
A. E. Aboul-Ata
Plant Virus and Phytoplasma Research Section, Plant Pathology Research Institute, Agriculture Research Center, 12619 Giza, Egypt
G. Anfoka
Department of Biotechnology, Faculty of Agricultural Technology,Al-Balqa` Applied University, Al-Salt 19117, Jordan
M. Zeidan
Molecular Genetics and Virology, Al-Qassmi Research Center,Baqa El-Gharbia 30100, Israel
H. Czosnek
Institute of Plant Sciences and Genetics in Agriculture,Robert H. Smith Faculty of Agriculture, Food and Environment,The Hebrew University of Jerusalem, Rehovot 76100, Israel
International Journal of Virology
Year: 2010 | Volume: 6 | Issue: 3 | Page No.: 126-137







ABSTRACT
Middle Eastern countries are major consumers of small grain cereals. For example, Egypt is the biggest bread wheat importer with 5.9 Million Tons (MT) although, it itself produces 10.5 MT. Jordan and Israel import almost all the grains they consume. Viruses are the major factors that impair production in Middle East. They are transmitted in non persistent, semi persistent and persistent manners by insects (aphids, leafhoppers and mites) and through soil and seeds. Hence, there is a need to control insect-borne cereal viruses not only in the field but also through plant quarantine services for imported seed-and soil-borne viruses. Viruses need to be controlled in the frame of regional collaborative activities involving the Middle Eastern countries. The means to be used to diagnose cereal viruses may include symptom observation, immunological technologies such as ELISA using polyclonal and monoclonal antibodies against the virus coat protein (raised against purified virions or against the virus capsid protein expressed in bacteria or in yeast), molecular techniques such as PCR (uniplex and multiplex), RFLP, SSCP and microarrays. In this study, we explore the different diagnosis, typing and detection techniques of cereal viruses available to the Middle Eastern countries and we review the ongoing collaborative research projects.
PDF Abstract XML References Citation
Received: April 03, 2010;
Accepted: May 17, 2010;
Published: September 07, 2010
How to cite this article
A. E. Aboul-Ata, G. Anfoka, M. Zeidan and H. Czosnek, 2010. Diagnosis of Cereal Viruses in the Middle East. International Journal of Virology, 6: 126-137.
DOI: 10.3923/ijv.2010.126.137
URL: https://scialert.net/abstract/?doi=ijv.2010.126.137
DOI: 10.3923/ijv.2010.126.137
URL: https://scialert.net/abstract/?doi=ijv.2010.126.137
INTRODUCTION
Cereals such as wheat, corn and rice are strategic crops, whereas, barley and sorghum are essential crops. Middle Eastern countries, as developing countries, suffer from insufficient production of wheat, rice and corn and therefore, need to rely on importations to cover the national needs. Wheat for instance, in 2007, Egypt produced 7.4 Million Tons (MT) and imported 5.9 MT. Israel produced 0.16 MT and imported 1.2 MT. Jordan imported 0.8 MT, almost all its needs. More information can be found in the FAO website(http://faostat.fao.org/site/342/default.aspx for imports; http://faostat.fao.org/site/339/default.aspx for production).
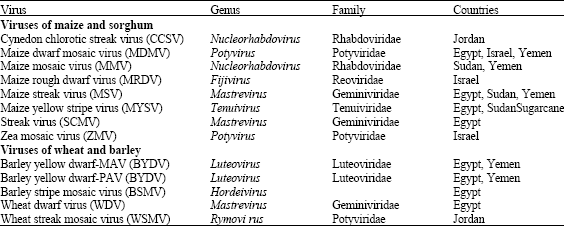
Viruses threaten cereal production i.e., Aboul-Ata and Ammar (1988) proved that Maize Yellow Stripe Tenuivirus (MYSV) can cause 80% losses in white maize and cause 70% in sweet corn when severe infection. Middle East had several outbreaks have been recorded in the Middle East (Ammar et al., 1988, 2008) as well as in neighboring European and Asian countries (Jones, 2009). Although, sixty-six viruses are able to infect grasses and cereal crops (Lapierre and Signoret, 2004), only a few of them causes economically important yield loss. The major cereal viruses in the Middle East are listed in Table 1. Symptoms caused by some of those viruses are shown in Fig. 1 and 2. The five cereal viruses with the highest impact in the Middle east are Wheat dwarf virus (WDV) (genus Geminivirus), Barley stripe mosaic virus (BSMV) (genus Hordeivirus) which is seed-borne, Wheat streak mosaic virus (WSMV) (genus Rymovirus) which is complexes with High plains virus in infected seeds (Ulman et al., 1997) and Barley yellow dwarf (BYDV) PAV, MAV, RMV and SGV strains-(genus Luteovirus) and Cereal yellow dwarf virus (CYDV-RPV ) (Genus Plerovirus) (Ammar et al., 2008; Aboul-Ata et al., 1992). WDV is a single-stranded DNA virus; BSMV, WSMV, BYDVs and CYDVs are single-stranded RNA viruses. BYDV was first identified on barley (Oswald and Houston, 1951). WSMVwas observed and identified in the early 1950s (McKinney, 1953). The presence of WDV was detected relatively late, at the end of the 1980s (Szunics et al., 2000).
