
Orhue Ehi Robert
Department of Soil Science, University of Benin, Benin City, Nigeria
Uzu Ogbonnaya Frank
Department of Soil Science, University of Benin, Benin City, Nigeria
Osaigbovo Ulamen Agbonsalo
Department of Crop Science, Faculty of Agriculture University of Benin, Benin City, Nigeria
International Journal of Soil Science
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 1-14







ABSTRACT
This study revealed that termites are found all over the world with numerous species that have existed on this earth in the past million of years. The most prominent among these various species under different land use patterns is the Isoptera. Generally, termite activities in the soil affect the nutrient and organic matter dynamics, productivity of the ecosystem via carbon sequestration, nutrient recycling as well as some physical properties of the soil.
PDF Abstract XML References
How to cite this article
Orhue Ehi Robert, Uzu Ogbonnaya Frank and Osaigbovo Ulamen Agbonsalo, 2007. Influence of Activities of Termites on Some Physical and Chemical Properties of Soils under Different Land Use Patterns: A Review. International Journal of Soil Science, 2: 1-14.
DOI: 10.3923/ijss.2007.1.14
URL: https://scialert.net/abstract/?doi=ijss.2007.1.14
DOI: 10.3923/ijss.2007.1.14
URL: https://scialert.net/abstract/?doi=ijss.2007.1.14
INTRODUCTION
Termite is an endogenic exopterygotous insect which belongs to the order Isoptera and it is one of the numerous organisms that inhabit the soil. The abundance, composition and hence their impact on soil processes vary greatly depending on vegetation and land use (Ayuke et al., 2004). The management practices such as continuous land preparation can cause alteration in the population structure, elimination and reduction of key species (Dangerfield, 1993). In terms of number, termites and ants predominate under agro-ecosystem (Bruno et al., 2001) and in terms of diversity termite group are in abundance in natural sites than the cultivated sites (Abe and Matsumoto, 1979; Collins 1979; Kondoh et al., 1980; Leaky and Proctor, 1987; Atkin and Proctor, 1988; Lavelle and Pashanarv, 1989; Fragoso and Lavelle, 1992; Dangerfield, 1993; Frageria and Baliga, 2004).
Generally, termite thrives by consuming detritus material and building mounds in which they control their environment. Depending on the species of termites and the surrounding environment, the mounds take on a variety of shapes and sizes. These mounds illustrate how termites are well suited for the environment. Construction of mounds from soil or mixture of soil and other material or within soil horizons must affect the physical and chemical characteristics of both the soil used for construction and the soil of the surrounding areas from which the materials are derived (Lee and Wood, 1971). During the period of occupation of the mound, organic debris or living plant tissue is collected, often over extensive foraging areas, transported to mounds and subjected to intense degradation when it is digested by termite. Plant nutrients and organic matter that they contain are withheld from circulation in the plant-soil system until they finally decay. Termite activities in the soil therefore affect the nutrient and organic matter dynamics and structure of soil. Such changes in soil properties have profound influence on the productivity of the ecosystem via carbon sequestration, nutrient cycling and soil texture.
Within the savanna and rainforest ecosystem, termites bring about an important change on the soil environment and can occupy a large portion of the land (Leed and Wood, 1971). Knowledge about the impact of termites on soil fertility is not only important for understanding the ecology of tropical system but also for evaluating the potential constraints for agricultural production in our fields (Bruno et al., 2001).
Some results of the research work carried out on physical and chemical changes in soil due to termite activity are difficult to relate to soils in our environment under different land use pattern. The elements estimated vary from one investigation to another and some of the studies lack the identification of the termites responsible for the mound structures. Therefore, this review will enable us understand the activities of termite in our soil as it affects both physical and chemical components of the mounds, the surrounding soils as well as nutrient dynamics in termite infested soils under different land use systems.
TERMITE BACKGROUND
Malaka (1996) reported that there are about 2100 species of termite while Kambhampati and Eggleton, (2000) noted that there are close to 3000 species of termite in the order of Isoptera that are phylogenetically separated into lower and higher termites. These orders according to Kambhampati and Eggleton include lower termite such as Mastotermitidae, Kalotermitidae, Hodotermitidae, Termopsidae, Rhinotermitidae and Serritermitidae while the Termitidae is the higher family of termite. Termites are further divided into either wood-dwelling or soil dwelling and the majority of the soil dwelling termites construct either above nests or dig underground galleries while the wood-dwelling termites do not create nests but live in excavated galleries in the hollow of trees (Ratcliffe et al., 1952; Malaka, 1996). Lee and Wood (1971) and Malaka (1996) reported that it is the Termitidae family that makes up the mound forming termite.
