
M.A. AL-Saleh
Department of Plant Protection, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia
I.M. AL-Shahwan
Department of Plant Protection, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia
M.A. Amer
Department of Plant Protection, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia
O.A. Abdalla
Department of Plant Protection, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia
International Journal of Virology
Year: 2009 | Volume: 5 | Issue: 3 | Page No.: 131-142







ABSTRACT
This study aimed at identifying the causal agent inducing virus-like symptoms on radish and lettuce plants in Riyadh Region, Saudi Arabia. Mottling, chlorosis and mosaic symptoms were observed on lettuce (Lactuca sativa Linn.) and radish (Raphanus sativus Linn.) plants in two areas in Riyadh region. Mosaic symptoms were observed on L. sativa, R. sativus, Eruca sativa Mill and Brassica rapa Linn., whereas local lesions were observed on Chenopodium ammaranticolor Coste and Reyn., which were mechanically inoculated with sap from infected radish and lettuce plants. Electron microscopy revealed filamentous flexuous particles typical of potyviruses. The aphid Brevicoryne brassicae transmitted the virus to lettuce and radish in a non-persistent manner. Turnip mosaic virus (TuMV) was the only detected virus by double antibody sandwich-enzyme linked immunosorbent assay among five viruses suspected of such disease symptoms. Amplification of total RNA extracted from infected lettuce and radish plants yielded 1-1.2 kbp using three degenerate primers which were designed for potyvirus group detection and 985 bp complementary DNA (cDNA) fragments using a specific oligonucleotide primer for TuMV detection. Nucleic acid sequence analysis of TuMV-L-SA revealed a range of 85.3 to 90.9% sequence identity with other TuMV isolates obtained from the GenBank. However, the TuMV-Ra-SA revealed a range of 87.7 to 94.1% sequence identity. The sequence similarity between TuMV-L-SA and TuMV-Ra-SA investigated in the current study was 89%. To our knowledge, this is the first report of TuMV on lettuce and radish in Saudi Arabia.
PDF Abstract XML References Citation
How to cite this article
M.A. AL-Saleh, I.M. AL-Shahwan, M.A. Amer and O.A. Abdalla, 2009. Etiology of a Mosaic Disease of Radish and Lettuce and Sequencing of the Coat Protein Gene of the Causal Agent in Saudi Arabia. International Journal of Virology, 5: 131-142.
DOI: 10.3923/ijv.2009.131.142
URL: https://scialert.net/abstract/?doi=ijv.2009.131.142
DOI: 10.3923/ijv.2009.131.142
URL: https://scialert.net/abstract/?doi=ijv.2009.131.142
INTRODUCTION
Turnip mosaic virus (TuMV) is an RNA virus which belongs to the genus Potyvirus, within the potyviridae family of plant viruses (Barnett et al., 1995; Provvidenti et al., 1996). All potyviruses have flexuous filamentous particles 700-750 nm long, each of which contains a single copy of the genome, which is a single-stranded positive sense Ribonucleic acid (RNA) molecule. The size of potyviruses RNA is about 10000 nucleotides long. The genomes of potyviruses have a single open reading frame that is translated into a single large polyprotein, which is hydrolysed, after translation, into several proteins by virus-encoded proteinases (Riechmann et al., 1992). Turnip mosaic virus is a widely spread and economically important virus (Walsh and Jenner, 2002). It has been ranked among the five most damaging vegetable viruses worldwide (Tomlinson, 1987) and the only potyvirus known to infect brassicas (Walsh and Jenner, 2002). It was first reported in crucifers by Gardner and Kendrick (1921) and Schultz (1921) in the United States of America. The virus is geographically widespread, of cosmopolitan distribution and has been reported in North America, Europe, Africa, Asia, Australia and New Zealand (Tomlinson and Ward, 1978; Rybicki and Hughes, 1990; Walsh and Tomlinson, 1985; Edwardson and Christie, 1991; Powell and Lindquist, 1992). Turnip mosaic virus has a wide host range and infects over 318 plant species in 156 genera of 43 plant families including many important crops such as cruciferae, compositae, chenopodiaceae, leguminosae and caryophyllaceae, weeds and is also known to infect monocots (Chen et al., 2003; Edwardson and Christie, 1991; Green and Deng, 1985). Turnip mosaic virus infects