Cereals are also infected by economically less important viruses such as Agropyron mosaic virus (AgMV) and Ryegrass mosaic virus (RyMV) (both belonging to the genus Rymovirus). Rice stripe necrosis virus (RSNV) (genus Furovirus), Maize streak virus (MSV) (genus Geminivirus), Rice yellow mottle virus (RYMV) (genus Sobemovirus), Maize rough dwarf virus (MRDV) (genus Fijivirus) and Brome mosaic virus (BMV) (genus Bromovirus) (Lapierre and Signoret, 2004).
Virus outbreaks can have dramatic effects. During the summer of 1989 and 1999, MYSV caused 80-90 yield loss in Egyptian maize (Ammar et al., 2008). Many of the corn fields have been eradicated because of large-scale-severe MYSV infection. The Middle Eastern countries never adopted breeding program for virus resistance. For instance Egypt has a strong cereal breeding program but does not have a program for virus resistance (Annual field book, Field Crop Research Institute, ARC, in Arabic). Diagnosis is the first step in the attempt to control virus infection. The variable epidemiological behavior of the multiple virus genera and families that affect cereals, together with the diversity in sequences of isolates from a same virus, complicates greatly control efforts.
| Table 1: | Viruses infecting maize, sorghum, wheat and barley in Middle Eastern countries (Ammar et al., 2008) |
 | |
 | |
| Fig. 1: | Viruses infecting maize in the Middle East: symptoms and vectors. (a) Symptoms of Maize streak virus MSV, (b) symptoms of Maize yellow stripe virus MYSV, (c) Cicadulina chinai vector MYSV, (d) symptoms of Maize dwarf mosaic virus MDMV, (e) symptoms of Sugarcane mosaic virus SMV, (f) symptoms of Maize mosaic virus MMV, (g) Peregrinus maidis vector of MMV, (h) and (i) symptoms of Maize rough dwarf virus (MRDV), (j) Maize vein enation disease caused by toxic-saliva of leafhopper vector (Ammar et al., 2008) |
Here, we discuss BYDV as an example of the seventeen recorded viruses infecting cereals in the Middle East. This study is aimed at presenting methodologies for cereal virus detection, typing and sub-typing as epidemiology and outbreak-causing factors and at exploring means of control using breeding programs for virus resistance. We wish to convince decision makers to list virus-causing problems as high priority for control, to encourage research institutes do develop improved detection means and to persuade breeders to initiate breeding programs for virus resistant varieties that framers will be willing to use. Regional organized scientific collaboration in Middle East area, for getting advanced technology exchange, will assist viral disease widespread follow up and control.
