
Dalia M. Omar
The Central Laboratory for Evaluation of Veterinary Biologics (CLEVB), El Abassia, Cairo, Egypt
Elham A. El-Ibiary
The Central Laboratory for Evaluation of Veterinary Biologics (CLEVB), El Abassia, Cairo, Egypt
A.S. Sadik
Department of Agriculture Microbiology (Virology Laboratory), Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Mamdouh H. Abdel-Ghaffar
Department of Agriculture Microbiology (Virology Laboratory), Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Badwy A. Othman
Department of Agriculture Microbiology (Virology Laboratory), Faculty of Agriculture, Ain Shams University, Cairo, Egypt
International Journal of Virology
Year: 2011 | Volume: 7 | Issue: 4 | Page No.: 123-134







ABSTRACT
In mid-February 2006, an outbreak of Highly Pathogenic Avian Influenza (HPAI) H5N1 affected the commercial poultry production sector and backyards in Egypt and resulted dramatic economic losses to the poultry industry of Egypt and continue to pose a serious threat to public health. The present study was designed for detection and isolation of Avian Influenza Viruses (AIV) circulating among poultry since their first detection in 2006. Tracheal and cloacal swabs were taken from the freshly dead birds. Till now (2010) the tested swab samples were inoculated into the allantoic cavity of 9-11-day-old SPF Embryonated Chicken Eggs (ECE) for virus isolation. The allantoic fluids (AF) of the inoculated ECE were examined for detection of Avian Influenza (AI) isolates using rapid haemagglutination test (HA). It was found that, all the inoculated isolates caused high mortalities up to 100% for the embryos 24-48 h post inoculation and gave very strong RBCs agglutination. The haemagglutination positive samples were identified and subtyped antigenically by serological tests using Haemagglutination Inhibition test (HI) using standard AI antisera and genetically by RT-PCR using specific primer. All the isolates were confirmed to be H5N1 and grouped according to the year of isolation as 2006, 2007, 2008, 2009 and 2010 group isolates. The HA titers of the above mentioned isolates were 6, 7, 8, 7 and 7, respectively. All the isolated AI viruses were titrated in ECE and examined for determination of their pathogenicity in Specific Pathogen Free (SPF) 4-6 week old chicken. All the AI isolates proved to be HPAI viruses where the Intravenous Pathogenicity Index (IVPI) score for them were 2.1, 2.5, 2.3, 2.2 and 2.3 and their titers in ECE were 10.1, 9, 9.3, 9.3 and 10 Log10 EID50/mL for 2006, 2007, 2008, 2009 and 2010 isolates, respectively.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: August 30, 2011;
Published: October 22, 2011
How to cite this article
Dalia M. Omar, Elham A. El-Ibiary, A.S. Sadik, Mamdouh H. Abdel-Ghaffar and Badwy A. Othman, 2011. Serological and Molecular Identification of Some Isolated Avian Influenza Viruses During Outbreaks in Egypt. International Journal of Virology, 7: 123-134.
DOI: 10.3923/ijv.2011.123.134
URL: https://scialert.net/abstract/?doi=ijv.2011.123.134
DOI: 10.3923/ijv.2011.123.134
URL: https://scialert.net/abstract/?doi=ijv.2011.123.134
INTRODUCTION
Avian Influenza (AI) is a highly contagious notifiable disease of birds caused by the influenza viruses that are members of the family Orthomyxoviridae (Lamb and Krug, 1996). Most influenza pandemics are associated with the type A viruses (Flemming et al.,1995) that are classified into various subtypes based on the antigenic differences between two surface glycoproteins; the hemagglutinin (16 HA subtypes) and neuraminidase (9 NA subtypes) (Easterday et al.,1997). Also, AI viruses can be divided into two distinct groups based on their ability to cause disease. The first group is Highly Pathogenic Avian Influenza (HPAI) that cause severe disease in domestic poultry which led to the culling of approximately 200 million birds worldwide, inflecting major economic damage to the poultry industry (Flemming et al., 1995; Cattoli et al., 2004; Check, 2006). The second group is Low Pathogenic Avian Influenza (LPAI) that produces a mild disease with respiratory signs, depression, egg production problems and low mortality rates. However, in some cases, this infection may cause significant mortality rates, generally when it occurs in a combination with other bacterial or viral infections (Cattoli et al., 2004), All 16 AI subtypes can cause LPAI disease in susceptible birds, while, the HPAI disease is restricted to strains belonging to H5 and H7 subtypes.