many crops, including cabbage and other brassica vegetables, oilseed rape, chicory, horseradish, lettuce, peas and rhubarb (Shattuck, 1992). TuMV, like other potyviruses, is transmitted by aphids in the non-persistent manner (Shukla et al., 1994). Eighty nine aphid species were reported to transmit TuMV in a non-persistent manner including Myzus persicae and Brevicoryne brassicae (Walsh and Jenner, 2002). There are potyvirus isolates that appear serologically similar to TuMV when tested with polyclonal antisera that do not readily infect brassicas (Lesemann and Vetten, 1985). Recently, TuMV which has been reported to infect garden rocket in Saudi Arabia, has been described by Al-Saleh et al. (2008). Subsequent to that investigation, viral symptoms were observed on naturally growing lettuce and radish plants in Riyadh region. That fact encouraged carrying out of the present study with the aim of isolation, identification and molecular characterization of the causal agent (s) infecting these crops in the central region of Saudi Arabia.
MATERIALS AND METHODS
Source and Host Range of the Virus Isolates
Samples of lettuce and radish plants growing naturally at Al-Hair and Al-Oyayna areas in Riyadh region, Saudi Arabia and showing mottling, mosaic and chlorosis symptoms (Fig. 1) were collected during the spring of 2007. Seeds of 14 plant species were germinated in small pots filled with a soil mix of sand and peatmoss in the ratio of 2:1. The plant species were: Raphanus sativus L., Eruca sativa Mill, Brassica rapa L., B. oleracea L., Lactuca sativa Linn, Spinacea oleracea, Lycopersicum esculentum L., Datura stramonium L., Chenopodium ammaranticolor Cost and Reyn, Solanum nigrum L., Gomphrena globosa, Nicotiana tabaccum L. and N. glutinosa L. The seedlings were transplanted in large pots filled with the same soil mixture.
 | |
| Fig. 1: | Field symptoms of Turnip mosaic virus (TuMV) on (a) radish and (b) lettuce plants |
Three seedlings of each of the plant species were transplanted in each of six pots. Inoculum was prepared by separately grinding leaves of R. sativus and Lactuca sativa showing mosaic symptoms, in 0.01 M potassium phosphate buffer (Winlab, United Kingdom), pH 7.0, in a ratio of 1:4 and then filtered through cheese cloth. Two sets of seedlings (two pots per each plant species) of the above plants were separately inoculated with the prepared inoculum from radish and lettuce after being dusted with carborundum (300 mesh)( Fisher Scientific, USA). One pot containing three seedling of each plant species was inoculated with sap from healthy radish or lettuce prepared in the same way as control. The seedlings were then rinsed with distilled water and kept in the greenhouse (Chen et al., 2003). These experiments were repeated twice.
Electron Microscopy
The leaf dip method described by Hill (1984) was adopted. Leaf pieces of infected lettuce and radish plants were sliced at the edges and immersed several times in a drop of potassium phosphate buffer, pH 7, on carbon coated copper grids. Excess buffer was removed using Whatman filter papers (Whatman international Ltd., England). A drop of 1% phosphotungstic acid (BDH Chemicals Ltd., England) was placed on each grid for 45 sec and was then removed using Whatman filter papers. The grids were then observed in the electron microscope for virus detection.
Aphid Transmission
Brevicoryne brassicae L. were collected from uninfected radish plants in Oyaynah area and left starving for an hour in Petri dishes size 94/16 (bottger, Germany). The aphids in some of these plates were then allowed to feed briefly on pieces of healthy radish leaves subsequent to the starvation period, while others were allowed to feed for 3 min on pieces of radish leaves infected with TuMV and showing mosaic symptoms. After the insects in both groups were fed for 3 min, they were quickly transferred singly by a brush onto healthy radish seedlings (5 insects/plant. 10 plants/treatment). The aphids in both groups were allowed to feed on these seedlings for 2 min after their transfer. The seedlings were then sprayed with an insecticide (cyprine 100 EC) to kill the insects. Seedlings were kept in the greenhouse and observations regarding symptoms expression were recorded. Lettuce plants were tested in the same manner (Shattuck, 1992).