 | |
| Fig. 2: | Viruses infecting wheat and barley in the Middle East: symptoms and vectors. (1) Symptoms of Wheat streak mosaic virus (WSMV), (2) the mite vector, (3) symptoms of Wheat dwarf virus WDV, (4) symptoms of Barley stripe mosaic virus BSMV, (5) symptoms of Barley stripe mosaic virus BSMV (Ammar et al., 2008) |
Viral-Infection Diagnosis, Detection and Typing
Nomenclature
A group of luteoviruses called Barley yellow dwarf viruses (BYDVs) produce symptoms on wheat, barley and other grain cereals characterized by yellowing of leaves starting from the leaf tip and expanding to the base, stunting and small spike formation (Fig. 1 and 2). BYDV was first reported in the early 1950s (Oswald and Houston, 1951). BYDVs are transmitted by aphids in a persistent, circulative, manner but not mechanically and not through seeds (D’Arcy and Burnett, 1995). Aphids acquire and transmit BYDVs while feeding on the phloem sieve tube elements of host plants (Power and Gray, 1995). The five strains and their principal vectors, in parenthesis, are: BYDV-RPV (Rhopalosiphum padi), BYDV-RMV (R. maidis), BYDV-MAV (Sitobion avenae), BYDV-SGV (Schizaphis graminum) and BYDV-PAV (R. padi, S. avenae and others). BYDV-PAV is the major strain infecting wheat in Egyptian wheat (Aboul-Ata et al., 1992; Lister et al., 1994). It is also dominant in the Middle East (Syria, Jordan) and in the Maghreb (Tunisia, Algeria and Morocco). Transmission depends on specific interactions between virus and insect proteins (Li et al., 2001; Andret-Link and Fuchs, 2005). This nomenclature system has been adopted by all BYDV researchers. These strains are also distinguishable serologically. The International Committee on the Taxonomy of Viruses (ITCV) has divided the BYDVs into two distinct virus groups: BYDV (PAV, RMV, SGV and MAV) and Cereal yellow dwarf polerovirus (CYDV) (RPV) (D'Arcy et al., 2000).
Diagnostic and Detection Methodologies Used in the Middle East Area
Serological Means
Different direct and indirect Enzyme-Linked Immunosorbent Assay (ELISA) methodologies are routinely used in all laboratories of the Middle Eastern countries to detect cereal viruses (Aboul-Ata et al., 1992).
| Table 2: | Characterization of BYDV-PAV sub-serotypes PAVcpA and PAVcpB from different hosts |
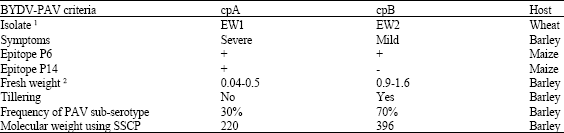 | |
| 1EW1: Egypt wheat isolate 1; EW2: Egypt wheat isolate 2, 2Not infected control, fresh weight 1.7 | |
| Table 3: | Virus-specific primers used in the multiplex RT-PCR detection method (Deb and Anderson, 2008) |
 | |
In this way the five-BYDV isolates (PAV, MAV, RPV, RMV and SGV) from Egypt have been serotyped in big field-collected cereal samples as well as joining weeds (Lister et al., 1994). Tissue blots have been used as targets for BYDV immunodetection during seasonal field surveys in Egypt (Makkouk and Comeau, 1994). Usually, polyclonal antibodies are produced in rabbits injected with purified virus particles. Monoclonal antibodies have been used to discriminate between BYDV-PAV serotypes (Mastarie et al., 1998). Different epitopes could differentiate between BYDV-PAV variants (Table 2). TAS-ELISA was used with monoclonal antibodies specific to CYDV-RPV (Belkahla and Lapierre; 1999; Mastarie et al., 1998). This technique was not only used to detect different BYDV-PAV variants (CpA and CpB), but also BYDV-MAV (Miller et al., 2002). Monoclonal antibodies were used to detect infection by BYDV and CYDV isolates (Belkahla and Lapierre, 1999; Mastarie and Lapierre, 1999).
In the frame of the US AID CDR-funded Middle East Research and Cooperation Program (MERC), Egypt, Israel and Jordan are using synthetic genes encoding the CP of the major cereal viruses for mass production of CP as source of antigens for antibody production (Table 3 and Fig. 3). The viral CP genes are over-expressed in E. coli under an inducible promoter and the protein is purified by affinity chromatography (Akad et al., 2004). Expressed CP constitutes a large source of antigens. The CP epitopes can be manipulated by modifying the sequence of the CP gene. Moreover, antibody specificity may be modulated by changing the sequence of the CP gene in order to increase the ability to detect a given virus strain. CP-produced polyclonal antibodies will assist other Middle East countries for cereal virus diagnosis and detection in plant quarantine and extension service.