The recent years, the pathogenic H5N1 subtype crossed the species barrier to infect humans as first reported in Hong Kong in 1997 (Chan, 2002) and in other mammals (Keawcharoen et al., 2004). Since 2003, H5N1 strains have spread from Asian to European countries, then to the Middle East and Africa (Ducatez et al., 2006, 2007).
In mid-February, 2006 a devastating HPAI H5N1 infected the commercial poultry production sectors and backyards in Egypt (Aly et al., 2006a, b), where the authorities do all their hazard efforts in order to minimize the huge economic losses, besides some human deaths in trial to stamp out the disease. Two months later and after the failure in disease stamping out, the authorities directed to use the long strategy with using of both homologous and heterologous vaccine but without regulation. Several repeated potential H5N1 outbreaks still occurred in vaccinated and non vaccinated birds in both backyard and commercial farms with continuous reporting of human cases and still considered a renewable problem warning with a possible endemic.
A global effort is underway to control or eradicate H5N1 in poultry and prevent human exposure, both of which may also reduce the risk of pandemic emergence. The scientific rationale for these programs is provided by ecologic, virologic, epidemiologic and immunologic studies. In particular, molecular and functional characterization of H5N1 viruses from poultry will help inform development and implementation of public health control measures involving diagnosis, immunization and antiviral drug therapy.
In order to investigate the origin of H5N1 viruses isolated in Egypt and to identify of avian influenza virus within the country, this research tried to characterize H5N1 isolates from different regions of poultry populations using the tracheal and cloacal swabs of infected chickens collected in period between 2006 and 2010 hoping that it will help the authorities and provide a comprehensive approach towards more efficient disease control and prevention for both poultry and humans.
MATERIALS AND METHODS
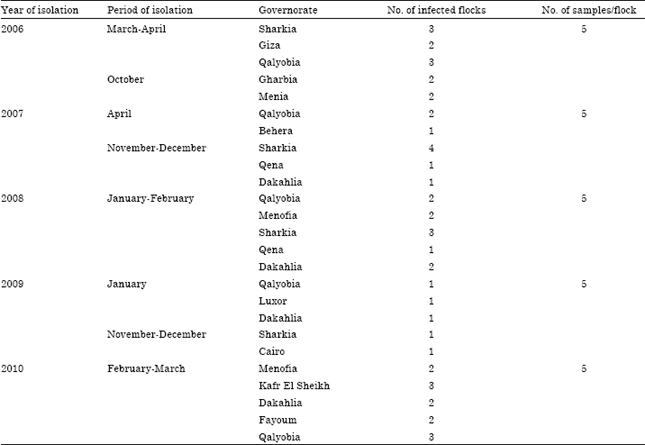
Case history of HPAI H5N1 in Egypt: During the outbreaks of AI in Egypt since 2006 till 2010, large number of flocks either broiler, layer and breeders all over the country, suffered from major clinical signs of AI outbreaks. The clinical examination was performed at the site. Fresh samples from each infected flock all over the country were used for diagnosis and detection the AI virus as shown in Table 1.
| Table 1: | A collective data on the samples collection in Egypt during period between February 2006 and July 2010 |
 | |
Sample management and preparation
Sample collection: Fresh samples of dead and sick birds from each infected flock were collected. These samples were stored on ice during transportation that never exceeds one day. In the laboratory, tracheal and cloacal swabs were taken on virus transport medium (tryptose phosphate broth with antibiotics). Samples were stored at -80°C until processed (Johnson, 1990).
Sample processing: The swab samples were clarified at 3000 g for 30 min and then supernatants were treated with multi-antibiotic mixture for AIV isolation as described by Lee and Suarez (2004).