Serological Detection of the Virus
DAS-ELISA described by Clark and Adams (1977) was used for the detection of the virus that induced symptoms in lettuce and radish. ELISA kits of Alfalfa Mosaic Virus (AMV), Cucumber Mosaic Virus (CMV) and Tomato Spotted Wilt Virus (TSWV), Lettuce Mosaic Virus (LMV) and Turnip Mosaic Virus (TuMV) were purchased from Agdia Inc., (Indiana, USA). Steps of ELISA procedure were followed as described on the label by Agdia, Inc. Each of four micro titer plates was coated with antibodies of each virus after being diluted with the coating buffer. Subsequent to incubation and washing, aliquots of 100 μL of each of the samples which were extracted in the extraction buffer, were added in two wells of each plate. One hundred microliters of the proper dilutions of the relevant antibody-alkaline phosphatase conjugate were dispensed in the wells of each plate subsequent to washing plates from samples sap. P-nitrophenyl phosphate solution was then added in the wells of each plate after washing from the conjugate solution. The plates were incubated for 1 h and the reaction was then stopped using 3 M NaOH. The plates were read at 405 nm in the plate reader (minireader II, Karl Kolb, Germany).
RNA Extraction and RT-PCR
The SV-Total RNA Isolation System (Promega, USA) was used for total RNA extraction from infected and uninfected lettuce and radish plants. The QIAGEN One Step RT-PCR Kit (QIAGEN, USA) was used to perform the RT-PCR according to the manufacturer’s instructions. The oligonucleotide primers used for Potyvirus degenerate primer were designed according to Hsu et al. (2005) as follow: PNIbF1 5`-ggB aaY aaT agt ggN caa cc-`3, PNIbF5 5`-gcc agc cct cca ccg tNg tNg aYa-`3 and PCPR1 5`- GGG GAG GTG CCG TTC TCD ATR CAC CA -`3 (where B = C, G and T, Y = C and T, N = A, C, G and T, D = A, G and T and R = A and G). The oligonucleotide primers used for TuMV detection were designed according to Sanchez et al. (2003) as follows: the upstream primer Tu 8705- 8726: 5`-CAA GCA ATC TTT GAG GAT TAT G-`3 and the downstream primer Tu 9690-9669 : 5`-TAT TTC CCA TAA GCG AGA ATA C-3` were used. RT-PCR was performed according to QIAGEN manufacturer’s (QIAGEN, USA) recommendations. Briefly, 10 μL of 5xQIAGEN One Step RT-PCR Buffer, 2 μL of 10 mM dNTP Mix, 10 μL of 5x Q-Solution, 2 μL of 10 pmol of each complementary and homologous primers, 2 μL of QIAGEN One Step RT-PCR Enzyme Mix, 5-10 units/reaction of RNase inhibitor, A total of 5 μL (200 ng) of RNA was added to the one-step and RNase-free water to 50 μL. The master mix was mixed gently, by pipetting up and down a few times. RT-PCR reaction mixture was amplified using the following cycling parameters: hold at 50°C for 30 min (RT step), hold at 95°C for 15 min (hot start to PCR), then subjected to 35 cycles of amplification: 94°C for 30 sec, 55°C for 45 sec and 72°C for 1 min and a final incubation at 72°C for 7 min for potyvirus detection, but hold at 50°C for 30 min (RT step), hold at 95°C for 15 min (hot start to PCR), then subjected to 35 cycles of amplification, 30 sec at 94°C for denaturation, 30 sec at 54°C for annealing and 60 sec at 72°C for extension, followed by a final hold at 72°C for 10 min for TuMV detection. For electrophoresis analysis, aliquots 10 μLeach of PCR amplified DNA (Promega, USA) products were mixed with gel loading buffer. Separation was done on a 1% agarose gel in 1XTBE buffer (1x = 89 mM Tris, 89 mM borate and 2 mM EDTA, pH 8.3). DNA was stained with ethidium bromide added to the gel at a concentration of 0.5 μg mL-1. DNA was visualized on a UV transilluminator and photographed using DNA documentation gel analysis (model No. OptiGo 500, Isogen, Holland). DNA 1 kb marker (Promega, country) was used to determine the size of RT-PCR amplified cDNA products (Sambrook et al., 1989).
