
H.S. El-Helaly
Plant Pathology Section, Faculty of Agriculture, Minufiya University Shebin El-Kom, Egypt
A.A. Ahmed
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
M.A. Awad
Plant Pathology Section, Faculty of Agriculture, Minufiya University Shebin El-Kom, Egypt
A.M. Soliman
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 106-113







ABSTRACT
Alfalfa Mosaic Virus (AMV) was isolated from naturally infected potato (Solanum tuberosum, L. cv. Spunta) plants, showing bright yellow blotching or mottling Calico and aucuba symptoms suspected to be due to virus infection. Leaf samples were collected from different localities in Minufyia Governorate, Egypt. AMV was readily mechanically inoculated by sap extracted from infected potato leaves to host plants (bioassay). The identity of AMV isolate was confirmed by sequence analysis of their Coat Protein (CP) gene. The sequence of the Coat Protein gene (CP) of AMV was determined from cDNA clones. The CP gene was cloned into pGE®-T Easy vector and transformed into Escherichia coli strain DH5α. The recombinant plasmids were obtained and sequenced. The nucleotide sequences were compared with corresponding viral nucleotide sequences reported in GenBank. The analysis showed that nucleotide sequence of the Egyptian isolate of AMV (GenBank Accession No. HQ288892) had percentage of similarity ranged from 97-98% with eight reported AMV isolates. Thus, a method of identification and detection by RT-PCR of AMV was established.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: July 23, 2011;
Published: October 25, 2011
How to cite this article
H.S. El-Helaly, A.A. Ahmed, M.A. Awad and A.M. Soliman, 2012. Biological and Molecular Characterization of Potato Infecting Alfalfa Mosaic Virus in Egypt. International Journal of Virology, 8: 106-113.
DOI: 10.3923/ijv.2012.106.113
URL: https://scialert.net/abstract/?doi=ijv.2012.106.113
DOI: 10.3923/ijv.2012.106.113
URL: https://scialert.net/abstract/?doi=ijv.2012.106.113
INTRODUCTION
Potato (Solanum tuberosum L.) is considered one of the important vegetable crops in the most countries of the world. The potato crop ranks fourth in economic importance after Wheat, Maize and Rice and also the first alternative for grain crops and is used for human consumption, animal feed and as a source of starch, carbohydrates, alcohol and protein (Horton, 1992).
In Egypt, potato is considered as one of the most important and economic vegetable crops. It can be grow in almost any season and in most of country, except during very hot months in July and August. The cultivated area with potatoes during all seasons (summer, winter and nili) of 2009 on republic level reached was about 329721 feddans (feddan = 4200 m2) which yielded about 3659284 tons with an average of 11.098 tons/feddan. The first Governorate in Egypt in it's potato production and cultivated area is Behera Governorate with (66851 faddans and 759479 tons), the second Governorate is Monufyia with 43724 faddans and 448890 tons, the third Governorate is Noubaria with 38042 faddans and 397977 tons (Ministry of agriculture and land reclamation economic affairs sector).
Potato plants are subjected to attack by several viral diseases, causing harmful effects and reduce crop yield and tuber quality. More than 25 different viruses or virus diseases have been reported and all are pathogenic to potato (Beemstar and Rozendal, 1972).
Alfalfa mosaic virus is the type member of the genus Alfamovirus in the Bromoviridae family of plant viruses, with cryptogram (R/1: 1.1+0.8+0.7/16: U/U: S/C, Ve/Ap) is a world - wide distributed virus (Jaspers and Bos, 1980).
AMV has a very wide host range. This virus can naturally infected many herbaceous and some woody plant hosts (150 species in 22 families and transmissible to over 430 species of 51 dicotyledonous family (Xu and Nie, 2006).
In Egypt, AMV is one of the most important and widely distributed virus, appeared on potato plants in several locations at Egypt, causing severe lose (Shalaby, 1993; Gamal El-Din et al., 1994). PCR and RT-PCR assays (Saiki et al., 1988; Soliman, 2002) which involve the enzymatic amplification of DNA fragment defined by two oligonucleotide primers, have been used to diagnose a number of plant viruses belonging to several different groups (Henson and French, 1993). The RT-PCR or PCR method has been successfully utilized to detect viruses from infected potato tubers (Singh and Singh, 1995), from leaves (Lopez-Moya and Loez-Abella, 1992). Xu and Nie (2006) reported that RT-PCR used to identify of eight Canadian potato AMV isolates was confirmed by sequence analysis of their Coat Protein (CP) gene. The objective of this study was to established identification and molecular characterization of potato infecting isolate of Alfalfa mosaic virus in Egypt.