Virus isolation: Methods: A volume of 0.2 mL from each specimen supernatant was inoculated into five 9-11-day-old Specific Pathogen Free (SPF), Embryonating Chicken Eggs (ECE) via the allantoic sac route according to the methods previously described by Payungporn et al. (2006). The mortality% of the inoculated ECEs was estimated. The Allantoic Fluids (AF) of died ECEs were collected and tested for the presence of Haemagglutinating (HA) activity.
Rapid HA tests: It was done according to Swayne et al. (1998). This method is based on the reaction between HA activity of virus and RBCs. The HA activity can be visualized by adding a drop of AF to a drop of 10% RBCs suspension. The resulting reaction is macroscopically visible.
Identification and characterization of AIV
Micro plates HA and HI tests (serological identification): A panel of standard positive sera against ND, EDS and AI-H5 both H5N2 (A/Chicken/Mex./232/94) and H5N1 (A/Goose/Guangdong/1/96) viruses were used (Central Veterinary Laboratory (CVL), Weibrige). The tested viruses were diluted in 2 fold serial dilution and 1% RBCs were added to determine the Haemagglutination (HA) titer of each virus. Each virus must be standardized to a concentration of 4 HA unit/50 L prior to testing with HI assay. All the standard positive antisera were diluted in 2 fold dilution and mixed with an equal volume of 4 HA unit antigens of each virus. After incubation, RBCs 1% suspension were added and the dilutions were examined for inhibition of the HA activity. The test was conduct also against negative reference serum. It was done according to OIE (2008).
In vivo pathogenicity test: The test was used to classify the H5 subtype of AI virus isolates as HP or LP for chickens based on the OIE definition. The virus is classified as HP if it kills ≥75% of 4-8 week old susceptible 6-week-old SPF chickens within 10 days when inoculated with the tested isolate using I/V route. The results were assessed as Intravenous Pathogenicity Index (IVPI). So, the IVPI is defined as the mean score per bird per observation over the 10-day period. An index of 3 means that all birds died within 24 h while an index of 0 means that no bird showed any clinical signs during the 10 days observation period. The test was done according to Terregino and Capua (2009).
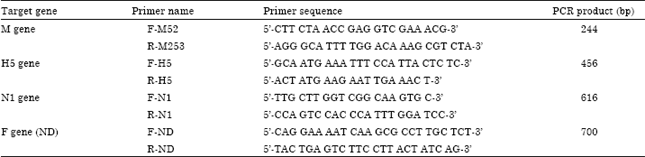
RT-PCR (genetic identification): The harvested AF was tested by RT-PCR following the same protocol described in a published paper of Khan et al. (2009). The primary application of RT-PCR is the identification of the AI isolates using a set of primers targeted to the conserved Matrix (M) gene. After that, the test was used for sub-typing of the AI viruses depending on specific primers directly targeted to the HA and NA subtypes (H5N1) as shown in Table 2. The technique was started with the RNA extraction from the collected samples using RNA Extraction kit (EZ-10 Spin column viral RNA Mini-preps Kit, Biobasic INC., Canada, Cat. # VT82112) and followed by RT-PCR (M-MULV Reverse Transcriptase Kit, sib Enzyme Ltd. Cat. # E317 and PCR system Kit, Biobasic Co., Canda. Cat. # D0088) producing large numbers of DNA molecules copies. The amplified PCR products were detected and analyzed by 1% agarose gel electrophoresis.
| Table 2: | Oligo-nucleotide primers for AI identification and HA sub-typing PCR |
 | |
 | |
| Fig. 1: | Electrophoretogram of AI PCR products |
Also, set of specific primers against F gene of ND (Table 2) were used in the genetic identification of the isolated viruses.
DNA sequencing of H5 gene: Sequencing of PCR amplicons was performed using specific primers for the conserved part of HA1 of H5 gene of each AI isolates. The identities of these PCR products were then confirmed by sequence analysis and comparison between sequences derived for this study and sequences posted in GenBank for other Egyptian and standard vaccinal AI strains using BLAST search. In addition, amino acid sequence alignments were used to infer amino acid substitutions between the isolates analyzed in Fig. 1.