Sequencing of the Coat Protein Gene of Tumv and Data Analysis
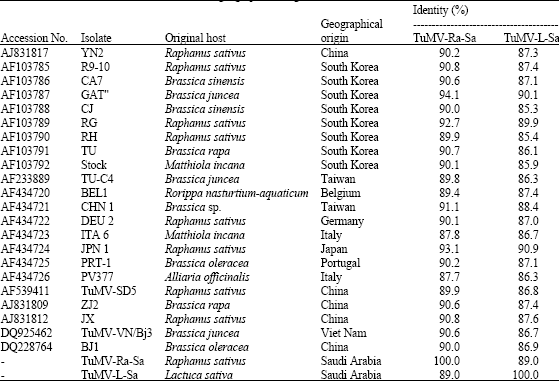
The nucleotide sequence of the amplified DNA fragment of expected size (985 bp) of the coat protein gene which included 54 bp of the 3`-end of NIb gene and 65 bp of the 3`-UTR was determined. The amplified PCR product was purified using the Wizard PCR clean up kit (Promega, USA). The detrermination of the nucleotide sequence of the isolated gene of TuMV was carried out in one direction with the specific complementary primer at King Faisal Specialist Hospital and Research Center, Biological and Medical Research Department, Riyadh, Kingdom of Saudi Arabia using AB 3730 xl DNA Analyzer (Applied Biosystem-HITACHI, Japan). Sequence analysis were performed in triplicates for each sample and the homolog tree analysis were done using DNAMAN trial version 5.2.10 program (Lynnon BioSoft, Quebec, Canada). The GenBank accession numbers for the different isolates of TuMV shown in Table 1 were used for comparison.
| Table 1: | Homology matrix between the Saudi Arabian isolates of TuMV (lettuce TuMV-L-Sa and radish TuMV-Ra-Sa) and 22 isolates of different host and geographical origins. |
 | |
RESULTS
In the repeated host range experiments, mosaic symptoms were observed on the inoculated radish, lettuce, garden rocket and turnip (Fig. 2a-d) which were mechanically inoculated with sap from infected plants whereas local lesions were observed on C. amaranticolor (Fig. 2e). No symptoms were observed on the rest of the inoculated plants. Also, no symptoms were observed on the control plants. TuMV was transmitted to both radish and lettuce by B. brassicae L., in a non persistent manner. Eight out of the 10 radish seedlings and 7 out of the 10 lettuce seedlings on which aphids fed on the infected plants were allowed to feed, exhibited mosaic symptoms. No symptoms were observed on the control plants on which aphids fed on healthy plants only, were allowed to feed. Although, several aphid vectors for TuMV were reported to occur in Saudi Arabia (Aldryhim and Khalil, 1996) where some such as M. persicae were more efficient than B. brassicae, the later was tested because it was the only species encountered prior to the insect transmission experimentation. The transmission electron microscope indicated presence of filamentous, flexible, virus particles (Fig. 2f) typical of potyviruses on the negatively stained, carbon coated grids that were prepared from infected leaf pieces of lettuce and radish using the leaf dip method. The twenty lettuce samples and the thirty radish samples that were collected from field grown plants in Riyadh governorate (El-Hayer and Oyayna fields) were showing conspicuous mosaic symptoms of light and dark green areas in the leaves. ELISA test of these samples was positive with TuMV antiserum only and negative with antisera to AMV, CMV, TSWV and LMV that commonly infect lettuce and radish. Turnip mosaic virus as a potyvirus infecting lettuce and radish plants was detected separately by RT-PCR using PCPR1/PNIbF1 and PCPR1/PNIbF5 PCR primers. The expected band on the agarose gel after RT-PCR amplification was about 1.2 and 1 kb, respectively. RT-PCR was, therefore, utilized successfully in this study to detect a potyvirus in the infected lettuce and radish plant tissues. Electrophoresis analysis of RT-PCR product showed single amplified fragments of 1.2 and 1 kb (Fig. 3), (lane 1, 3, 5, 6) respectively. No amplified fragments of cDNA were obtained from uninfected lettuce leaves (lane 2, 4).
