
Ashraf M. Metwally
Cairo Poultry Company, Giza, Egypt
Ausama A. Yousif
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Egypt
Iman B. Shaheed
Department of Pathology, Faculty of Veterinary Medicine, Cairo University, Egypt
Walaa A. Mohammed
Department of Viral Vaccines of Poultry, Veterinary Serum and Vaccine Research Institute, Abbasia, Egypt
Attia M. Samy
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Egypt
Ismail M. Reda
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Egypt
International Journal of Virology
Year: 2009 | Volume: 5 | Issue: 1 | Page No.: 1-17







ABSTRACT
Infectious Bursal Disease (IBD) serotype I viruses continue to cause major economic losses in the Egyptian poultry industry despite the implementation of intensive vaccination programs. A recent increase in IBD related mortality in vaccinated farms prompted this investigation into the genetic character of the circulating IBD Virus (IBDV). Bursa and proventriculus samples were RT-PCR tested using novel primers flanking VP2 region coding the two major and two minor hydrophilic peaks. Infectious Bursal Disease virus was detected in tested samples. Phylogenetic analysis of the sequenced PCR product and deduced amino acid sequences of IBDV Giza 2008 VP2 demonstrated the continued circulation of very virulent IBDV (vvIBDV). The mutations reported in Giza 2008 demonstrate that Egyptian field viruses are isolating from their European ancestors. Some of the aa mutations have lead to a change in some of the exposed regions of the viral protein. Present findings explain the continued presence of vvIBDV in intensively vaccinated flocks.
PDF Abstract XML References Citation
How to cite this article
Ashraf M. Metwally, Ausama A. Yousif, Iman B. Shaheed, Walaa A. Mohammed, Attia M. Samy and Ismail M. Reda, 2009. Re-Emergence of Very Virulent IBDV in Egypt. International Journal of Virology, 5: 1-17.
DOI: 10.3923/ijv.2009.1.17
URL: https://scialert.net/abstract/?doi=ijv.2009.1.17
DOI: 10.3923/ijv.2009.1.17
URL: https://scialert.net/abstract/?doi=ijv.2009.1.17
INTRODUCTION
Infectious Bursal Disease (IBD) serotype I viruses continue to cause direct and indirect significant economic losses to the poultry industry. The direct economic impact of IBD is due to the high mortality rates (Chettle et al., 1989; Van den Berg et al., 1991). The indirect economic impact is due to IBDV-induced immunosuppression of infected birds (Allan et al., 1972), which is a leading cause of vaccination failure and bad performance in chicken (Giambrone et al., 1976; Giambrone, 1979).
Immunosuppression following IBDV infection is due to destruction of B-lymphocyte precursors in the bursa of Fabricius (Hirai et al., 1981). Histopathologic lesions occur in the bursa, spleen, thymus, harderian gland and cecal tonsils. The first signs of infection occur in the bursa and it is the most severely affected organ. Degeneration and necrosis of individual lymphocytes in the medullary region of the bursa occur as early as one day post infection (Cheville, 1967).
Infectious Bursal Disease viruses are non-enveloped, icosahedral members of the genus Avibirnavirus of Birnaviridae (Dobos et al., 1979; Hirai and Shimakura, 1974). The double stranded RNA genome of IBDV is composed of 2 segments; A and B codes for five Viral Proteins (VP). The larger segment, A, encodes VP2, VP4 and VP3 in large Open Reading Frame (ORF). In addition, segment A also contain a small ORF partially overlapped the other ORF, which encodes VP5. The smaller segment, B contain one ORF encoding VP1; the RNA-dependent RNA polymerase. The major structural proteins of the virion are VP2 and VP3, both of which are constituents of the IBDV capsid. VP2 carries the major neutralizing epitopes (Azad et al., 1987; Becht et al., 1988). Neutralizing monoclonal antibodies against VP2 can be used to differentiate the serotypes and strains (Becht et al., 1988; Fahey et al., 1989). The VP2 is also responsible for antigenic variation (Brown et al., 1994; McAllister et al., 1995; Snyder et al., 1988; Vakharia et al., 1994a) and virulence (Brown et al., 1994; Yamaguchi et al., 1996).