The sub family of Termitidae according to Pearce (1997) includes Macrotermitinae, Apicotermitinae, Termitinae and Nasutitermitinae while the species associated with these subfamilies are shown in Table 1.
According to Ratcliffe et al. (1952), Krishna and Weesmer et al. (1969) and Kambhampati and Eggleton (2000) all known species of termite live in colonies that are compose of distinct caste namely the workers, soldiers, nymphs, larvae and reproductive (King and Queen). Each caste according to Kambhampati and Eggleton (2000) has a specific role that contributes to the productivity of the colony. The workers represent the majority of termite in a colony; they build the nest or mound, take care of the eggs and the young ones and gather food while the soldiers protect the community from predators. The nymphs are immature ones that may develop into adults whereas the larvae are the immature ones of the reproductive and the reproductives are the actively reproducing males and females.
| Table 1: | Species associated with various termitinae subfamily |
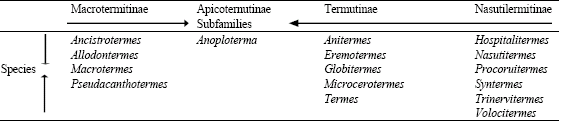 | |
| Source: Pearce (1997) | |
GEOGRAPHICAL DISTRIBUTION OF TERMITES
According to Tsukamoto and Sabang (2005), climate and zoogeography are the major determinant of termite distribution at global level. The great majority of termites live in tropical and subtropical regions but they extend into the temperate zone to about 45°N (Harris, 1970) or 48°N (Emerson, 1955) and about 45°S (Araujo, 1970). Termite distribution can be related to temperature and rainfall. The distribution changes with latitude and the limits of survival are between latitude 45 and 50° north and south (Pearce, 1997).
Within the tropics, many species in Africa and South America are found in rainforests (Lee and Wood, 1971), whereas in Australia only four species are known to be restricted to tropical rainforest and the majorities are found in the sclerophyll forest, woodland and savanna (Gray and Calaby, 1970). Termites are also known to have reached Hamburg, Germany (Pearce, 1997) and in Southern portion of Canada (Grace, 1989, 1990). Some of the genera with old origin such as Zootermopsis according to Pearce (1997) are in British Columbia, Porotermes at tip of Chile, Tasmania and South East Australia and the Archotermopsis in Himalaya, Mastotermes in Australia and Papua New Guinea, Heterotermes in the hotter regions of South Western USA, Central America, Caribbean, India while the Reticulitermes are common in Canada, Middle East, Japan, China, Coptotermes are common in tropical America, Africa, Far East, Pacific Island and Japan; the lower termites are common in paleartic, neartic and Australia south of the tropic of Capricorn. Pearce reported further that the higher termite family of Termitidae occurs mainly in tropical latitude with the fungus growers and humus soil-feeders found in Ethiopia and oriental regions.The macrotermitinae are mainly found in Africa, Middle East and Asia.
Malaka (1996) while citing Harris (1970) reported that harvester termites are common throughout subtropical dry grassland areas where they compete with man’s flock for the sparse grazing. Abe and Matsumoto (1979), Kondoh et al. (1980) and Lavelle and Pashanan (1989) recorded that abundance of termites is common to tropical lowland forests In Africa, termites are widespread, restricted mainly by desert areas and lower temperature found at higher altitude. Lee and Wood (1971) while citing Grasse (1950) noted that fungus feeding termite were rare or absent from dry areas of West Africa and that their preferred habitat being humid savanna and forest. Kemp (1955) reported a similar distribution in East Africa where the humid woodland near the coast supported many more species and greater proportion of mound builders than the dry inland areas. In Nigeria as in other parts of West Africa termite representatives of only three family according to Sands (1965a), Malaka (1973, 1983) include Kalotermitidae, Rhinotermitidae and Termitidae. In the mangrove and freshwater swamp of Nigeria, Ancistrotermes, Cryptotermes, Marotermes subhejahnus, Odontotermes, Microcarotermes and Annitermes evuncifer have been reported to be common Malaka (1977a, b) whereas in the rainforest area. Malaka (1996) reported that there are more diverse species of termites in the zone as a result of favourable climate and vegetation. In the savanna as well as Sahel zones according to Malaka termite species identified include Macrotermes bellicosus, Mmacrotermes subluyallinus, Ttrinervitermes species.