 | |
| Fig. 2: | Mosaic symptoms of Turnip mosaic virus (TuMV) on inoculated (a) radish, (b) lettuce, (c) garden rocket, (d) turnip (d) plants and (e) local lesions on Chenopodium amaranticolor. An electron micrograph showing filamentous and (f) flexible virus particles of TuMV |
To confirm these results, RT-PCR was performed on total RNA extracted from 30 mg infected and uninfected lettuce and radish plants with TuMV isolate using the SV total RNA Isolation System Kit (Promega, USA). The RNA was reverse transcripted using reverse transcriptase. The reverse transcription reaction was primed with the complementary primer specific for TuMV. The resulting cDNA was amplified by PCR after adding the complementary and homologous primers. RT-PCR amplification of viral RNA was carried out on the total RNA isolated from infected and uninfected lettuce and radish plants using specific primer for TuMV-CP Sanchez et al. (2003) designed to amplify 985 bp of the coat protein gene. Electrophoresis analysis of RT-PCR product showed a single amplified fragment of 985 bp. Figure 4 shows the agarose gel electrophoresis of RT-PCR amplified TuMV-CP cDNA from infected lettuce and radish (lane 1, 2), respectively. No amplified fragments of cDNA were obtained from healthy lettuce leaves (lane 3).
The PCR product of TuMV-CP gene which was isolated from lettuce and radish and sequenced two times in the research center of King Faisal specialist Hospital, at Riyadh, Saudi Arabia, indicated that their sequences were composed of 878 and 864 nucleotides in length, respectively.
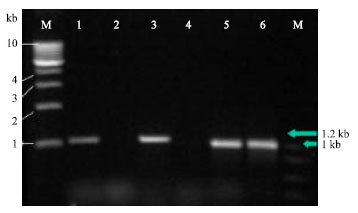 | |
| Fig. 3: | Different potyviruses detected by RT-PCR and potyvirus degenerate primers. Total RNA of potyvirus-infected lettuce and radish plants was extracted and analyzed. The RT primer was PCPR1. The PCR primer pair used to detect potyvirus infection was PCPR1/PNIbF1 from lettuce and radish Lane 1 and 3 and the size of the RT-PCR fragment was about 1.2 kb and PCPR1/PNIbF5 and the size of the RT-PCR fragment was about 1.0 kb from lettuce and radish Lane 5 and 6. Lane M represents 1KB DNA Marker. A healthy tissue control via SV-Total RNA Isolation System kit (lane 2 and 4). RT-PCR products were analyzed in 1% agarose gel |
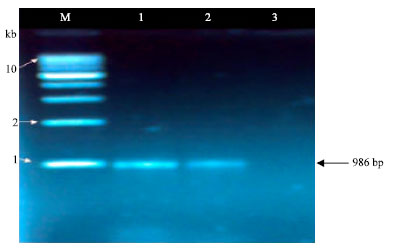 | |
| Fig. 4: | Gel electrophoresis on RT-PCR amplification of a fragment from TuMV genome using specific primer pair designed to amplify 986 bp fragment of CP gene. Lane M represents 1KB DNA Marker. Lanes 1, 2 were loaded from different two isolates infected lettuce and radish with TuMV and processed with the SV-Total RNA isolation system, a healthy tissue control via SV-total RNA isolation system kit (lane 3) |
The CP gene sequence of the TuMV Saudi Arabian isolates of TuMV isolated from lettuce and radish were, hence, unequal in length. The homology percentages and the results of the multiple alignments done along with the sequences previously obtained by GenBank sequence data at the National Center for Biotechnology Information (NCBI) are shown in Table 1 and Fig. 5, respectively.