Two distinct serotypes, I and II, have been identified (Jackwood and Saif, 1987; Jackwood et al., 1985; McFerran et al., 1980). All known pathogenic IBDV strains belong to serotype I. Pathogenic IBDV serotype I isolates are commonly grouped based on antigenic and pathogenic properties in one of 6 categories; mild, intermediate, intermediate plus, classical, variant and very virulent as described by Van den Berg (2000).
Reverse transcription polymerase chain reaction (RT-PCR) using various primers is applied for detection of IBDV (Lee et al., 1992; Wu et al., 1992; Stram et al., 1994). Studies of nucleotides and deduced amino acids sequence changes occurring in segment A have been adopted to differentiate or correlate between IBD viruses either field or vaccine strains (Vakharia et al., 1992; Brown et al., 1994; Qian and kibenge, 1994; Vakharia et al., 1994b; Van den Berg et al., 1996; Yamagushi et al., 1997; Sellers et al., 1999; Yu et al., 2001; Zierenberg et al., 2000).
Several reports have classified the Egyptian IBDV isolates as classical IBDV (Khafagy et al., 1991; El-Sanousi et al., 1994; Bekhit, 1996a, b). On the other hand, some reports have provided partial evidence of the presence of antigenically variant IBDV strains in Egyptian flocks (El-Sanousi et al., 1994; Sultan, 1995). In 2002 direct detection of IBDV antigens in bursal homogenates using monoclonal antibodies against classical and variant epitope markers provided evidence of the presence of antigenically variant IBDV strains in Egyptian flocks (Metwally et al., 2003). A year later variant IBDV was isolated (Hussein et al., 2003).
Infectious Bursal Disease (IBD) serotype 1 viruses continue to cause major problems in the Egyptian poultry industry. The emergence of variant and vvIBDV has caused considerable concern regarding the vaccine control of IBD in spite of extensive and multiple administrations of various live vaccines (Hassan et al., 2002). In 1999, a new Egyptian IBDV strain, designated 99323, was isolated and identified (Eterradossi et al., 2004). The nucleotide sequencing of the variable region of gene encoding VP2 further showed an atypical antigenic profile of strain 99323 related to some critical amino acids changes. The nucleotides sequence of the 99323 isolate was mostly similar with to that of reference European vvIBDV strain 89163 (98.0% nucleotides identity). Abd El-Moaty (2004) identified 2 Egyptian isolates; Kal2001 and Giza2000. Sequence analysis of Kal2001 showed sequence homology with classical IBDV strains ranging between 98.8 and 99.6%. While, Giza2000 showed relatedness to vvIBDV strains with sequence homology ranging between 98.1 and 98.3%.
Re-emergence of variant or highly virulent forms has been the cause of significant economic losses. Vaccination failures were described in different parts of the world. The inception of very virulent IBD created the need for a better characterization of the circulating strains so that, the vaccination schedule could be adapted faster to a new epidemiological situation (Van den Berg, 2000). This study aims at characterization of one of the circulating IBD viruses in broiler flocks receiving classical IBDV vaccines. In addition, this report also describes a novel IBDV primer and its use in the molecular characterization of a central immunogenic region of the viral VP2.
MATERIALS AND METHODS
Sampling and Sample Preparation
Samples were collected from a commercial broiler flock with a slight increase in reported mortalities due to clinical IBD. Gross examination of the dead birds revealed hemorrhages, swelling and exudates in bursa, with bursa/body weight ratios averaging 1.9. Hemorrhages were also noticed on the mucosa of the proventriculus.