COMPONENTS OF CONSTRUCTED TERMITE MOUNDS
According to Wood and Sands (1978), termite nest systems may be arboreal, either within or attached to the outside of the vegetation epigeal or subterranean. Depending upon the species and location (Ratcliffe et al., 1952; Abensperg Traun and Perry, 1998) and moisture availability and environmental conditions (Pearce, 1997) termite mound can take a variety of forms and that in colder regions mounds may be underground and in arid regions, the mound may become smaller to reduce water loss. Pearce observed further that Armitermes evuncifer species in West African can be subterranean in dry areas and above ground in wet regions. The nests or mounds according to Malaka (1996) and Lee and Wood (1971) consist of four main types of structures based on material from which mounds are composed and they include
| • | Fabric dominated by re-packed orally transported soil particles without admixture of excreta such as large mound of Macrotermitidae. |
| • | Fabric dominated by re-packed orally transported soil particles with admixture of excreta constructed by Coptotermes species and Trinervitermes species. |
| • | Fabric dominated with excreta consisting of soil mixed orally with transported soil as seen in the mounds of Odontotermes species. |
| • | Fabric dominated by excreta consisting of organic matter derived from ingesting of plant materials as seen in Amitermes species and Nasutitermes species. |
Malaka (1996) reported that the construction of these mounds result in the transport of soil from either deep or shallows horizons or both to the soil surface where it is formed into mounds and covered runways on the soil surface and over or within vegetation on which the termites are feeding. Pearce (1997) reported that mounds are usually made after colony developed below the ground and that Amitermes, Cubitermes and Procubitermes are often mushroom shape with dry tips to help remove rain quickly while the mound of Cubitermes species often do not have this umbrella cap.
In Nigeria according to Collins (1979) the Macrotermes may have two kinds of mounds in similar conditions. One has a spiral plate supported by a central pillar; the other does not have this arrangement and the spiral plate has downward directing vanes that are covered in salt deposits. The magnetic mounds of Amitermes in Australia according to Pearce (1997) are wedge shaped and reach 5.5 m in height with a width of only 0.9-1.2 m whereas the Macrotermes goliath can attain a width of 12 m at least 4.8 m high and can number as many as 17 ha-1. Odontotermes mounds are larger in India than in Africa and species differences also occur between mounds in India: Odontotermes Obesus mounds are conical while Odontotermes wallanensus are dome-shaped (Pearce, 1997).
Lee and Wood (1971) recorded that the termite mounds range in size from small domed to conical structures, only a few centimeters in height and diameter to the colossal mounds built by some species of African Macrotermes which reach 9 m or more in height and 20-30 m in diameter at the base. Harris (1955) recorded a mound of Macrotermes species in Ethiopia of approximately 9 m in height and that of Nasutitermes triodiac in Australia to be above a height of 8 m. Merdraci and Leppage (2001) reported Odontotermes in Lampto Savanna to be the main contributor in building large mounds of varied sizes 2-20 m in diameter with 10 cm-2 m in height. In terms of space occupied and amount of soil deposited on the surface, the species of Macrotermitinae reported by Meyer (1960) in Congo and that of Macrotermes bellicosus (Nye, 1955) were 240,0000 and 306000 kg ha-1, respectively. According to Lee and Wood (1971), the weight of mounds and area of ground covered by termite mound varies with species and localities and that the space occupied by these mounds has significant effect on the landscape, vegetation and local agricultural practices.
The mounds in various ecosystems according to Lee and Wood (1971) however are not static objects but are continually being eroded by rainfall and occupied mounds are repaired, enlarged by the termites while abandoned mounds are gradually eroded away.
TERMITE ABUNDANCE BASED ON LAND USE PATTERNS
The presence of termite in a place depends on various factors such as soil, vegetation type as well as climatic features (Pearce, 1997). It is often not possible to say which factor is most important on basis of observational evidence alone (Lee and Wood, 1971). Tsukamoto and Sabang (2005) reported that at local level, the interrelated two factors, soil condition and litter quality, are most important and that land use practices disturb both levels of regulation. At the initial stage of the conversion practice of natural forest to tree plantations, clear felling, site preparations, introduction of seedlings from outside seriously damage the indigenous soil biota. Such perturbation and/ or the establishment of herbaceous stratum could provide niches that allow some exotic species to colonize (Fragoso et al., 1999; Pellens and Garay, 1999; Muys et al., 1992; Yeates, 1988). Termite mounds are relatively easy to count and the densities of the large mounds of Macroternutinae in Africa and of Nasuititermes triodiae in northern Australia which support colonies of several million individuals are usually less than 10/ha while densities of the smaller mounds which support colonies of several thousand individuals may approach 1000 ha-1. High densities of the large mounds are reached only in Savanna or sparsely timbered shrub or tree savanna while smaller mounds can be extremely abundant in both savanna and dense forest and there are few estimates of the abundance of mounds of all species in selected habitat (Lee and Wood, 1971). Sands (1965) observed that the number of mounds may approach or even exceed 1000 ha-1 in the rain forest areas of West Africa. Glover et al. (1964) reported an average of 5-7 ha-1 in Kenya savanna while Bouillon and Kidieri (1964) reported 2-3 ha-1 in Congo savanna of both Odontotermes species and Macrotermes bellicosus. In the tropical rain forest of Congo Maldague (1964) reported 875 ha-1 of Cubitermes fungiflabe species mounds.