Virus propagation and titration: It was done according to Woolcock et al. (2001), the AF containing infectious virus of each sample were diluted 10 fold serial dilution using PBS up to 10-4 for virus propagation and 10-15 for virus titration. SPF ECE 9 to 11-day old was inoculated by allantoic sac route with each dilution. For propagation, AF from haemagglutination positive egg was pooled and aliquoted for long term storage at -70°C. For titration, the dead and live egg per dilution were examined by slide HA test the after observation period and calculate the titer as 50% Egg Infectious Dose (EID50).
RESULTS
H5N1 infection in the poultry: It was reported that all the infected flocks, suffered from severe clinical signs, had high percentage of mortality up to 100% within 3-4 days from the onset of first clinical signs. Listlessness, recumbency, anorexia, reduced activity, reduction in “in-house” noise, dehydration, decreased feed and H2O intake, drop in egg production in breeders and layers leading to total cessation followed by nervous signs, diarrhea may be evident as bile or ureate stained loose droppings with variable amount of intermixed mucus. Cyanosis of the wattle and comb, petechial hemorrhages on the hock, peri-orbital oedema and conjunctivitis were observed. Sudden death occurs in a recombinant position.
Virus isolation: The inoculated ECE were examined and embryo mortality was recorded (Table 3).
| Table 3: | Results of virus isolation of different tracheal and fecal swabs collected from severe infected poultry flocks |
 | |
| Table 4: | Results of the antigenic and genetic Identification of AI isolates |
 | |
| Table 5: | Results of the characterization of AI isolates |
 | |
Embryo deaths occurred as early as 18-24 h post inoculation and all samples showing complete positive haemagglutination. NB.: All the AI samples which isolated from different flocks during specific period of time would be gathered together under large group and named as 2006, 2007, 2008, 2009 and 2010.
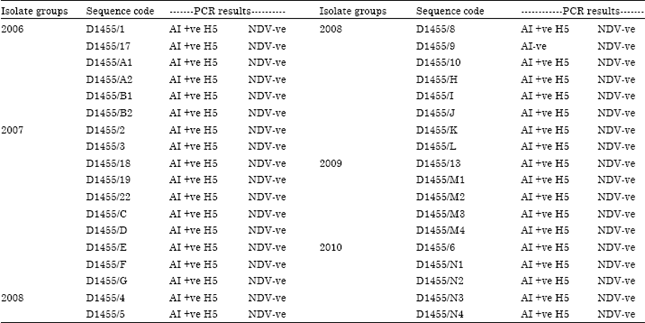
Identification and characterization of AIV: The data from Table 4 showed that all isolate groups were H5-AIV positive while they were ND and EDS negative. Also, the results of RT-PCR test for M, H genes (Table 4, 6) confirmed that isolates were H5N1 AIV positive where they yielded a strong positive reaction as shown in Fig. 1. While RT-PCR using F gene primers proved that all the isolated viruses were ND negative (Table 6).
The HA titer for each AI group were 6, 7, 8, 7 and 7 for the 2006, 2007, 2008, 2009, 2010 AIV groups respectively as shown in Table 5. While, the IVPI value of the same AI groups ranged from 2.1-2.3. So, all of them considered to be a highly pathogenic AIV for chicken (Table 5). On the other hand, the AIV titer was expressed as EID50 and ranging from 109-1010.1 for all AI isolate groups as shown in Table 5.
The genetic analysis of the isolated
AI viruses: It was shown from phylogenetic tree (Fig. 2) and Table 7 that: The AI isolates from 2006 were identical with each other and fell into the Egyptian group B reference strains with about 100% identity with them.