| Table 1: | IBDV strains used in sequence analysis and phylogeny |
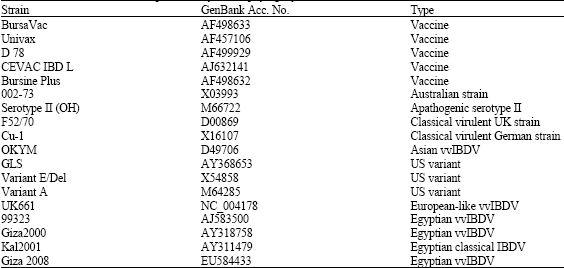 | |
A routine IBDV vaccination program was meticulously implemented before the increase in mortalities. One-day-old broiler chicks were vaccinated using Univax® BD (Shering-Plough, USA) according the manufacturer’s recommendations. At 16 days, the chicks were vaccinated using Bursine® Plus (Fort Dodge, USA). Samples from bursae and proventriculi of 3 4-weeks-old chickens that succumbed to the disease were collected and preserved in formalin for histopathology or at -80°C until used for RNA extraction. Bursa and proventriculus samples were collected from SPF chicks (obtained from the SPF production facility in Fayoum, Egypt), processed and preserved as before. SPF samples served as negative controls in the experiment.
Viruses and Reference Sequences
The vaccinal IBDV strain Bursa-Vac® 3 (Schering-Plough, USA) and virulent SPF-chicken propagated IBDV (Yousif et al., 2006) were used as control viruses in every RT-PCR experiment. GenBank published classical, very virulent, vaccinal and variant sequences were selected for sequence comparisons and phylogenetic analysis (Table 1).
Total RNA Extraction
Samples were prepared for RNA extraction by disrupting one part of each bursa or proventriculus sample in sterile saline (1:1). Bursal homogenates were pooled. Proventriculus samples were also homogenized and pooled as before. A previously tested IBDV-positive bursa from a challenge virus (see above) and SPF tissues were prepared as tested samples. The IBD vaccine included in the experiment was reconstituted in RNAse-free water. RNA was also extracted from bovine sera, ovine sera, plant and bacterial cells for specificity testing of the primers. Total RNA extraction was carried out using RNeasy® Mini kit (QIAGEN, GmbH, Hilden, Germany) according to the manufacturer’s instructions.
Primer Design and Reverse Transcription/Polymerase Chain Reaction (RT/PCR)
Novel primers recognizing conserved regions of the IBDV VP2 flanking the hypervariable region were designed after reviewing published primers and sequences (Bayliss et al., 1990; Heine et al., 1991; To et al., 1999; Spatas and Ignjtovic, 2000; Banda et al., 2001). The primer sequences were as follows; the forward primer [AUS GU: 5’-TCA CCG TCC TCA GCT TAC CCA CAT C-3’] and the reverse primer [AUS GL: 5’-GGA TTT GGG ATC AGC TCG AAG TTG C-3’]. Primers were used for amplification of a 620 bp fragment within IBDV VP2. Oligos were manufactured by Metabion GmbH, (Lena-Christ-Strasse, Germany).
RT-PCR
Briefly, the reaction mixture contained 1x of OneStep RT-PCR Enzyme Mix (containing Omniscript Reverse Transcriptase, Sensiscript Reverse Transcriptase and HotStarTaq DNA Polymerase), 0.2 U μL-1 RNase inhibitor, 400 μM of each of the deoxynucleotide triphosphates and 100 pmol each of primers, in a total volume of 50 μL QIAGEN OneStep RT-PCR Buffer containing 2.5 mM magnesium chloride (MgCl2). The PCR reaction was performed in the thermal cycler (Perkin Elmer 9700) as follows: 20 min at 50°C (RT reaction); 95°C for 15 min (initial PCR activation); 39 three-step cycles of 94°C for 30 sec (denaturation), 59°C for 40 sec (annealing) and 72°C for 1 min; then 72°C for 10 min (final extension). Products were subject to electrophoresis in 1.2% agarose gel containing 0.5 μg mL-1 ethidium bromide.