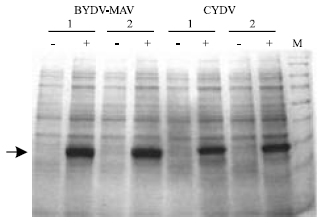 | |
| Fig. 3: | Over-expression of the coat protein of Barley yellow dwarf virus (BYDV-MAV) and Cereal yellow dwarf virus (CYDV) in E. coli. The sequences of the CPs were retrieved from Genbank. The genes were synthesized according to the bacterial codon usage. The CP genes were cloned in the pCAL vector. Two colonies (1, 2) were grown without (-) and with (+) induction with IPTG. The arrow shows the CP |
Detection and Discrimination Using Single and Multiplex PCR
DNA probes have been used to detect BYDV (Habili et al., 1987). Today PCR and reverse-transcription PCR (RT-PCR) are routine for plant virus detection (Henson and French, 1993; Thomson and Dietzgen, 1995; Webster et al., 2004). Several viruses infected a single plant can be detected by multiplex PCR (Gambino and Gribaudo, 2006). A multiplex PCR was designed for BYDV and CYDV detection (Deb and Anderson, 2008). The PCR is one of the most accurate detection means of cereals viruses, provided the sequence is known in order to design specific primers (Robertson et al., 1991). Using the primers described in Table 3 (Deb and Anderson, 2008), singleplex and multiplex PCR allowed to detect BYDV-MAV, BYDV-PAV and BYDV-SGV, but not BYDV-RMV, on wheat plants in Jordan (Fig. 4A). The sequence of BYDV-MAV from Jordan was 91% homologous to BYDV-MAV from the USA (Ueng et al., 1992). On the other hand, the sequence of BYDV-PAV from Jordan was almost identical (98 to 100% homology) to the virus characterized from the USA (Accession No EF521841.1, EF521845; EF521836) but different (78% homology only) from the isolates from Iran (Accession No. FJ687402.1, FJ687396.1; FJ687398.1). Another cereal virus, MDMV was detected by PCR in Jordan and in Egypt on maize plants (Fig. 4B). BYDV-PAV was detected in Egypt by PCR (Fig. 4C).
Virus Typing by RFLP and SSCP
In Egypt, wheat genotypes infected with BYDV-PAV show different degrees of severity. This biological diversity was paralleled with polymorphism in the genome sequence, which can be revealed by restriction fragment length polymorphism (RFLP) and by single strand confirmation polymorphism (SSCP). RT-PCR-RFLP was used to discriminate between BYDV species (Kundu et al., 2009). RFLP allowed to fingerprint CpA and CpB BYDV-PAV variants (Fig. 5A, B). SSCP was used for BYDV-PAV typing (Mastarie et al., 1998). Samples could be infected with PAVcpA, with PAVcpB and some with the two variants (Fig. 5C).
Nucleic Acid Based Phylogeny of Cereal Viruses
The CP of two distinct groups of BYDV-PAV (PAVcpA and PAVcpB) has been sequenced (Mastarie et al., 1998; Miller et al., 2002). Two severe (lethal) and two mild cpA isolates were compared with those of several known PAV cpA isolates to assess variability and locate potential determinants of severity.
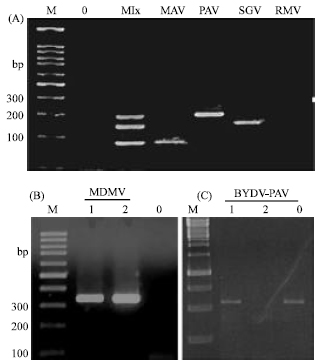 | |
| Fig. 4: | PCR-detection-based detection of cereal viruses in Jordan and Egypt. (A): Singleplex detection of BYDV-MAV (MAV), BYDV-PAV (PAV) and BYDV-SGV (SGV) and multiplex (Mlx) detection of the three viruses in Jordan. (B): detection of MDMV in Jordan. (C): detection of BYDV-PAV in Egypt. The primers used are shown in Table 4. M: molecular weight marker, 1 kbp ladder. 0: non-infected plant |
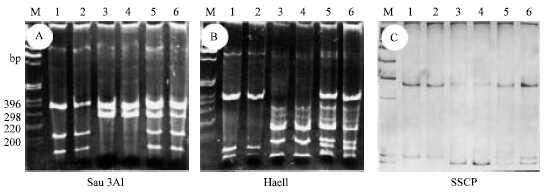 | |
| Fig. 5: | Discrimination between BYDV-PAV sub-serotypes using RFLP (A,B) and SSCP (C). Lanes 1 and 2: samples with cpB type; lanes 3 and 4: samples with cpA type; lanes 5 and 6: samples with both cpA and cpB types. M: molecular weight marker, 1 kbp ladder (Mastarie et al., 1998) |
The isolates FHv1 and FHv2 were different from the mild isolates by eight amino acid substitutions. Phylogenetic tree analyses indicated that, in contrast to the mild isolates, the field-lethal isolates (FHv1 and FHv2) fell into the same cluster, regardless of the genomic region analyzed (Mastarie et al., 1998; Miller et al., 2002; Brown et al., 1996; Boonham et al., 2007). BYDV-PAVs were found all over the world.