 | |
| Fig. 5: | The phylogenetic homology tree based on multiple sequence alignments of the TuMV-L-Sa and TuMV-Ra-Sa isolates compared to previously sequenced isolates |
The nucleic acid sequence analysis of the lettuce isolate of turnip mosaic virus designated, TuMV-L-Sa, revealed a range of 85.3 to 90.9% sequence identity with the isolates obtained from the GenBank. The highest sequence similarity was found with TuMV JPN 1 isolate isolated from Raphanus sativus, Japan #AF434724 (90.9%), while the lowest was found with CJ isolate from Chinese cabbage, South Korea No. AF103788 (85.3%). However, sequence analysis of the nucleic acid of the radish isolate, TuMV-Ra-Sa, revealed a range of 87.7 to 94.1% sequence identity with the GenBank isolates. The highest sequence similarity was found with TuMV GAT strain isolated from Brassica juncea, South Korea No. AF103787 (94.1%) whereas the lowest was found with PV377 isolate from Alliaria afficinalis, Italy No. AF434726 (87.7%). A phylogenetic tree illustrating homologous relationships based on multiple alignments of CP nucleotide sequences of 22 TuMV isolates and the Saudi isolates of TuMV (lettuce isolate, TuMV-L-Sa and radish isolate, TuMV-Ra-Sa) is shown in Fig. 5.
DISCUSSION
Turnip mosaic virus detected in vegetables in this study as well as in a earlier study (Al-Saleh et al., 2008) has been reported to cause diseases in vegetables and other crops elsewhere in the world (Zdenka, 1980; Stavolone et al., 1998; Hughes et al., 2002) and is also being recently reported to continue inducing disease (Robertson and Ianson, 2005; Pallett et al., 2008). The virus has many strains which are variable in their pathogenicity, antigenicity and the plant species they infect (Stavolone et al., 1998; Pallett et al., 2008; Zdenka, 1980). This could probably explain the somewhat narrower host range of the TuMV isolate in this investigation which infected relatively a few number of plants, different from the wide host range isolates reported in earlier studies (Chen et al., 2003; Edwardson and Christie, 1991; Green and Deng, 1985). N. glutinosa was also not infected with an isolate of TuMV used in another investigation (Zdenka, 1980). One of the possible reasons for the spread of this virus is the aphid species from which two species were recorded in this study, B. brassicae and M. persicae, in addition to the others reported in earlier studies (Aldryhim and Khalil, 1996) out of the many aphid species reported to transmit this virus.
To develop a similar identification method for the new virus which belongs to the potyvirus group, local conserved regions in the core domain of the potyvirus coat protein and in the NIb replicase protein were selected to provide nucleotide sequences of the construction of degenerate primers for application in a potyvirus group specific combined assay of RT-PCR. The available potyvirus sequence data made it possible to develop a method for the identification of potyviruses based upon the RT-PCR. The sequence between the downstream and upstream primers used in this study represents a highly conserved region of the coat protein gene which should be identical in size in every potyvirus (Langeveld et al., 1991). Three degenerate primers, located at the NIb and CP gene regions, were designed for potyvirus detection. Using these primer pairs, 1.0-1.2 kb cDNA fragments of the 3-terminal region of 6 potyviruses were successfully amplified from infected plant tissues. RT-PCR products were sequenced and found to be derived from the expected viruses (Hsu et al., 2005). An RT-PCR based method, which has the potential to detect members of the genus Potyvirus by using new designed potyvirus degenerate primers, was developed. Since, the amino acid sequence GNNSGQPSTVVDN is highly conserved among potyviruses, the primers PNIbF1 and PNIbF5 that are derived from the 5` and `3 region of this coding sequence should amplify any potyvirus. However, more experiments are needed to confirm this possibility. Although, similar methods have been described previously by Langeveld et al. (1991), Gibbs and Mackenzie (1997) and Chen et al. (2001), the procedure used in this study was different since, two primer pairs were used to avoid false negative results. Accordingly, a potyvirus should be amplified from the RT-PCR method resulting in a product of 1.0-1.2 kb, depending on the size of the `5-terminus of the CP gene and also the `3-terminus of the NIb gene, but no specific amplification was observed with cucumovirus, carmovirus, potexvirus and tobamovirus. Furthermore, using the PCPR1 potyvirus degenerate primer, rather than a dT primer, as the reverse primer for the RT-PCR can avoid the potential problem of interaction with plant poly(A)+ mRNAs. Therefore, this RT-PCR method can be used as a rapid detection method for potyviruses (Hsu et al., 2005).