Sequencing and Sequence Analysis
Reverse transcription polymerase chain reaction (RT-PCR) products were purified from gels and sequenced by the gene-sequencing unit (VACSERA, Egypt). Identification of homologies between nucleotide and amino acid sequences of the Egyptian IBDV strains and other IBDV strains published on GenBank was done using BLAST 2.0 and PSI- BLAST search programs (National Center for Biotechnology Information (NCBI) http://www.ncbi.nlm.nih.gov/), respectively. The scores designated in the BLAST search have a well-defined statistical interpretation, making matches easier to distinguish from random background hits (Altscul et al., 1997). The obtained nucleotide sequences comparisons and their multiple alignments with reference IBDV viruses as well as the deduction of amino acid sequences were done using the BioEdit sequence alignment editor (Hall, 1999), ClustalW software for multiple sequence alignment (Thompson et al., 1994)), ClustalV (Higgins and Sharp, 1989) and MegAlign (DNASTAR, Lasergene, Version 7.1.0, USA). The phylogenetic trees were constructed using MegAlign (DNASTAR, Lasergene, Version 7.1.0, USA) for tree reconstruction of sequences by Neighbor-joining method based on ClustalW. Bootstrapping values were calculated using a random seeding value of 111 (Thompson et al., 1994). ClustalV was used when end gaps were faced. Sequence divergence and identity percents were calculated by MegAlign (DNASTAR, Lasergene, Version 7.1.0, USA).
Histopathology
Three Bursa samples were fixed in 10 % formol saline, processed by the conventional method and, stained by Haematoxylin and Eosin (Bancroft et al., 1996). The obtained slides were examined by the light microscope and scored on a scale from 1-5 based on lesion characteristics (Poonia and Charan, 2000).
RESULTS AND DISCUSSION
RT-PCR and Sequence Analysis
Extracts from tested bursal and proventriculus pools produced 620 bp amplicons. The fragment size was exactly as calculated by in silico analysis. Positive control and negative control extracts indicated primer specificity (Fig. 1). Sequencing of the PCR product was conducted in both directions and a sequence of 563 nucleotides was used for nucleotide analysis and deduced amino acid analysis. The original sequence was trimmed to remove ambiguous nucleotide sequences usually present in the beginning of the sequencing reaction. The sequence was submitted to GenBank database (Accession number: EU584433).
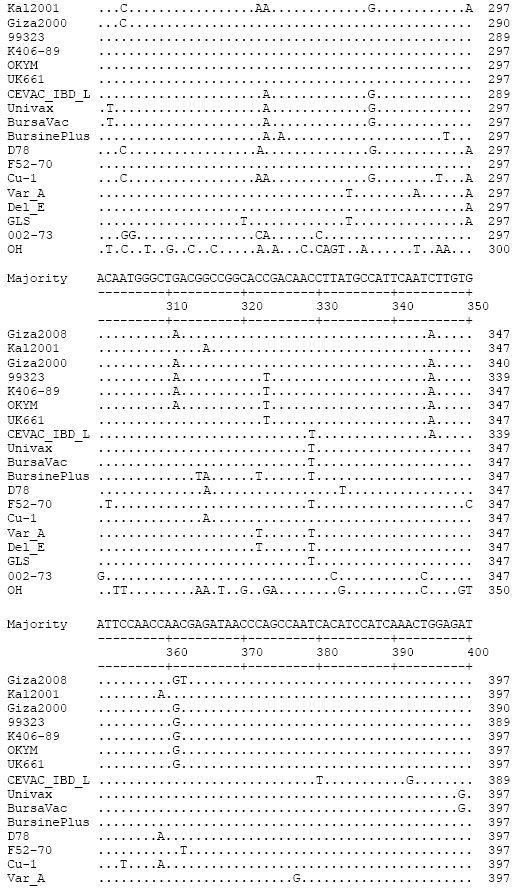
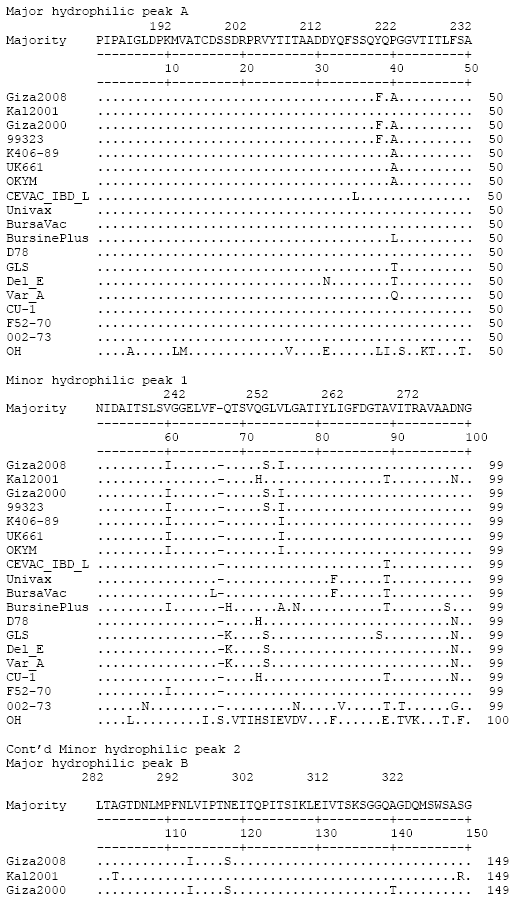