Parkr et al. (1994) noted that the Woodoo Woodland appeared to be more optimal for Drepanotermes tamminensis species than the casuarinas shrubland in Western Australia Wheatbelt. In agro forestry, in Tororo District of Uganda, Nyeko and Olubayo (2005) reported abundance of Macrotermes bellicocus and Macrotermes sabhyalumus. In a survey conducted in Nakasongola District of Uganda and Central Uganda, Sekamatte (2001) recorded Ancistrotermes, Allondontotermes, Armitermes, Cubiternes hodotermes, Macroterma, Microtermes, Odontotermes and Pseudocasthotermes as most abundant species.
Nel and Malan (1974) noted that mounds of Trinervitermes trinervoides were not randomly distributed over the veld in South Africa but tended to aggregate in certain area, while Nyeko and Olubayo (2005) observed that Macrotermes bellicocus were rare in valley in the agroforest in Tororo District of Uganda. Cowie et al. (1989) and Sands (1998) recorded that Macrotermes, Odontotermes mounds were associated with land cultivated with wide range of field crops and trees. Pearce et al. (1995), Sands (1998) and Sekamate (2001) discovered the Pseudocanthotermes termite mounds to be found in various field crops and nursery plot or newly transplanted tree seedlings. Amitermes evuncifer termite mounds have been found to be commonly seen in field of root crops such as yam, cassava and some tree crops (Sand, 1998).
According to Sands (1998) Trinervitermes species have been reported to be common in cotton field, grass crops including sugar cane, upland rice and wheat whereas Trinervifermes trinervoides mounds were reported to be common in pastures in South Africa. Tsukamoto and Sabang (2005) reported that termite mounds were significantly lower in Acacia mangnum plantation than the primary forest. Lee and Wood (1971) showed that in Frankenwald, South Africa, the abundance of termite mounds increased as secondary plant succession advanced from two year fallow to undisturbed veld. Ekundayo and Aghatise (1997), Brouwer et al. (1991), Dangerfield (1990), Lal (1998) and Lobry de Bruyn and Conacher (1990) reported that the number of termite mounds were greater in order of fallow greater than residential greater than cropping while on the other hand Ekundayo and Orhue (2002) found that the number of termite mound per unit area increased from cropping through the riparian forest and the palm plantation to residential area.
Soil feeders have been reported by Pearce (1997) to be abundant in humid forest of Africa and South America. According to Pearce, as areas of low, open vegetation give way to dense forest, the species of Macrotermitinae increased.
In the lowland forest in Sarawak, high numbers of soil feeding termite mounds were seen while in the continuous forest of Amazonia, greater species richness was found than the scattered forest (Martins, 1994). The removal of trees and bushes followed by cultivation resulted in loss of Macrotermes, cubitermes, trinervitermes and shallow mound of Coptotermes in Nigeria (Pearce, 1997), Black and Wood (1989) reported that some subterranean species such as Microtermes increased in numbers as they could move down into the soil to avoid the effect of surface disturbances. The clearing of primary rainforest in Malaysia and Indonesia for cultivation allowed termites such as Coptotermes to survive where tree stumps remains (Eggleton et al., 1995).
Sands (1965b) in his study of distribution and abundance of termite mound in Northern Nigeria reported high densities of termite mound in an area with about 90% exposure. Kemp (1955) found that in the dry thickets of inland Tanzania, termite mounds only occurred in shaded tree sites. In semi-desert where little vegetation is present, Psanmotermes is often found underneath small rocks (Pearce, 1997). Bodot (1964, 1967) who studied the interaction of various factors observed that there were more termite mounds in area with more clay and low water holding capacity than area with high water holding capacity. Soils with seasonal cracking as noted by Ratcliffe et al. (1952) do not accommodate termite mounds. Meyer (1960) in Congo recorded much termite mound in soils with high clay content while Lee and Wood (1971) reported high termite mounds in soil with higher depth compared to shallow soil in Australia in savanna woodland. In sandy soil few mounds according to Pearce (1997) were present especially of humus-feeding termites and that of desert of Hodotermitidae which have mound than other regions. Jones (1988), Nutting and Jones (1990) and Grace (1992) recorded that true desert dwelling termites such as some Armitermes and Psammotermes species are found in sandy and gravelly soils whereas Trinervitermes species are absent in extremely dry arid conditions but commonly seen in semi-arid and wet open areas.