MATERIALS AND METHODS
A-isolation: The virus was isolated from infected potato plants growing under field conditions at different location in Minufyia Governorate, Egypt. Leaf samples showing calico and aucuba symptoms (doubted to be due to virus infection) were collected, examined serologically by ELISA (Clark and Adams, 1977) and the positive ones were homogenized in a mortar, after adding phosphate buffer (1:5 w/v, 0.1 M, pH 7.2, Xu and Nie, 2006), then the extracted sap was passed through a double layer of cheesecloth. The virus was purified biologically thought; the single local lesion technique as described by Khan and Monroe (1963) was followed. N. glutinosa L. plants were used as a local lesion host. Single local lesion was isolated, grinded in phosphate buffer, pH 7.2 and back inoculated onto N. glutinosa L. Finally, the resulting local concentric lesions were singly back inoculated onto potato plants. Inoculated potato plants were kept in the greenhouse and used as a source of infection in the following experiments.
B-mechanical transmission: Crude sap was extracted from the infected tissues by in phosphate buffer (1:5 w/v, 0.1 M, pH 7.2), (Xu and Nie, 2006), the leaves of healthy potato plants and indicator (N. glutinosa L., N. tabacum L., White burly and Chenopodium quinoa L.) pants, were first dusted with carborandum (600 mesh) and gently rubbed with a fresh prepared inoculum, using fore finger (Shalaby, 1993; Mervat, 1999; Xu and Nie, 2006).
C-rt-pcr test
Extraction of total RNA from plant tissues: Total RNA was isolated from the infected potato plants using RNA Purification Kit obtained from BioFlux according to manufacture’s instructions.
| Table 1: | Primer pairs designed for RT-PCR based on coat protein gene sequence AMV RNA 3 |
 | |
| aSequence location is based on the complete RNA 3 sequence of AMV- 425 M (GenBank accession No. K02703) (Barker et al., 1983) | |
Design and synthesis of the primers: The primer set was designed by Xu and Nie (2006) for RT-PCR amplification of AMV RNA based on the alignment of Coat Protein (CP) gene sequences of known AMV strains obtained from the NCBI website (GenBank). The primer set of AMV-F2 and AMV-R2 was designed to amplify the entire CP gene of AMV RNA 3 (Table 1).
cDNA synthesis: Two microlittere of RNA and 1.5 μL of 10 FM of the complementary primers (AMV-R2) were mixed in a sterile RNase-free microcentrifuge tube with nuclease-free water to a final volume of 15 μL. The tubes were heated to 70°C for 5 min, then cooled immediately on ice and spun briefly to collect the solution at the bottom of the tube. The following components were added to the annealed primer/template: 5 μL of 5X M-MLV reaction buffer [250 mM of Tris-HCl (pH, 8.3), 375 mM of KCl, 15 mM of MgCl2 and 50 mM of DTT (dithiothretol)], 2 μL of 10 mM deoxynucleoside triphosphates (dNTPs, 25 units of RNasi® ribonuclease inhibitor and 200 units of M-MLV RT enzyme (Promega) and nuclease-free water to final volume of 25 μL. the tubes were mixed gently by flicking the tubes and incubated for 60 min at 37°C.
Polymerase chain reaction (PCR): PCR amplification was performed in a final volume of 25 μL as the following: 2.5 μL of cDNA, 2.5 μL of 2.5 mM of dNTPs, 2.5 μL of 10X buffer, 2.5 μL of 25 mM MgCl2, 1 μL of each forward and reverse sense primer at 10 M, 0.2 μL Taq DNA polymerase and water.
Mixtures were incubated for 2 min at 94°C, followed by 35 cycles of 30 sec at 95°C, 30 sec at 58°C and 30 sec at 72°C, with a final incubation of 7 min at 72°C. PCR- amplified DNA fragments were separated by agarose gel electrophoresis in 1% agarose (Seakem LE, FMC) minigels in 0.5X TBE buffer pH, 8.0 using 100 bp DNA ladder. The gels were visualized with UV light after staining for 10 min with 10 g mL-1 of ethidium bromide and visualized with UV illumination using Gel Documentation System (Gel Doc 2000 BIO RAD). The expected size of the PCR product was 669 bp for AMV.
Cloning and sequencing of RT-PCR product: Amplified fragments from Egyptian isolate were extracted using Gel Extraction kit (Qiagen).
The PCR product was ligated into pGEM®-T Easy vector (Promega, USA) and the recombinant plasmids were introduced into E. coli strain DH5α as described by manufacturer’s instructions. DNA was prepared from selected white colonies, digested with EcoRI and fractionated on agarose gels using 1 Kb DNA ladder (Promega, USA).
The nucleotide sequence of clones having 669 bp insert were selected for dideoxy sequencing [ABI PRISMTM Dye Terminal Cycle Sequencing Ready Reaction Kit with AmpliTaq DNA Polymerase, FS (University of Wisconsin, Madison, Biotechnology Dept., USA) and performed on 377 XL automated DNA Sequencer (Applied Biosystem Co.). The nucleotide sequence was compared and analyzed using DNAMAN Sequence Analysis Software (Lynnon BioSoft, Quebec, Canada) with those of AMV isolates available in GenBank.