Nucleotide sequence analysis of Giza 2008 IBDV VP2 returned a 97.1% identify with 99323 and 98.9% identity with Giza2000. We were able to calculate identity between 91.8 and 93.7% comparing Giza 2008 with the available vaccinal strain sequences. Giza 2008 sequence was around 97% identical to the vvIBDV strains UK661 and OKYM. Multiple nucleotide substitutions were observed along the nucleotide sequence of Giza 2008 compared to a consensus sequence (Fig. 2).
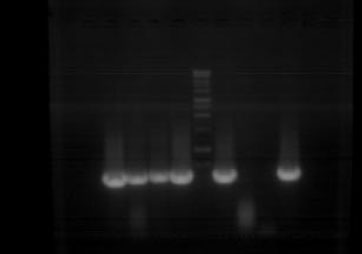 | |
| Fig. 1: | RT-PCR testing of control reference and selected samples for IBDV VP2 |
A unique substitution (C509 T) was observed. However, compared to the consensus, several other characteristic substitutions specific for Egyptian vvIBDV strains isolated after 1989 and shared with the variant strains Del/E, Variant A and GLS, were also observed [G225A, G293A, G497A]. Most of the nucleotide substitutions that characterize the vvIBDV strains were also observed in Giza 2008 (Fig. 2).
A consensus of 174 amino acids was used for sequence analysis of the deduced aa sequences of Giza 2008 [correspond to the region from aa 183 to aa 356 according to numbering of strain F52/70 (Bayliss et al., 1990)] (Fig. 3). Analysis of the deduced amino acid sequences of Giza 2008 in comparison with Giza2000 and 99323 showed that a single aa mutation (A321T) in the major hydrophilic peak B was not present in Giza 2008. However, a single aa change in the major hydrophilic peak A (Y220F) was present in 3 of 4 sequenced Egyptian strains. The vvIBDV-specific mutation (P222A in the major hydrophilic domain A) was present in all characterized vvIBDV sequences in this analysis including Giza 2008 (Fig. 3). Another mutation shared by all vvIBDV strains was observed (V256I). There were no mutations similar to any known unique variant IBDV sequences used in this comparison (Fig. 3). The aa changes lead to change in surface probability indices indicating increased probability of surface exposure in one location (around Thr250, Ser251, Val252) and sequestration from the surface in two other locations (Ser17, Ser18 and Gln19 as well as Ala321), data not shown.
The nucleotide phylogenetic tree of Giza 2008 VP2 and other reference classical, very virulent, variant and vaccinal strains of IBDV revealed that all tested reference sequences grouped together as reported previously (Eterradossi et al., 2004) (Fig. 4). The Egyptian sequences of vvIBDV Giza 2008 and Giza 2000 grouped together, however, Giza 2008 was located on a separate branch with a high bootstrap value separating both branches (Fig. 4). The European, Asian and Egyptian vvIBDV strains, isolated before 2000, grouped in a separate cluster within the vvIBDV group (Fig. 4). Phylogenetic analysis of the deduced aa sequences revealed that Giza 2008 branched separately from Giza2000 and 99323 (Fig. 5).