Where there is a rich well drained grassland such as in Ivory Coast, Pearce (1997) reported that humivorous and fungus-growing termites are common while in poor drainage areas grass-feeders such as Trinervitermes and Macrotermes species are common.
Lee and wood (1971) while citing Williams (1966) reported that in Africa species of Cubitermes were abundant in regions of heavy to moderate rainfall, rare in regions of light or more season rainfall and absent from regions with annual rainfall of less than 250 cm. The effect of a relatively rapid change in climate over a short distances can be seen in reduced population of the termite fauna with increasing attitude as illustrated in Kemp (1955) study in Tanzania where Coptotermes lacteus - a common mound-building species in Woodland and sclerophyll forest on the eastern plateau, rarely occurs above 1200 m and the even more abundant Nasutitermes exitiosus rarely occurs above 900 m. Termites of different genera or different species of same genus can have its differing temperature tolerance as discovered in the behaviour of Coptotermes formosanus that have higher temperature tolerance (Sponsle and Appel, 1991).
CHEMICAL AND PHYSICAL PROPERTIES OF TERMITE MOUNDS
The modification of soil profiles activity depends on the fact that termites remove soil from various depths and bring it to the surface in form of covered runways or mounds from which it is distributed by water and wind erosion (Malaka, 1977a). The significance of physical disturbance of soil profile depends partly on the nature of the soil brought to the surface by the termites and partly by the physical and chemical changes in the soil due to the activities of the termites. Chemical changes are brought about by the incorporation of organic matter while physical changes appear to be due to selection and sorting of certain particles resulting in a change of structure and particle size distribution (Malaka, 1977a, b). The changes in texture brought about by redistribution of mounds and other structures in the surface is likely to be accompanied by changes in physical properties such as structural stability, bulk density, infiltration rate, permeability and water holding capacity (Wood and Sands, 1978). Studies by Pomeroy (1983), Arshad et al. (1982), Wood et al. (1983), Menant et al. (1985) and Abhadie and Lepage (1989) showed that termite play a major role in both physical and chemical properties of soil as well as nutrient cycling and soil metabolism.
PHYSICAL CHANGES AND TERMITES MOUNDS
According to Lee and Wood (1971), the particle size of soil from various termite mounds when compared with adjacent soil showed that some degree of selection of particular size fraction is general and that few termite species consistently select material within a precise range of particle size.
Nye (1955), Harris (1956), Stoop (1964) and Grasse (1984) noted that particle size of mounds of Macrotermes species in Africa showed that their composition is close to that of subsoil, but there is some selection in favour of fine size fraction in soils that are not rich in clay or against the finer fraction in soils rich in clay at Arush, Tanzania. The changes in Macrotermes mound shape in Eastern Africa were related to the ratio of sand to clay in mounds and the subsoil from which materials were obtained (Hesse, 1955). Hesse (1955) reported further that tall, thin mounds had sand to clay ratios of 1:1 to 3:1 while larger, domed-shaped mounds had ratios of 2:1 to 18:1. According to Kemp (1955), Cubitermes species mounds had 67.2% clay and 26.5% sand compared with 30.8% clay and 63.0% sand for the surrounding soils. Stoop (1964) recorded that the mounds of Cubitermes scinteurensis had low proportion of 100-500 μ particle size and higher proportion of less than 100 μ particle size of soil compared to surrounding soil with higher values.
Pathak and Lehri (1959) found that in the termite mound soil, 67.7% of clay aggregates had particle size greater than 2 μ compared with 48.1% in undisturbed soil and 19.2% of silt size particles were incorporated in aggregates in the mound soil while 5.6% were aggregates in undisturbed soil. Watson (1962) reported that the termite mound had 94% fine material (clay + silt) compared to 52% clay and silt in adjacent soil in West Africa. When the physical properties of termite modified soil by Drepanoteormes tammunensis, Amitermes obenntis and Amitermes neogermams were examined; Debruyn and Conacher (1987) found that the mounds had significantly higher clay content than the surface surrounding soil. Kang (1978), Lal (1988), Breuwer et al. (1991), Malaka (1977), Ekundayo and Aghatise (1997) and Ekundayo and Orhue (2002) observed that the various termite mound soil had higher values of silt and clay and lower values of sand in relation to bulk surrounding soil from which the mounds were constructed. Ghilarov (1962) describe the termite mound of Anacanthotermes species to be strongly cemented with clays while Greaves (1962) observed the termite mound of Soptotermes brunneus to consist of compact red sandy soil in Western Australia. In a comparison of granulometry between mound and the surrounding soil Merdraci and Lepage (2001) reported higher increase of finer soil particles (clay + silt) than the surrounding soil in Lamto Savanna. Lobry de Bruyn and Conacher (1990) however observed that variation in particle size was due to variation in site characteristics, termite species and genera, land use at the sampling site.