RESULTS
Virus isolation and propagation: Alfalfa mosaic virus was isolated from naturally infected potato plants (Solanum tuberosum L. cv Spunta) grown in Minufiya Governorate, Egypt. The infected plants showed yellow blotching and bright mottling of potato leaves (Calico or aucuba), (Fig. 1). Samples collected from potato plants were used to inoculate healthy potato plants and other indicator plants by mechanical inoculation under greenhouse conditions as described before and symptoms were observed daily for 30 days were recorded.
Mechanical inoculation: Results showed that AMV was readily mechanically inoculated by sap extracted from infected potato leaves. A calico symptoms were appeared on potato plants, chlorotic local lesions on N. glutinosa L., mosaic on N. tabacum L. White burly and necrotic local lesion on Chenopodium quinoa L. (Fig. 1).
Amplification of the RT-PCR: RT-PCR amplification of viral RNA was carried out on the total RNA isolated from infected potato plants. Electrophoresis analysis of RT-PCR product showed a single amplified fragment of 669 bp and no fragments were amplified from the RNA extracted from symptomless or healthy plants (Fig. 2).
Cloning of RT-PCR fragment into pGE®-T Easy vector: The pGEM®-T Easy Vector System is convenient system for the cloning of PCR products. This vector is characterized by adding a 3′terminal thymidine to both ends. These single 3′-T overhangs at the insertion site greatly improve the efficiency of ligation of a PCR product into the plasmids by preventing recircularization of the vector and providing a compatible overhang for PCR products generated by certain thermostable polymerases. This allows PCR inserts to ligate efficiently with the vector.
 | |
| Fig. 1(a-d): | Reaction of some plants to mechanical inoculation with the isolated virus: A. Calico and aucuba symptoms on Solanu tuberosum L. cv. Spunta. B. Chlorotic local lesions on inoculated Nicotiana glutinosa L. C. Necrotic local lesions on Chenopodium quinoa L. D. Mosaic on White Burly tobacco on Nicotiana tabaccum L. |
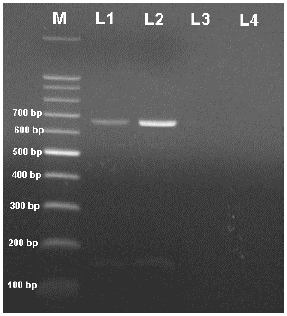 | |
| Fig. 2: | Agarose gel electrophoresis analysis of RT-PCR amplified products. M: 100 bp DNA ladder, L1, L2: Two potato plants infected with AMV; L3, L4: healthy plant sample as virus control |
Isolation of recombinant plasmids: Several white colonies resistant to ampicillin were selected to test for recombinant plasmids containing the AMV-PCR product. Restriction enzyme digestion with EcoRI will release the cloned gene only if inserted in the right direction.
Sequence analysis: Nucleotide sequencing of the RT-PCR amplified fragment in the recombinant plasmid was completed to determine if this RT-PCR fragment was AMV or not and to compare the sequence from this isolate with those of other AMV isolates available in GenBank. The nucleotide sequence of the Egyptian isolate of AMV was submitted in the GenBank under Accession No. HQ288892.
Multiple sequence alignment of the nucleotide sequence of the coat protein gene of AMV [Egyptian isolate (HQ288892)] with the corresponding sequences of eight different AMV isolates available in GenBank [New Zealand (AF215663); Italy (AJ130706); Canada (DQ314756); Australia (AF332998); Netherland (K03542); USA (AY340070); Brazil (FJ858265) and Korea (AF294433)] were analyzed using DNAMAN software (Fig. 3).
Sequence comparisons showed the percentage of similarity ranged from 97-98% of the eight reported isolates of AMV with the Egyptian isolate of AMV. The similarity of the nucleotide sequences suggested that the architecture of the alfalfa viruses is highly conserved.
DISCUSSION
Egypt is the most important of the Mediterranean countries produced and exported for the potato crop, where production and export of Egyptian potatoes in times where there are no fresh potatoes (harvest date) in Europe. It is also one of the main important vegetable crops in Egypt.
Under field conditions, potato plants are subjected to infection with many viruses (Dimitrova and Krachanova, 1979; Salazar, 1996).
Such viruses have been shown to cause great economic losses and considered to be the major limiting factors facing vegetative propagated plants specialty potatoes. Among such diseases Alfalfa mosaic virus was found to be widely distributed on potato plants.