Pathology
The pathological alterations in the bursae collected from tested flock were more or less the same but with little differ in its degree of severity. The main lesions in the bursa were congestion of blood vessels, edema and inflammatory cells infiltrations in the interstitial tissues, mainly lymphocytes, accompanied with proliferation of the connective tissues (Fig. 6a). Necrosis of glandular epithelium was also observed. Moreover, the lymphoid follicles appeared scattered in the interstitial tissue, depleted and atrophied with presence of vacuoles in the cortical and medullar portion.
 | |
 | |
 | |
 | |
 | |
| Fig. 2: | Nucleotide sequences of the VP2 variable domain in the IBDV strain Giza 2008 and other reference classical, virulent, very virulent, variant and vaccinal IBDV strains shown in Table 1. Dots indicate position where the sequence is identical to the consensus |
There were large numbers of cyst containing serous fluids displaced and replaced the lymphoid follicles (Fig. 6b). Some follicles were converted to cysts contain eosinophilic necrotic cells and nuclear debris and infiltrated by heterophils (Fig. 6c). There was necrosis of lymphocytes and lympho-epithelial cells with presence of its nuclear debris in lymphoid follicles (Fig. 6d). The bursal lesion score were calculated for the bursae that were provided. The scores are presented as averages. The pathological finding in the flock were scored from 4-5 with an average of 4.6.
The reemergence of IBDV outbreaks in vaccinated broiler flocks despite the intensive and meticulous application of available commercial live and inactivated IBDV vaccines is a matter of great concern to poultry producers worldwide (Van den Berg, 2000; Kabell et al., 2005). In Egypt the situation is exacerbated in the absence of a dynamic vaccine production mechanism to follow up the evolving genetic and antigenic makeup of circulating IBDV. No major change in the vaccination routines has been adopted by commercial poultry producer although vvIBDV has been identified since 1989 (Zierenberg et al., 2000) and variant IBDV has been confirmed since 2003 (Hussein et al., 2003; Metwally et al., 2003).
 | |
 | |
| Fig. 3: | ClustalW multiple sequence alignment of the deduced amino acid sequences of the Giza 2008 VP2 in comparison to previously characterized Egyptian and reference strains |
Nucleic acid-based methods are useful tools for direct detection and subtyping without isolation and propagation (Stram et al., 1994). Reverse transcription polymerase chain reaction (RT-PCR) techniques on selected fragments of the genome, essentially the variable domain of VP2, followed by sequencing and phylogenetic comparison represents a valuable molecular alternative for the classification of IBDV strains (Van den Berg, 2000).
In this study we show that vvIBDV belonging to the Egyptian strains, which is in fact distantly related to the European strain, have succeeded in surviving in the Egyptian environment despite the intensive vaccination programs adapted. Others have also reported this observation (Eterradossi et al., 2004). Phylogenetic analysis shown that Giza 2008 is isolating, together with Giza2000, away from the vvIBDV that was initially identified in Egypt.