Ghilarov (1962) reported the bulk density of termitaria to be 1.112 g cm-2 compared with 1.225 g cm-3 for surrounding soils. Malaka (1977), Ekundayo and Aghatise (1997), Ekundayo and Orhue (2002) and Reddy and Rapu (2005) reported that the bulk density of surface mound were higher than that of the surrounding soils. Malaka (1977b) however reported that the mound soil of three species of Cubitermes, Macroyermes, Trinerviterma germinatus which do not support vegetation had an average bulk density of 1.80 g cm-3 or greater with accompanying porosity less than 33% while mound material of Amitermes evuncifer which commonly support vegetation had an average bulk density of 1.33 g cm-3 and a porosity of 48%.
Pathak and Lehri (1959) reported that mounds of Odontotermes abscuriceps in India had a water-holding capacity about five times than that of adjacent soil. It was reported by Elkins and Sabadol (1986) and Jauean and Valentin (1987) that the surrounding soil had less water-holding capacity relative to the various termite mounds examined. Kemp (1955) have earlier reported a small increase in water holding capacity in mounds of Cubitermes umbratus in Africa compared to the adjacent soil. Ghilarov (1962) reported a fall in water-holding capacity of about 8% in mounds of Anacthotermes species relative to surrounding soils in central Asia while Goodland (1965) found a reduced water holding capacity in an unidentified termite mound compared with the surrounding soil in Rupunmu savanna. Lal (1988) found low water infiltration in mound soil in relation to bulk surrounding soil. Arshad (1982) reported favourable water availability together with good drainage in termite mound soil compared to the surrounding soil.
CHEMICAL CHANGES AND TERMITES MOUNDS
Lal (1988) and Brouwer et al. (1991) reported that mound soil of various termite species can have either higher or lower values of organic carbon total nitrogen, available phosphorus, exchangeable calcium, magnesium and potassium, effective cation exchange capacity in relation to surrounding soils. It has been described by Lavelle et al. (1994) that pedogenesis, organic matter decomposition and nutrient cycling are highly influenced by termites.
Ekundayo and Aghatise (1997), recorded a higher organic carbon, C/N ratio, Ca, Mg, K, P in mounds of Macrotermes and Odontotermes species than the surrounding soils in Nigeria. Also Reddy and Rapu (2005) recorded increased organic matter in termite mound compared to adjacent soil of Podili and Talupula in India. Frageria and Baligar (2005) reported that termite activities significantly increased exchangeable cations, micro-nutrients, organic matter content and the pH of the mound soil while the soil acidity in terms of Al was decreased in an oxisol of cerrado region of Brazil. Lobry de Bruyn and Conacher (1990), Debruyn and Conacher (1987) and Duponnois et al. (2005) found that the termite mounds of Cubitermes and Trinervitermes have higher carbon and N, CEC, NH4 and P while Macrotermes subhyalinus mound however have lower NO3 and carbon content compared to the surrounding soil in Burkina Faso.
Malaka (1997a) however, recorded low organic carbon content but high Ca, K and Mg in Macrotermitanae as compared to their surrounding soils. In the Cerrado, Brazil soil carbon stabilization was higher in termite mound as demonstrated by higher carbon content in silt size separates (Zech et al., 1997). Hesse (1955) found low C/N ratio in the termite mound in comparison to adjacent soils. Kemp (1955) using Cubitermes mound and Stoop (1964) using Macrotermes mound recorded higher organic matter content, P, cations, pH in both mounds, low pH in Macrotermes. Lee and Wood (1971) found that the organic carbon content of mounds and other structures parallel the organic matter content that is it is generally higher and occasionally lower than that of adjacent soil. Similarly, nitrogen content according to Lee and Wood (1971) was roughly parallel to organic matter content but not necessarily in the same proportion as carbon content. Lee and Wood (1971) in their investigation in Australia found that exchangeable Ca, Mg, K were generally high in termite modified soils than unmodified samples. Lee and Wood reported further that CaCO3 were occasionally found usually in a band near the base of the mound and sometimes the quantity may be sufficient that the mound may be used as a fertilizer in the initial agricultural system. Holt and Lepage (2005) and Marisa (2005) reported increase in carbon and nitrogen content of various termite mounds as compared to the surrounding soils. In the analysis of Anacanthotermes mound in Central Asia Kozlova (1951) reported 0.85% NO3 in the mound compared to adjacent soil with 0.022% NO3 and 47 kg ha-1 readily available nitrogen.