BYDV-PAV cpA was diagnosed in Australia, Japan France Egypt, Morocco and USA. BYDV-PAV cpB was detecetd in Egypt, France and Morocco. Mixed infections of BYDV-PAV cpA and cpB were found in Egypt and France (Mastarie et al., 1998; Miller et al., 2002).
Virus Detection Using Microarrays
DNA microarrays, which were introduced to measure the levels of expression of multiple genes in a high throughput mode, have become a major tool for the detection of human as well as of animal and plant pathogens (Uttamchandami et al., 2007). Microarrays have been successfully applied for the detection of HIV, influenza and SARS, respectively (Wang et al., 2002; Sengupta et al., 2003; Long et al., 2004). In the last few years this technology has been applied for the detection of pathogens of economically important agricultural crops (Boonham et al., 2007). Microarrays have been successfully used to detect a number of potato viruses, cucurbit-infecting Tobamoviruses, the potato wart-causing quarantine pathogen Synchitrium endobioticum, Plum pox virus isolates and Fusarium species (Pasquini et al., 2008).
For the microarray-based detection of pathogens, the standard method used today involves the design of a series of 70-mer oligonucleotides (or oligos) with a sequenced derived from that of the pathogen to be detected. For each pathogen, 3 to 5 oligos covering the entire genome are sufficient for adequate detection. The oligos are amino-labelled at their 3` end in order to ensure stable binding to the glass slide on which they are printed using a microarray printer. A single glass slide may contain more than 10,000 oligos (termed the probes). Hence, a single array may contain probes representing hundreds of pathogens. Usually the pathogen is identified by hybridization of fluorescently (usually Cy3) labelled DNA derived from RNA of infected plants that has been reverse-transcribed (or DNA when the pathogen as a DNA genome) and amplified by PCR using random primers. Shorter oligonucleotides have also been used (Wei et al., 2009).
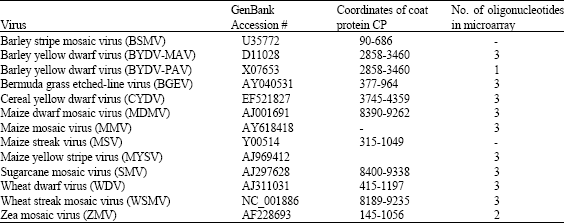
In the frame of our Egypt-Israel-Jordan MERC program, we have designed a 70-mer oligonucleotide microarray based on the sequence of the major cereal viruses present in the Middle East (Table 4). Between 2 and 3 oligonucleotides were designed to specifically represent each virus. A microarray has been manufactured and is presently tested.
| Table 4: | Cereal viruses used to design the diagnostic oligonucleotide microarray and to over-express the coat protein in E. coli |
 | |
CONCLUSIONS
In the Middle East area, diagnosis and typing need coordinated efforts to promote a regional effort to curb the deleterious effects of viruses affecting cereal production. Several technologies are in hand, each with its advantages and disadvantages. Some are based on serology, others on nucleic acid sequences. The expression of the viral coat protein in prokaryotic (E. coli) or eukaryotic (yeast) systems, using synthetic genes can provide the means to obtain unlimited amounts of antigen but also allows fine-tuning the epitope by modeling in silico the CP gene and the expressed protein to fit the particularities of the local viral pathogens. PCR, RT-PCR and RFLP-PCR may provide easy ways to diagnose a given virus and even one of its strains. Microarrays offer a platform where all important viruses (up to hundreds species and strains) can be represented and amenable to diagnosis in a single operation. This technology has been proven to be effective for the diagnosis of human pathogens (Sengupta et al., 2003) as well as plant pathogens (Boonham et al., 2007) and it is thought to be applicable for counteracting agricultural bioterrorism (Uttamchandani et al., 2007). The technology is still expensive compared to ELISA for example (with the high cost of microarray printer and scanner). However, the method allows discriminating all pathogens of a given crop in a single experiment, while many ELISA tests and many different antibodies will need to be tested one by one to identify the culprit pathogens. Prices are expected to decrease with the popularization of the technology.