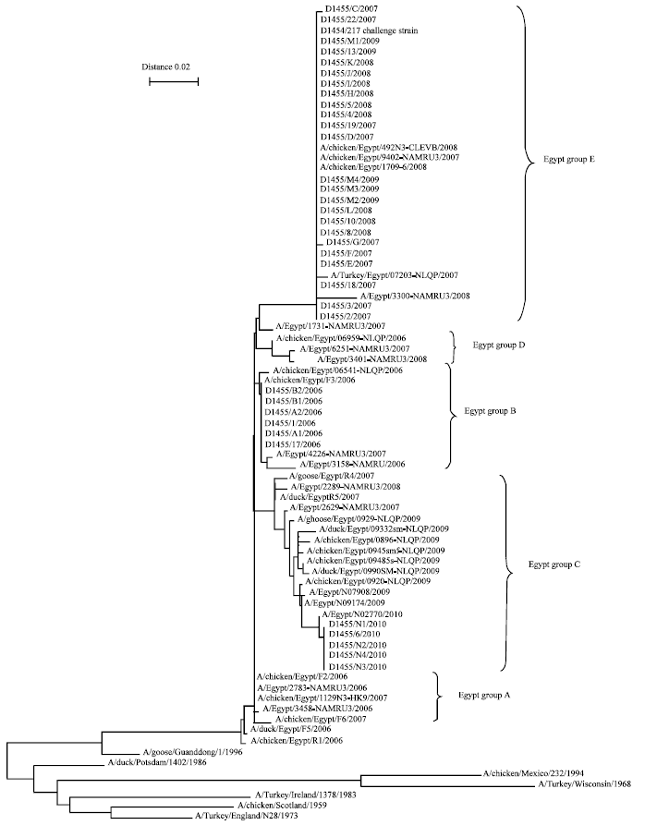 | |
| Fig. 2: | Phylogenetic analysis of the AI |
| Table 6: | Results of the genetic identification and subtyping of the isolated viruses by RT-PCR using specific primers |
 | |
| Table 7: | Sequence identity matrix of Egyptian AIV isolates |
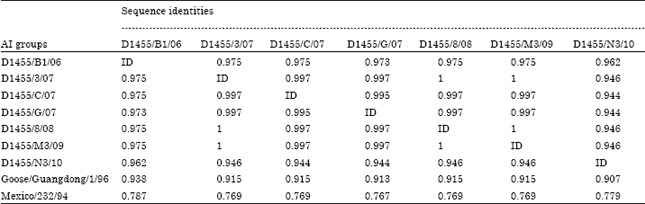 | |
The AI isolates from between 2007-2009 were grouped together with Egyptian group E reference strains with about 98-99.5% identity with them.
The AI samples from 2010 were identical with each other and showed the closest relation to the Egyptian group C reference strains with about 98% identity with them.
It was found that 2006 isolates were identical to the 2007, 2008, 2009 groups with 97.5% and to 2010 group with 96.2%. While, the degree of identity between 2010 isolates and 2007, 2008, 2009 was 94.5%.
The deduced amino acid sequence of HA gene of the 2007, 2008, 2009 AI isolates revealed substitutions of about 10 amino acids with 2006 strains and about 13 amino acids with the 2010 isolates as shown in Table 8.
DISCUSSION
Since the initial outbreaks of HPAI H5N1 in 1997 (Claas et al., 1998), the virus has diverged both antigenically and genetically with new genotypes and variant viruses continuing to emerge (Wan et al., 2007).