Jones (1973) and Cabrera (1993) recorded the lowest carbon content and lowest C/N ratio in the soil of Macrotermes and Odontotermes mounds. Bruno et al. (2001) however found that the largest increase of mineralize carbon took place but with lower pH in the soil of termite mound of Macrotermes in Northern Tanzania. Duponuois et al. (2005) while citing Ndiaye et al. (2003) reported that termite mound exhibited higher %C and K content in Burkina Faso. Among the African species, Boyer (1956) found that Amitermes species had a C/N ratio of 18.7-18.82 accompanied with high pH compared with C/N ratio of 8.26-10.31 for surrounding soil while Maldugue (1959) reported that the mound of Thoracotermes brevinatus had C/N ratio of 12 compared with 15.0 in the adjacent soil. Bernhard-Reverret (1982) found lower N content in the mounds of Macrotermes and Odontotermes species than the surrounding soil. Of the 46 samples consisting mainly of soil taken from mounds of 13 species only 2 samples had C/N ratio lower than the soils from which they were derived, while the remaining samples consisting of carbon or containing high proportion of carbon had C/N ratio much higher than the surrounding soils.
It has been reported by Pearce (1997) that Trinervitermes, Hodotermes species mound had great levels of N, Silica, Ca, Al, Fe, Mg, Cr, while the Odontotermes species had higher ferric and aluminum oxide, CaCo3 than the adjacent soils. Pearce also reported further that the Russian Anacesthotermes ahalgerianus had higher water soluble salts, phosphorus and potassium salts while in Okarago Delta, various termite mounds have been discovered to be rich in calcite crystals.
Watson (1962) found high concentration of Ca and high pH in un-identified species of African Macrotermitidae. Goodland (1965) found high Ca and Mg concentration in termitaria with Ca concentration averaging more than three times and sometimes up to seven times than those in the surrounding soil in the Repununi Savannah lands of Guyana. Gokhale et al. (1958), Pathak and Lehri (1959) reported high concentration of Ca, Mg, P in Odontotermes species and Microcerotermes species mounds respectively but Pathak and Lehri (1959) however recorded low K content in Odontotermes species mound. Ellis (2001) also found high content of Ca, high pH and P as well as high content of CaCO3 in the termite mound of Microhodotermes viator in South Africa.
The abundance of the CaCO3 according to Ellis (2001) has equally led to the presence of petrocalcic horizon in the soil profile. Donova et al. (2001) discovered high content of Kaolinite and higher pH and a decrease relative amount of quartz in the termite mound of Cubitermes fungifaber in humid forest of Southern Cameroon. Siame (2005) concluded in his investigation that termite mounds have high levels of Ca, P, K as well as higher amount of organic matter which contribute to better crop development. In the examination of mounds of species in northern Tanzania, Bruno et al. (2001) reported that the termite mound had low Zn but higher Na and Cu contents than the native and degraded savanna soils with higher basic cations than the termite mounds.
CONCLUSION
From the review, termites have been known to exist for several years and its study only known within past century. Termites are known all over the world and several families and species exist. The most prominent among these termites is the Isoptera. Generally, termites are the principal decomposer of organic materials thereby helping to recycle nutrient in our soils. Their role as nutrient recycler in our environment helps to balance nutrients in the soil. The particle size selection aids the re-arrangement of soil which brings about an improvement in some physical properties.
REFERENCES
- Abensperg-Traun, M. and D.H. Perry, 1998. Distribution and characteristics of mound-building termites (Isoptera) in Western Australia. J. Royal Soc. Western Aust., 81: 191-200.
Direct Link - Ayuke, F.O., M.R. Raam, M.J. Swilt and M.L. Oponda-Mbai, 2004. Assessment of Biomass Transfer from Green Manure to Soil Macrofauma in Agro-ecosystem Soil Macrofauma Biomas. In: Managing Nutrient Cycles to Sustain Soil Fertility in Sub-Shara African, Andre, B. (Ed.). Academy of Science, Nairobi, pp: 411-422.
- Bouillon, A. and S. Kidieire, 1964. Repartition Des Termitieres De Bellicostermes bellicosus Nex Grasse et Noisof Dans I'ubange D'pre's Les Photos Aerienues Correlations Ecologiques Qu=elles R`ev`ele. In: Efudes Surles Termites Africans, Bouillon, A. (Ed.). Leopoldville University, Leopoldville, pp: 373-376.