The CP gene sequences of the Saudi Arabian isolates of TuMV from lettuce and radish were composed of 878 and 864 nucleotides in length, respectively. The length of nucleotide sequence for TuMV isolates reported from different plant species, so far, ranges between 864 and 867. These differences in lengths are probably inherent since they were encountered in isolates infecting the same or different plant species in the same or distant geographical locations (Table 1) (Zhuang et al., 2006; Ha et al., 2008; Shi et al., 2008). However, the length of the nucleotide sequence of the lettuce isolate of TuMV is reported for the first time in this study and accounted for 878 nucleotides.
Based on symptoms expression, serology, molecular detection and the nucleotide sequences of CP genes, the agents that infected lettuce and radish were identified as isolates of TuMV and designated (TuMV-L-Sa and TuMV-Ra-Sa). These findings confirmed the wide spread of this virus in cruciferous crops reported by Edwardson and Christie (1991). It is also reported to be naturally occurring in ornamental crops, lisianthus (Chao et al., 2000) and statice (Powell and Lindquist, 1992). To our knowledge, this is the first record that TuMV infects lettuce plants. TuMV may have been infecting lettuce for many years without being detected. Present results suggest that the ability to infect lettuce is a common characteristic among TuMV strains. This is further supported by our observations that TuMV-infected plants were frequently found in lettuce fields adjacent to turnip or radish. Thus, the virus is disseminated from TuMV-infected cruciferous plants to lettuce and radish plants by aphids. This can happen in any geographical areas where both lettuce and cruciferous crops are grown side by side. TuMV induces severe systemic symptoms on lettuce and radish. Its infection will jeopardize lettuce and radish production.
In this study, we have been able to identify and characterize TuMV from two plant species, which was the main objective of this study. The significance of this investigation stems from the fact that this is the first encounter of this virus on these two crops in Saudi Arabia. Although, the two TuMV isolates from radish and lettuce are from the same region and infected the same host range, they were found not to be closely related to each other (Fig. 5). It is worth to mention that the radish isolate was also found to be more closely related to the GenBank isolates used in this study than the lettuce isolate. These results are also of great significance since, they will help in the management of this disease.
ACKNOWLEDGMENT
We thank the Agriculture Research Center at the College of Food and Agricultural Sciences for the financial support of this research project (designated PLP 41) and the help of Dr. Ahmed Ali Al-Qahtani at the Biological and Medical Research Department, King Faisal Specialist Hospital and Research Center, in providing the sequencing facility, is greatly appreciated.
REFERENCES
- AL-Saleh, M.A, I.M. Al-Shahwan, M.A. Amer and O.A. Abdalla, 2009. Serological and molecular detection of a turnip mosaic virus isolate infecting lettuce in the Kingdom of Saudi Arabia and determination of its coat protein gene nucleotide sequence. Proceedings of the 1st International Conference on Biotechnology.
- Barnett, O.W., G. Adam, A. Brunt, J. Dijkstra and W.G. Dougherty et al., 1995. Family potyviridae. Proceedings of the Sixth Report of the International Committee on Taxonomy of Viruses, 1995, Springer-Verlag, Wien, New York,-pp: 348.
CrossRef - Chen, C.C., C.H. Chao, S.D. Yeh, H.T. Tsai and C.A. Chang, 2003. Identification of turnip mosaic virus isolates causing yellow stripe and spot on calla lily. Plant Dis., 87: 901-905.