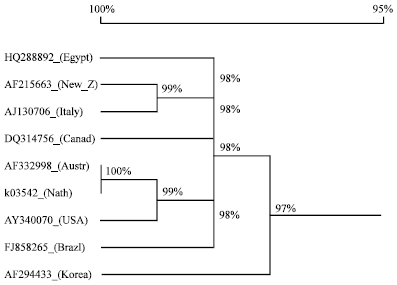 | |
| Fig. 3: | A phylogenetic tree showing relationships among reported isolates of AMV and the Egyptian isolate based on the nucleotide sequences. Horizontal distances indicate degree of relatedness |
This disease was investigated during the course of the present study.
Alfalfa mosaic virus was isolated from naturally infected potato plants (Solanum tuberosum L.) grown in different locations; this virus was isolated before from naturally infected potato in Egypt (Shalaby, 1993; Gamal El-Din et al., 1994). And also in different other countries by Dimitrova and Krachanova (1979), Xu and Nie (2006) and Pourrahim et al. (2007).
The isolated virus easily transmitted by mechanical means to different hosts. These results harmonized with those reported for AMV (Shalaby, 1993; Shafie et al., 1997; Mervat, 1999; Al-Shahwan, 2002; Xu and Nie, 2006; Jana et al., 2009; Parrella and Acanfora, 2010).
The present study demonstrated the successful use of RT-PCR to detect AMV in total nucleic acid extracts from infected potato plants, only one day is required for positive identification of the virus from infected tissue, amplified products for AMV was established as follows: The size of the major product from AMV-infected tissue was identical to that of the 669 bp from the CP gene of AMV; the specific primers did not amplify viral cDNA from extracts of uninfected potato plants.
Two AMV CP gene-specific primers sets (AMV-F/R and AMV-F2/R2) in this study, were designed and were highly specific, sensitive and useful in RT-PCR for detection and amplifying AMV CP gene sequence from total RNAs extracted from potato leaves.
Both primers (AMV-F/R and AMV-F2/R2) designed by Xu and Nie (2006) were specific for AMV RNAs and did not produce amplicons from potato leaves infected with several other viruses (Potato virus Y, Potato virus X and Potato leaf roll virus) or a viroid (Spindle tuber viroid). RT- PCR amplification of AMV RNA based on the alignment of Coat Protein (CP) gene sequences of known AMV strains obtained from the NCBI website (GenBank).
Amplified AMV-cDNA was cloned into pGEM®-T Easy Vector. The recombinant plasmid was successfully introduced into EcoRI (Soliman, 2002). Recombinant colonies which had correct insert (669 bp), were validated by plasmid minipreparation, followed by digestion of EcoRI restriction enzyme. Colonies were identified and shown to contain insert with the expected molecular weight size (Barker et al., 1983).
Sequence analysis of nucleotides and amino acids showed that this isolate (Egyptian isolate) was closely related to each other isolates and there were only slight differences in both nucleotides and amino acid sequences in their CP gene. Sequence comparisons showed the percentage of similarity ranged from 97-98% of the eight reported isolates of AMV with the Egyptian isolate of AMV. The similarity of the nucleotide sequences suggested that the architecture of the alfalfa viruses is highly conserved.
AMV isolate was detected in potato plants in Egypt and characterized at the biological, serological and molecular level in this study and their genetic relationships with other known AMV strains were established. RT- PCR protocol was developed for the detection of AMV in potato and the confirmation of PCR results.
REFERENCES
- Barker, R.F., N.P. Jarvis, D.V. Thompson, L.S. Loesch- Fries and T.C. Hall, 1983. Complete nucleotid sequence of Alfalfa mosaic virus RNA3. Nucleic Acids Res., 11: 2881-2891.
Direct Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Henson, J.M. and R. French, 1993. The polymerase chain reaction and plant disease diagnosis. Ann. Rev. Plant Pathol., 31: 81-109.
Direct Link - Parrella, G., N. Acanfora and M.G. Bellardi, 2010. First record and complete nucleotide sequence of Alfalfa mosaic virus from Lavandula stoechas in Italy. Plant Dis., 94: 924-924.
CrossRefDirect Link - Pourrahim, R., S. Farzadfar, A.R. Golnaraghi and A. Ahoonmanesh, 2007. Incidence and distribution of important viral pathogens in some Iranian potato fields. Plant Dis., 91: 609-615.
CrossRefDirect Link - Saiki, R., D.H. Gelfand, S. Stoffel, S.J. Scharf and R. Higuchi et al., 1988. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 239: 487-491.
CrossRefPubMedDirect Link - Singh, M. and R.P. Singh, 1995. Digoxigenin-labelled cDNA probes for the detection of potato virus Y in dormant potato tubers. J. Virol. Methods, 52: 133-143.
CrossRef - Xu, H. and J. Nie, 2006. Identification, characterization and molecular detection of Alfalfa mosaic virus in potato. Phytopathology, 96: 1237-1242.
CrossRefDirect Link