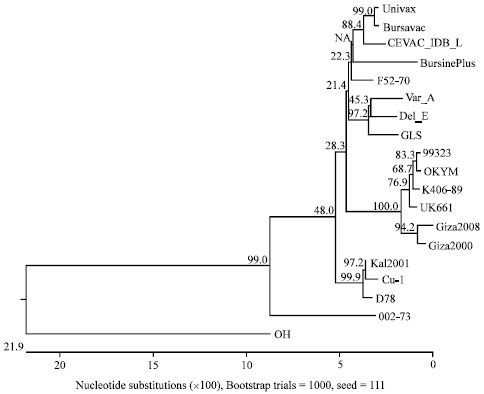 | |
| Fig. 4: | Nucleotide phylogenetic tree of Giza 2008 VP2 and other reference classical, very virulent, variant and vaccinal strains of IBDV |
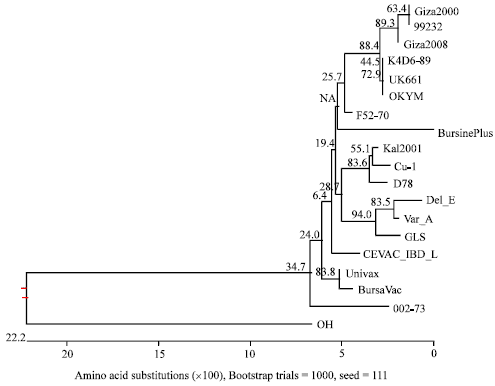 | |
| Fig. 5: | Phylogenetic tree of deduced amino acid sequences of Giza 2008 VP2 and other reference classical, very virulent, variant and vaccinal strains of IBDV |
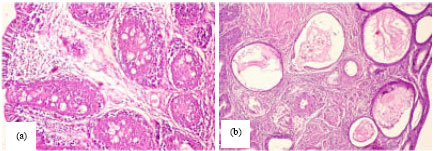 | |
| Fig. 6: | Histopathological findings of bursae recovered from dead birds from IBDV vaccinated commercial broiler flocks in Giza |
This could indicate that vaccine-directed immunological pressures are only aiding in the evolution of the virus. Giza 2008 is genetically distinct from vaccine and classical IBD strains.
The nucleotide and subsequent aa changes acquired by Giza 2008 VP2 have lead to significant changes in the folding pattern of this region of the VP2 as predicted by protein analysis (data not shown). These accumulated changes will increase chances that more neutralization escape mutants will evolve in the near future (Letzel et al., 2007). There is a threat of emergence of new vvIBDV outbreaks in the foreseeable future if current vaccination programs do not take into account the newly circulating antigenic features.
The bursal pathology recorded indicated that the lesions were not induced by any of the intermediate or intermediate plus vaccine strains in use (Bolis et al., 2003; Rautenschelin et al., 2003; Abdel-Alim and Kwakab, 2006). This was supported by our sequencing data. The retrieved viral sequences were those of vvIBDV and not related to any of the vaccines.
In conclusion, present data demonstrate the success and continuous evolution of the vvIBDV in the Egyptian environment. It also demonstrates that there is a threat of emergence of new vvIBDV outbreaks in the foreseeable future if current vaccination programs do not take into account the newly circulating antigenic features. There is an urgent need to develop dynamic mechanisms to produce local vaccines and/or methodologies to combat the inevitable reemerging IBDV mutants.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Azad, A.A., M.N. Jagadish, M.A. Brown and P.J. Hudson, 1987. Deletion mapping and expression in Escherichia coli of the large genomic segment of a birnavirus. Virology, 161: 145-152.
PubMed - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Banda, A., P. Villegas, J. El-Attrache and C. Estevez, 2001. Molecular characterization of seven field isolates of infectious bursal disease virus obtained from commercial broiler chickens. Avian Dis., 45: 620-630.
Direct Link - Bayliss, C.D., U. Spies, K. Shaw, R.W. Peters, A. Papageorgiou, H. Muller and M.E.G. Boursnell, 1990. A comparison of the sequences of segment A of four infectious bursal disease virus strains and identification of a variable region in VP2. J. Gen. Virol., 71: 1303-1312.
PubMedDirect Link - Becht, H., H. Muller and H.K. Muller, 1988. Comparative studies on structural and antigenic properties of two serotypes of infectious bursal disease virus. J. Gen. Virol., 69: 631-640.
PubMedDirect Link - Bolis, D.A., F.J. Paganini, V.A. Simon, M.F. Zuanaze, H. Scanavini Neto A.R.A. Correa and N.M.K. Ito, 2003. Gumboro disease: Evaluation of serological and anatomopathological responses in vaccinated broiler chickens challenged with very virulent virus strain. Rev. Bras. Cienc. Avic., 5: 1-13.
Direct Link - Cheville, N.F., 1967. Studies on the pathogenesis of Gumboro disease in the bursa of fabricius, spleen and thymus of the chicken. Am. J. Pathol., 51: 527-551.