| Table 8: | The differences in nucleotide and amino acid sequences of partial HA gene of Egyptian AI H5N1 strains isolated in 2007, 2008 and 2009 in comparison to the 2006 and 2010 groups |
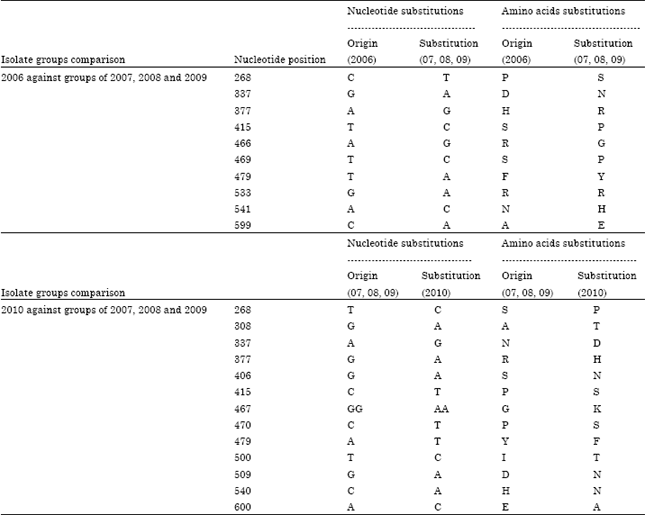 | |
Similar to that of seasonal influenza viruses, the continuous and rapid evolution of those HPAI viruses necessitates reviewing and updating for vaccine development. Also, continuous monitoring of virus evolution and genetic changes of H5N1 and other important avian influenza viruses with epizootic potential is critical to detect genetic drift or a genetic shift that may lead to new outbreaks or a future influenza pandemic. In order to slow the rate of mutation and reassortment of viruses circulating in poultry populations, complete eradication by culling or culling with vaccination are commonly used (Marangon et al., 2008).
Sample selection and handling are important in the detection of the AI virus. The type and quality of samples collected will determine the success of diagnosis. In addition, storage conditions from the time of collection until the specimen is processed in the laboratory are very important for the diagnosis of AI, this agreed with Johnson (1990). Swabs collected for AI surveillance should be taken from clinically ill or dead animals where the virus is present in the highest quantity at this time.
Virus Isolation (VI) techniques is considered the reference standard for the diagnosis of AI virus in the collected samples during a period of 2006-2010 in Egypt. This was agreed with Swayne et al. (1998), who told that virus isolation is the best test method to accurately identify the presence of AIV from tracheal and cloacal swab samples. VI is necessary to confirm the presence of virus in an index case and to perform further characterization of the virus. ECE is the most widely used and sensitive host system for the isolation and propagation of poultry adapted AIV (Woolcock et al., 2001).
Because of high sensitivity of VI, this method may be used to detect AI virus during any stage of an active infection. So, AI maybe detected within 24 h of infection in an individual bird and for several weeks post exposure in a flock (Swayne and Halvorson, 2003).
Additional tests as HA assay on fluids from eggs inoculated with the tested samples was applied to confirm presence of AI virus. The positive sample for HA was then tested by HI assay to differentiate AI virus from other haemagglutinating viruses like ND and EDS (Beard, 1980). Once avian paramyxoviruses and adenoviruses had been ruled out, the presence of AI virus can be confirmed by type specific HI test using AI virus-specific subtyping serums for H5 antigens (reference H5N1 and H5N2 antisera). This agreed with Webster et al. (2002) who said that VI is very sensitive, buts not highly specific or selective because other viruses that maybe present in poultry samples will grow in chicken embryos.
All the isolated Egyptian strains (2006-2010 groups) of AIV were also tested for determination their virulence for chicken. The IVPI of the AI isolates were range from 2.1-2.3 indicating that they were classified as HP for chicken. This was agreed with Terregino and Capua (2009), who mentioned that any influenza A virus, regardless of subtype, yielding a value >1.2 in an IVPI test is considered to be highly pathogenic avian influenza virus. Also, it was found that, all the Egyptian isolates had a titer ranged from 109-1010.1 EID50 and HA titer ranged between 6-8 log2.
Also RT-PCR technique using specific set of primers against H5 and N1 genes of AIV was successful in analyzing samples and confirming the presence of H5N1 AI.
This was agreed with Lee et al. (2001), who said that the application of molecular methods for the detection of viral nucleic acid has become an important, high speed and low cost tool for detection of AI virus type A and directly targeted to the different HA and NA subtypes.
Phylogenetic analysis of the HA gene of strains analyzed for this study showed five distinct subgroups (Fig. 1). In late 2007, 2008 and 2009 a new variant (group E) appeared and was clearly distinct from 2006 and early 2007 Egyptian strains (group B) as in Fig. 2. The HA gene of the variant strains in that subgroup had at least 10 different amino acid mutations from 2006 Egyptian H5N1 and 13 different amino acid mutations from 2010 strains sequenced from infected poultry flocks (Table 8). Similar findings of the emergence of possible vaccine escape variants were reported in Egypt one and half year after the implementation of vaccination in poultry (Balish et al., 2010).