CrossRefDirect Link - Chen, J., J. Chen and M.J. Adams, 2001. A universal PCR primer to detect members of the Potyviridae and its use to examine the taxonomic status of several members of the family. Arch. Virol., 146: 757-766.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Gibbs, A. and A. Mackenzie, 1997. A primer pair for amplifying part of the genome of all potyvirids by RT-PCR. J. Virol. Methods, 63: 9-16.
PubMedDirect Link - Green, S.K. and T.C. Deng, 1985. Turnip mosaic virus strains in cruciferous hosts in Taiwan. Plant Dis., 69: 28-31.
Direct Link - Ha, C., P. Revill, R.M. Harding, M. Vu and J.L. Dale, 2008. Identification and sequence analysis of potyviruses infecting crops in Vietnam. Arch. Virol., 153: 45-60.
Direct Link - Hsu, Y.C., T.J. Yeh and Y.C. Change, 2005. A new combination of RT-PCR and reverse dot blot hybridization for rapid detection and identification of potyviruses. J. Virol. Methods, 128: 54-60.
CrossRefDirect Link - Hughes, S.L., S.K. Green, D.J. Lydiate and J.A. Walsh, 2002. Resistance to Turnip mosaic virus in Brassica rapa and B. napus and the analysis of genetic inheritance in selected lines. Plant Pathol., 51: 567-573.
Direct Link - Langeveld, S.A., I.M. Dore, I. Memelink, A.R.L.M. Derks, C.I.M. Van der Vlugt, C.J. Asjes and J.E. Bol, 1991. Identification of potyviruses using polymerase chain reaction with degenerate primers. J. Gen. Virol., 72: 1531-1541.
Direct Link - Pallett, D.W., J.I. Cooper, H. Wang, J. Reeves and Z. Luo et al., 2008. Variation in the pathogenicity of two turnip mosaic virus isolates in wild UK Brassica rapa provenances. Plant Pathol., 57: 401-407.
CrossRefDirect Link - Riechmann, J.L., S. Lain and J.A. Garcia, 1992. Highlights and prospects of potyvirus molecular biology. J. Gen. Virol., 73: 1-16.
Direct Link - Robertson, N.L. and D.C. Ianson, 2005. First report of turnip mosaic virus in Rhubarb in Alaska. Plant Dis., 89: 430-430.
Direct Link - Rybicki, E.P. and F.L. Hughes, 1990. Detection and typing of maize streak virus and other distantly related geminiviruses of grasses by polymerase chain reaction amplification of a conserved viral sequence. J. Gen. Virol., 71: 2519-2526.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sanchez, F., X. Wang, C.E. Jenner, J.A. Walsh and F. Ponz, 2003. Strains of turnip mosaic potyvirus as defined by the molecular analysis of the coat protein gene of the virus. Virus Res., 94: 33-43.
Direct Link - Shi, M., H.Y. Li, J. Schubert and X. Zhou, 2008. Sequence analysis of cp and Hc-pro genes of turnip mosaic virus isolates from china. Acta Virol., 52: 59-62.
Direct Link - Stavolone, L., D. Alioto, A. Ragozzino and J.F. Laliberte, 1998. Variability among Turnip mosaic potyvirus isolates. Phytopathology, 88: 1200-1204.
Direct Link - Tomlinson, J.A., 1987. Epidemiology and control of virus diseases of vegetables. Ann. Applied Biol., 110: 661-681.
CrossRefDirect Link - Tomlinson, J.A. and C.M. Ward, 1978. The reactions of swede (Brassica napus) to infection by turnip mosaic virus. Ann. Applied Biol., 89: 61-69.
CrossRefDirect Link - Walsh, J.A. and C.E. Jenner, 2002. Turnip mosaic virus and the quest for durable resistance. Mol. Plant Pathol., 3: 289-300.
CrossRefDirect Link - Walsh, J.A. and J.A. Tomlinson, 1985. Viruses infecting winter oilseed rape (Brassica napus ssp. oleifera). Ann. Applied Biol., 107: 485-495.
CrossRefDirect Link