Direct Link - Dobos, P., B.J. Hill, R. Hallet, D.T. Kells, H. Becht and D. Teninges, 1979. Biophysical and biochemical characterization of five animal viruses with bi-segmented double stranded genomes. J. Virol., 32: 593-605.
PubMed - El-Sanousi, A., H.M. Madbouly, M.S. Saber, C.F. El-Bagouri, N.A. Abd El-bar, A. Batrawi and I.M. Reda, 1994. Infectious bursal disease virus infection among Egyptian poultry flocks. III- Antigenic characterization of IBDV by antigen capture ELISA (AC-ELISA) using monoclonal antibodies (MAbs). Beni Suef. Vet. Med. Res., 4: 300-307.
- Eterradossi, N., C. Gauthier, I.M. Reda, S. Comte and G. Rivallan et al., 2004. Extensive antigenic changes in an atypical isolate of very virulent infectious bursal disease virus and experimental clinical control of this virus with an antigenically classical live vaccine. Avian Pathol., 33: 423-431.
Direct Link - Fahey, K.J., K. Erny and J. Crooks, 1989. A conformational immunogen on VP-2 of infectious bursal disease virus that induces virus-neutralizing antibodies that passively protect chickens. J. Gen. Virol., 70: 1473-1481.
CrossRefDirect Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hassan, M.K., M. Afify and M.M. Aly, 2002. Susceptibility of vaccinated and unvaccinated Egyptian chickens to very virulent infectious bursal disease virus. Avian Pathol., 31: 149-156.
PubMed - Kabell, S., K.J. Handberg, Y. Li, M. Kusk and M. Bisgaard, 2005. Detection of vvIBDV in vaccinated SPF chickens. Acta Vet. Scand., 46: 219-227.
Direct Link - Letzel, T., F. Coullibaly, F.A. Ray, B. Delmas, E. Jagt, A.A.M.W. van Loon and E. Mundt, 2007. Molecular and structure bases for the antigenicity of VP2 of infectious bursal disease virus. J. Virol., 81: 12827-12835.
Direct Link - Poonia, B. and S. Charan, 2000. T-cell suppression by cyclosporine- A enhances infectious bursal disease virus infection in experimentally infected chickens. Avian Pathol., 30: 311-319.
Direct Link - Rautenschelin, S., H.Y. Yeh and J.M. Sharma, 2003. Comparative immunopathogenesis of mild, intermediate and virulent strains of classic infectious bursal disease virus. Avian Dis., 47: 66-78.
Direct Link - Spatas, S.I. and J. Ignjatovic, 2000. Antigenic and sequence heterogeneity of infectious bursal disease virus strains isolated in Australia. Arch. Virol., 145: 773-785.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Van den Berg, T.P., 2000. Acute infectious bursal disease in poultry: A review. Avian Pathol., 29: 175-194.
CrossRefDirect Link - Yamaguchi, T., M. Ogawa, Y. Inoshima, M. Miyoshi, H. Fukushi and K. Hirai, 1996. Identification of sequence changes responsible for the attenuation of highly virulent infectious bursal disease virus. Virology, 223: 219-223.
PubMedDirect Link - Yousif, A.A., W.A. Mohammad, M.H. Khodeir, A.A. Abo Zeid, A. El-Sanousi, M.S. Saber and I.M. Reda, 2006. Oral administration of Hyperimmune IgY: An Immunoecological approach to curbing acute infectious bursal disease virus infection. Egypt J. Immunol., 13: 85-94.
Direct Link - Yu, L., J.R. Li, Y.W. Huang, J. Dikki and R. Deng, 2001. Molecular characteristics of full-length genomic segment A of three Infectious Bursal disease viruses in China: Tow attenuated strains and virulent field strain. Avian Dis., 45: 862-874.
PubMedDirect Link - Zierenberg, K., H. Nieper, T.P. van den Berg, C.D. Ezeokoli, M. Voss and H. Muller, 2000. The VP2 variable region of African and German isolates of infectious bursal disease virus: Comparison with very virulent, classical virulent and attenuated tissue culture-adapted strains. Arch. Virol., 145: 113-125.
Direct Link