Finally, the results suggest an endemic situation of AI in Egypt. New outbreaks of H5N1 occurred in the different years from 2006-2010 indicating that the virus is circulating in the region. The traditional poultry breeding in Egypt makes the spread of H5N1 strain difficult to control.
So, the emergence of these variant strains and their spread in a short period of time to several governorates in Egypt and its considerable antigenic variation from previous Egyptian isolates emphasizes the need for continuous monitoring of genetic and antigenic changes in HPAI H5N1 as an early warning system of the detection of new variants and faster response to contain disease spread in the future.
REFERENCES
- Balish, A.L., C.T. Davis, M.D. Saad, N. El-Sayed and H. Esmat et al., 2010. Antigenic and genetic diversity of highly pathogenic avian influenza A (H5N1) viruses isolated in Egypt. Avian Dis., 54: 329-334.
CrossRefDirect Link - Cattoli, G., A. Drago, S. Maniero, A. Toffan and E. Bertoli et al., 2004. Comparison of three rapid detection systems for type A influenza virus on tracheal swabs of experimentally and naturally infected birds. Avian Pathol., 33: 432-437.
PubMed - Chan, P.K.S., 2002. Outbreak of avian influenza A (H5N1) virus infection in Hong Kong in 1997. Clin. Infect. Dis., 34: S58-S64.
CrossRef - Claas, E.C.J., A.D.M. Osterhaus, R. van Beek, J. de Jong and G.F. Rimmelzwaan et al., 1998. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet, 351: 472-477.
Direct Link - Ducatez, M.F., C.M. Olinger, A.A. Owoade, S. de Landtsheer and W. Amemerlaan et al., 2006. Avian flu: Multiple introduction of H5N1 in Nigeria. Nature, 442: 37-37.
CrossRef - Ducatez, M.F., Z. Tarnagda, M.C. Tahita, A. Sow, S. de Landtsheer et al., 2007. Genetic characterization of HPAI (H5N1) viruses from poultry and wild vultures, Burkina Faso. Emerg. Infect. Dis., 13: 611-613.
CrossRefDirect Link - Johnson, F.B., 1990. Transport of viral specimens. Clin. Microbiol. Rev., 3: 120-131.
PubMedDirect Link - Keawcharoen, J., K. Oraveerakul, T. Kuiken, T. Fouchier and A. Amonsin et al., 2004. Avian influenza in tigers and leopards. Emerg. Infect. Dis., 10: 2189-2191.
PubMed - Lee, M.S., P.C. Chang, J.H. Shien, M.C. Cheng and S.K. Shieh, 2001. Identification and subtyping of avian influenza viruses by reverse transcription PCR. J. Virol. Methods, 97: 13-22.
Direct Link - Lee, C.W. and D.L. Suarez, 2004. Application of real-time RT-PCR for the quantification and competitive replication study of H5 and H7 subtype avian influenza virus. J. Virol. Methods, 119: 151-158.
CrossRef - Marangon, S., M. Cecchinato and I. Capua, 2008. Use of vaccination in avian influenza control and eradication. Zoonoses Pub. Health, 55: 65-72.
PubMed - Payungporn, S., S. Chutinimitkul, A. Chaisingh, S. Damrongwantanapokin and C. Buranathai et al., 2006. Single step multiplex real-time RT-PCR for H5N1 influenza A virus detection. J. Virol. Methods, 131: 143-147.
CrossRef - Wan, X.F., G. Chen, F. Luo, M. Emch and R. Donis, 2007. A quantitative genotype algorithm reflecting H5N1 avian influenza niches. Bioinformatics, 23: 2368-2375.
CrossRef - Woolcock, P.R., M.D. McFarland, S. Lai and R.P. Chin, 2001. Enhanced recovery of avian influenza virus isolates by a combination of chicken embryo inoculation methods. Avian Dis., 45: 1030-1935.
PubMed