In any case, virus diagnostic means need to be coordinated and standardize to comply with the highest quality control. This could be done only if protocols are shared, tested and implemented by the various laboratories and plant protection services. The tests should be agreed by the national ministries of agriculture and specialized laboratories should be licensed to perform the tests provided regular inspection. Once protocols are established and agreed upon, surveys of infected plants and carrying vectors could be performed on a regular basis according to the epidemiology of each virus and variant. The cooperation of the growers is essential in such an endeavour. A close cooperation between the grower and the laboratory is a must, including educational presentations, instructions and publications of leaflets. Extreme awareness of the grower will allow a rapid passage of information from the field to the laboratory in order to take rapid countermeasures on a local, regional, national and international basis. A public database should be the repository of any relevant information, from symptoms, epidemiology, diagnosis, to world-wide database. The web should help exchange of communication and information.
Besides efforts towards more precise, rapid and cheap detection means, breeding and release of virus-resistant material is a must and efforts should be encouraged by decision makers at the highest governmental levels, by public funding and by private companies. Plant-extension and protection service can assist big and small-farming farmers in technology transfer. A regional effort that is aimed at providing answers to the above concerns comprises researchers from Egypt, Israel and Jordan. The project, entitled Monitoring of cereal virus and virus-like diseases for prevention through regional detection and quarantine systems is funded by the US Agency for International Development, Middle East Research and Cooperation (MERC) program.
ACKNOWLEDGMENT
This study is supported by a grant from the US Agency for International Development, Middle East Research and Cooperation (MERC) program. Project M21-037 Monitoring of cereal virus and virus-like diseases for prevention through regional detection and quarantine systems.
REFERENCES
- Akad, F., N. Dotan and H. Czosnek, 2004. Trapping of Tomato yellow leaf curl virus (TYLCV) and other plant viruses with a GroEL homologue from the whitefly Bemisia tabaci. Arch. Virol., 149: 1481-1497.
CrossRefDirect Link - Ammar, E.D., S. Elnagar, A.E. Abul-Ata and G.H. Sewify, 1989. Vector and host-plant relationships of the leafhopper-borne maize yellow stripe virus. J. Phytopathol., 126: 246-252.
CrossRefDirect Link - Andret-Link, P. and M. Fuchs, 2005. Transmission specificity of plant viruses by vectors. J. Plant Pathol., 87: 153-165.
Direct Link - Boonham, N., J. Tomlinson and R. Mumford, 2007. Microarrays for rapid identification of plant viruses. Ann. Rev. Phytopathol., 45: 307-328.
PubMed - Brown, C.M., S.P. Dinesh-Kumar and W.A. Miller, 1996. Local and distant sequences are required for efficient read-through of the barley yellow dwarf virus-PAV coat protein gene stop codon. J. Virol., 70: 5884-5892.
Direct Link - D'Arcy, C.J., L.L. Domier and M.A. Mayo, 2000. Family Luteoviridae. In: Virus Taxonomy: Seventh Report of the International Committee on the Taxonomy of Viruses, Van Regenmortel, M.H.V., C.M. Fauquest, D.H.L. Bishop, E.B. Carstens and M.K. Estes et al. (Eds.). Academic Press, San Diego, pp: 775-784.
- Deb, M. and J.M. Anderson, 2008. Development of a multiplexed PCR detection method for Barley and Cereal yellow dwarf viruses, Wheat spindle streak virus, Wheat streak mosaic virus and Soil-borne wheat mosaic virus. J. Virol. Methods, 148: 17-24.
PubMed - Gambino, G. and I. Gribaudo, 2006. Simultaneous detection of nine grapevine viruses by multiplex reverse transcription polymerase chain reaction with coamplification of a plant RNA as internatal control. Phytopathology, 96: 1223-1229.
PubMed - Habili, N., J.L. Mcinnes and R.H. Symons, 1987. Nonradioactive photobiotin-labelled DNA probes for the routine diagnosis of barley yellow dwarf virus. J. Virol. Methods, 16: 225-237.
