricultural Research Center Experimental Station, Giza [lane 2 (five leaves samples) and lane 5 (four stems samples)] Qalyubia [lane 3 (five leaves samples) and lane 6 (four stems samples)] Sharqiya Governorate. Purplish blue color indicates positive reaction |
| Table 4: | Prevalence of BBTMV in different oranges of faba bean plants using DAS- ELISA |
 | |
A purplish blue color was obtained from tissues in positive reactions, whereas extracts from healthy plants remain green in negative reactions.
Prevalence of the virus in different plant organs: Leaves, stems, flowers (sepals, petals and sexual organs) and roots infected with BBTMV were tested using DAS-ELISA. Results in Table 4 indicate that BBTMV was detected in leaves, stems, roots and all flower parts (sepals, petals, pistils and anthers). The percentages of virus detection in leaves and stem were higher than that in roots, whereas the percentages of anthers and pistils were higher than in sepals.
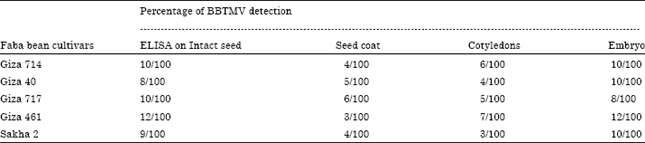
Prevalence of the virus in seeds and seed parts using DAS- ELISA: Data in Table 5 indicate that BBTMV was reliably detected in intact seeds and all seed parts (seed coats, cotyledon and embryos) of immature seeds in faba bean cvs. Giza 714, Giza 717, Giza 416 and Sakha 2.
| Table 5: | Percentage of BBTMV detection in different parts of seeds in five faba bean cultivars using DAS-ELISA |
 | |
DISCUSSION
When plant pathologists, become involved in immunology, the goal, generally is to generate an antibody probe which will significantly identify a target antigen in the assay. One of the most important objectives of this investigation is to produce immunogamaglobulins (IgG) and IgG conjugate specific to BBTMV. In this investigation using the modified method of purification, the present isolate of BBTMV was purified with a relatively high degree of purity and sufficient quantities for antiserum production. After sucrose density-gradient centrifugation of purified virus three zones were observed one faint upper and two distinct lower zones were collected from the sucrose density-gradient columns. The two lower zones were found infective when collected together because the separation of each zone alone was very difficult. The two lower zones were collected, pelleted, resuspended and tested for purity by UV-absorption spectra. The three boundaries one faint and two distinct lower zones resembling the three components in preparations of bean pod mottle virus (Semancik and Bancroft, 1964). Gibbs et al. (1968) found that, the purified preparations of BBSV produce three boundaries with sedimentation coefficients of 127, 100 and 60 s. the slowest sedimenting particles (top component) consist of protein only. The fastest sedimenting ones (bottom) contain more nucleic acid (36%) than those sedimenting at intermediate (25%) speed. The maximum and minimum absorbances were 260 nm and 240 nm, respectively. The UV-absorbance ratios of max/min and A260/280 were 1.48 and 1.71, respectively. The yield of the purified BBTMV preparation was 1.57 mg kg-1 of faba bean leaves. The predominant particle in samples from the virus zone was isometric and about 25 nm in diameter. Similar results were obtained by (Matthews, 1991).
Purified virus preparation used for production of polyclonal antiserum should be free of contaminating plant antigens. Even when a preparation appears to be pure by biochemical criteria, it frequently happens that a lengthy immunization schedule leads to the appearance of antibodies to contaminating substances that were present in trace amounts, these preparation that may contain plant material when injected into rabbits causes the production of antibodies, which react to this material in the serological test (Van-Regenmortel, 1982). It if for this reason that the amount of antigenic material administered to animals should be kept to a minimum (Matthews, 1991). In the present study, polyclonal antibody raised against BBTMV was prepared. The antiserum produced against BBTMV had titers of 1024 as determined by indirect ELISA technique. However, since the caprylic acid method is simpler and less time consuming (Makkouk et al., 1987) it was adopted for the isolation for BBTMV immunoglobulin. The concentrations of IgG and IgG conjugated with alkaline phosphatase were 1.0 mg mL-1 and 1:1000 for BBTMV using direct ELISA test clearly showed that, the IgG and IgG conjugate can be readily applied for virus detection in infected faba bean extracts. Since the production of polyclonal antibodies specific to BBTMV one of the main objectives of this work to cover the continues increasing needs for accurate and fast virus detection and for production of virus-free materials (Khattab, 2002).
Present results showed that, the detection of BBTMV in infected faba bean could be readily applied by tissue blot immunoassay (TBIA) and dot-blot immunoassay (DBIA) on nitrocellulose membranes (Makkouk et al., 1987; Makkouk and Kumari, 1996). In the present study, BBTMV was detected in leaves, stems, roots and different flower parts (sepals, petals, pistils and anthers) of faba bean cultivars Giza 461. These results are in harmony with those obtained by Sidaros et al. (2006).
Present results indicated that, BBTMV was detected in intact seeds and all seed parts (seed coats, cotyledons and embryos) of immature seeds of all tested faba bean cvs. These results are in agreement with those reported by Mali et al. (2003).
REFERENCES
- Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Cockbain, A.J., R. Bowen and S. Vorra-Urai, 1976. Seed transmission of broad bean stain virus and echtes ackerbohnenmosaik-virus in field beans (Viciafaba). Ann. Applied Biol., 84: 321-332.
CrossRef - Gibbs, A.J., G. Giussani-Belli and H.G. Smith, 1968. Broad-bean stain and true broad-bean mosaic viruses. Ann. Applied Biol., 61: 99-107.
CrossRef - Lommel, S.A., A.H. McCain and T.J. Morris, 1981. Evaluation of indirect enzyme-linked immunosorbent assay for the detection of plant viruses. Phytopathology, 72: 1018-1022.
CrossRefDirect Link - Lin, N.S., Y.H. Hsu and H.T. Hsu, 1990. Immunological detection of plant viruses and a mycoplasmalike organism by direct tissue blotting on nitrocellulose membranes. Phytopathology, 80: 824-828.
CrossRefDirect Link - Semancik, J.S. and J.B. Bancroft, 1964. Further characterization of the nucleoprotein components of bean pod mottle virus. Virology, 22: 33-39.
CrossRef - Steinbuch, M. and R. Audran, 1969. The isolation of IgG from mammalian sera with the aid of caprylic acid. Arch. Biochem. Biophys., 134: 279-284.
CrossRef