- Brouwer, J., S.C. Geiger and R. Vandenbelot, 1991. Likely positive effects of termites on soil fertility and tree and crops growth near Niamey, Niger. Would tillage reduce these? Proceedings of the 12th International Conference of International Soil Tillage Research Organization, July 8-12, 1991, Ibadan, Nigeria, pp: 82.
- Bruno, G., L. Johannes and F. Maike, 2001. Carbon and nitrogen mineralization in cultivated and natural savanna soils of Northern Tanzania. Biol. Fertility Ssoil, 33: 301-309.
Direct Link - Cabrera, M.L., 1993. Modeling the flush of nitrogen mineralization caused by drying and riveting soils. Soil Sci. Soc. Am. J., 57: 63-66.
Direct Link - Dangerfield, J.M., 1990. Abundance, biomass and diversity of soil macrofauna in savanna and associated managed habitats. Pedobiologia, 34: 141-150.
Direct Link - Donovan, S.E., P. Eggleton, W.E. Dubbin, M. Batch and L. Dibog, 2001. The effect of a soil-feeding termite Cubitermes fungifaber (Isoptera: Termitidae) on soil properties: Termites may be an important source of soil microhabitat heterogeneity in tropical forest. Pedobiologia, 45: 1-11.
Direct Link - Dupponois, R., M. Peougy, J. Thwuou, D. Masse and M. Lepage, 2005. Functional diversity of soil microbial community, rock phosphate dissolution and growth of Acacia seyal as influenced by of grass-litter and soil-feeding termite nest structures amendments. Geoderma, 124: 349-361.
Direct Link - Eggleton, P., D.E. Bignell, W.A. Sands, B. Waiter, T.C. Wood and J.H. Lawson, 1995. The species of richness of termites (Isoptera) under different levels of forest disturbances in the Mbalaiyo forest Reserve, Southern Cameroon. J. Trop. Ecol., 11: 85-98.
Direct Link - Ekundayo, E.O. and V.O. Aghatise, 1997. Soil properties of termite mounds under different land use types in a Type paleudult of Midwestern Nigeria. Environ. Monit. Assess., 45: 1-7.
Direct Link - Fragoso, C. and P. Lavelle, 1992. Earthworm communities of tropical rainforest. Soil Biol. Biochem., 24: 1397-1408.
Direct Link - Fragoso, C., I. Lavelle, E. Blanchart, R.K. Senapati and J.J. Jimenetz et al., 1999. Earthworm Communities of Tropical Agro Ecosystems Origin, Structure and Influence of Management Practices. In: Earthworm, Management in Tropical Agro Ecosystems, Lavelle, P., L. Brussaard and P. Hendrix (Eds.). CABI Publishing, Oxon, pp: 27-55.
- Kondoh, M., H. Watanabe, S. Chiba, T. Abe, M. Shiba and S. Saito, 1980. Studies on the productivity of soil animals in Pasoh Forest Reserve, West Malaysia. Malaysia. Seasonal Change in the Density and Biomass of Soil. Macrofauna Oligochaeta, Hirudinea and Arthropoda. Memo graph of Shiranme Grakuen College, pp: 1-26.
- Martins, C., 1994. Diversity and ecology of termites in Amazonian forests. Pedobiologica, 38: 407-428.
Direct Link - Muys, B., N. Lust and P.H. Granval, 1992. Effects of grassland afforestation with different tree species on earthworm communities, litter decomposition and nutrient status. Soil Biol. Biochem., 24: 1459-1466.
Direct Link - Nutting, W.L. and S.C. Jones, 1990. Methods for studying the ecology of subterranean termites. Sociobiology, 17: 167-190.
Direct Link - Pellen, R. and L. Garay, 1999. Edaphic macro arthropod communities in fast growing plantations of Eucalyptus grandis Hill ex maid (Myrtaceae and Acacies mangium wild (Leguminosae) in Brazil. Eur. J. Soil Biol., 35: 77-89.
Direct Link - Tsukamoto, J. and J. Sabang, 2005. Soil macro-fauna in an Acacia mangium plantation in comparison to that in a primary mixed dipterocarp forest in the lowlands of Savawala, Malaysia. Pedobiologia, 49: 69-80.
Direct Link - Pearce, M.J., J.W.M. Logan and A. Tiben, 1995. Termites (Isoptera) from the Darfur region of the Sudan with comments on their pest status. J. Arid Environ., 30: 197-206.
CrossRef