CrossRef - Henson, J.M. and R. French, 1993. The polymerase chain reaction and plant disease diagnosis. Ann. Rev. Plant Pathol., 31: 81-109.
Direct Link - Jones, R.A.C., 2009. Plant virus emergence and evolution: Origins, new encounter scenarios, factors driving emergence, effects of changing world conditions and prospects for control. Virus Res., 141: 113-130.
PubMed - Kundu, J.K., J. Jarosova, S. Gadiou and G. Cervena, 2009. Discrimination of three BYDV species by one-step RT-PCR-RFLP and sequence based methods in cereal plants from the czech republic. Cereal Res. Commun., 37: 541-550.
CrossRefDirect Link - Li, C., D. Cox-Foster, S.M. Gray and F. Gildow, 2001. Vector specificity of Barley Yellow Dwarf Virus (BYDV) transmission: Identification of potential cellular receptors binding BYDV-MAV in the aphid, Sitobion avenae. Virology, 286: 125-133.
CrossRef - Long, W.H., H.S. Xiao, X.M. Gu, Q.H. Zhang, H.J. Yang, G.P. Zhao and J.H. Liu, 2004. A universal microarray for detection of SARS coronavirus. J. Virol. Methods, 121: 57-63.
PubMed - Makkouk, K.M. and A. Comeau, 1994. Evaluation of various methods for the detection of barley yellow dwarf virus by the tissue-blot immunoassay and its use for virus detection in cereal inoculated at different growth stages. Eur. J. Plant Pathol., 100: 71-80.
CrossRefDirect Link - Mastarie, J. and H. Lapierre, 1999. Sequences of 3'half of the genomes of Barley Yellow Dwarf Virus-PAV cpA isolate that vary in symptom severity. Eur. J. Plant Pathol., 105: 801-811.
Direct Link - Mastarie, J., H. Lapierre and J.T. Dessense, 1998. Asymmetrical distribution of Barley Yellow Dwarf Virus PAV variants between host plant species. Phytopathology, 88: 818-821.
PubMed - Miller, A.W., S. Liu and R. Beckett, 2002. Barley Yellow Dwarf Virus: Luteoviridae or Tombusviridae? Mol. Plant Pathol., 3: 177-183.
Direct Link - Pasquini, G., M. Barba, A. Hadidi, F. Faggioli and R. Negri et al., 2008. Microarray-based detection and genotyping of Plum pox virus. J. Virol. Methods, 147: 118-126.
PubMed - Robertson, N.L., R. French and S.M. Gray, 1991. Use of group specific primers and polymerase chain reaction for the detection and identification of luteoviruses. J. Gen. Virol., 72: 1473-1477.
CrossRef - Sengupta, S., K. Onodera, A. Lai and U. Melcher, 2003. Molecular detection and identification of influenza viruses by oligonucleotide microarray hybridization. J. Clin. Microbiol., 41: 4542-4550.
CrossRefDirect Link - Szunics, L., E. Pocsai, L. Szunics and G. Vida, 2000. Viral diseases on cereals in central hungary. Acta Agronomica Hungarica, 48: 237-250.
CrossRef - Thomson, D. and R.G. Dietzgen, 1995. Detection of DNA and RNA plant viruses by PCR and RT-PCR using a rapid virus release protocol without tissue homogenization. J. Virol. Methods, 54: 85-95.
PubMed - Ueng, P.P., J.R. Vincent, E.E. Kawata, C.H. Lei, R.M. Lister and B.A. Larkins, 1992. Nucleotide sequence analysis of the genomes of the MAV-PS1 and P-PAV isolates of barley yellow dwarf virus. J. General Virol., 73: 487-492.
CrossRefDirect Link - Uttamchandani, M., J.L. Neo, B.N. Ong and S. Moochhala, 2009. Applications of microarrays in pathogen detection and biodefence. Trends Biotechnol., 27: 53-61.
CrossRef - Webster, C.G., S.J. Wylie and M.G.K. Jones, 2004. Diagnosis of plant viral pathogens. Curr. Sci., 86: 1604-1607.
Direct Link - Wei, T., M.N. Pearson, D. Blohm, M. Npolte and K. Armstrong, 2009. Development of a short oligonucleotide microarray for the detection and identification of multiple potyviruses. J. Virol. Methods, 162: 109-118.
CrossRef