
R. Ramjegathesh
Department of Plant Pathology, Tamil Nadu Agricultural University,Coimbatore-641 003, India
E.G. Ebenezar
Agricultural college and Research Institute (TNAU), Killikulam-628 252, Tamil Nadu, India
International Journal of Plant Pathology
Year: 2012 | Volume: 3 | Issue: 2 | Page No.: 34-44







ABSTRACT
Ten isolates of A. alternata causing leaf blight disease of onion were collected from ten different conventional onion growing areas of Tamil Nadu and their pathogenicity was established. The species was identified by ICAR as Alternaria alternata (ITCC-Indian Type Culture Collection No. 5470). All the isolates produced light brown conidia with muriform shape, but the isolates varied in the size (length, width and beak length) of the conidia, the number of cells per conidium and sporulation times. Among the solid media tested host leaf extract agar and modified czapek's dox medium increased the growth of mycelium followed by potato dextrose agar medium and carrot agar medium. The maximum growth was observed in maltose followed by glucose, sucrose and fructose. Among the nitrogen sources, potassium nitrate followed by sodium nitrate and ammonium molybdate increased the growth of mycelium. Low pH (4-4.5) was found as ideal for the growth of A. alternata, while the minimum growth was observed in pH 9.0, irrespective of isolates. The conidia of A. alternata isolates germinated at 46°C.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: March 30, 2012;
Published: June 05, 2012
How to cite this article
R. Ramjegathesh and E.G. Ebenezar, 2012. Morphological and Physiological Characters of Alternaria alternata Causing Leaf Blight Disease of Onion. International Journal of Plant Pathology, 3: 34-44.
DOI: 10.3923/ijpp.2012.34.44
URL: https://scialert.net/abstract/?doi=ijpp.2012.34.44
DOI: 10.3923/ijpp.2012.34.44
URL: https://scialert.net/abstract/?doi=ijpp.2012.34.44
INTRODUCTION
Onion (Allium cepa var. aggregatum L.) the queen of kitchen, once considered as the poor man's staple spice, is flying out of the reach of even middle class families. Now it is used as salad or cooked in various ways in different curries, fried, boiled or baked. It is also used in processed form. Eg. flakes, powder, paste and pickles. Onion ranks medium in calorific value, low in protein and very low in vitamins. It is also a good cleanser and healer. Onion juice is applied to burns and bites or stings. It is believed to be very useful to cure sores and ulcers. Onion is affected by various plant pathogens of which leaf blight was reported to be one of the major diseases in India (Gupta and Pandey, 1986; Tripathi et al., 2008; Ramjegathesh et al., 2011) and it causes yield loss 25 to 75% (Bisht and Agarwal, 1994). The disease causes extensive damage to bulbs as well as seed crop and average intensity was found in the range of 5 to 45% (Gupta et al., 1985). An understanding the role of environmental conditions and the survival of pathogen is necessary to develop management practices (Sallam, 2011). Therefore, objective of the study included isolation and identification of pathogen causing leaf blight disease of onion and the effect of mycelial growth on different media, pH, thermal death point and different carbon and nitrogen sources.
MATERIALS AND METHODS
Collection and isolation of the fungus: Leaves of onion (Allium cepa var. aggregatum) showing typical symptoms of dark blighted lesions caused by Alternaria alternata were collected and the fungus isolated by the following technique indicated below.
The infected leaf bits along with some healthy portions were cut into small bits and surface sterilized using 1:1000 mercuric chloride solutions for 30 sec. The bits were washed thoroughly in sterile distilled water for three times to remove traces of mercuric chloride. The molten warm PDA medium was poured in sterilized petri plates and allowed to solidify. The surface sterilized leaf bits were placed on PDA medium. These plates were incubated at room temperature 28±2°C and observed periodically for fungal growth. Apparently pure colonies were developed from PDA slants. The slants were incubated at 28±2°C for sporulation for 10-18 days. Then such slants containing apparently pure culture were used for further purification.
Total of ten isolates were isolated from different growing areas. The pathogen was identified up to species level based on their cultural and morphological characters. Then the identification was further confirmed from Indian type culture collection and identification, culture supply services, Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi.
Virulence (Pathogenicity) of the isolates: The pathogenicity of purified isolates of A. alternata was tested and it was proved by Koch's Postulates conducted on the Agri found onion red varieties (Elwakil et al., 2009; Jain et al., 2005). The plants were raised in mud pots under glass house conditions. Three replications were maintained in each isolate. The conidial suspension (5x105 spores mL-1) was in prepared sterile distilled water from 9-day-old PDA cultures of the different isolates of A. alternata. These spore suspensions were sprayed on the onion plants of 30-day-old plants. Water congestion was provided to the plants both 24 h prior and after inoculation by covering the plants with moist polythene bag. The inoculation was done on cool evening hours. The plants sprayed with sterile distilled water served as control. The inoculated plants were maintained in glass house condition. The severe symptoms were observed on 14 days after inoculation and the disease intensity was recorded. The symptoms were observed and compared with the original symptoms. The fungus was reisolated from artificially inoculated onion leaves and compared with original culture isolate.
Environmental factors affecting to the growth of isolates: The effect of different culture media, carbon, nitrogen sources, pH and thermal death point on the mycelial growth of A. Alternata in vitro agar plates were studied.
Morphological characters: The morphological characters viz., conidia shape, size (length and width), number of cells /conidia, colour of conidia and sporulation time (days) were observed on ten isolates. Spores of A. alternata were taken from the pure culture and mounted in a glass clean slide. Spores are mixed with lactophenol thoroughly in order to obtain a uniform spread over and covered with cover slip. The morphological characters were measured in 50 conidia under high power objective using ocular and stage micrometers.
Cultural characters
Growth characters on solid media: The composition and preparations of the above media (Synthetic media viz., Asthana and Hawker's medium 'A', Czapek (Dox) Agar medium, Glucose phosphate agar medium, King's B medium, Modified Czapek (Dox) Agar medium, Nutrient Agar medium, Richards' medium, Starch agar medium, Water agar medium and non synthetic media viz., Host Leaf Extract Agar medium, Carrot Extract Agar medium, Potato Dextrose Agar medium) were prepared as suggested by "Ainsworth and Bisby's Dictionary of the fungi" by Ainsworth (1961) and Plant Pathological methods, fungi and bacteria by Tuite (1969) except host leaf extract medium.
Twenty milliliter of the sterilized and melted (warm) medium was poured into the sterile petri plates and allowed to solidify. The actively growing 10-day-old culture 9 mm disc was placed on the centre of the petri plates. The plates were incubated at room temperature (28±2°C) for 10 days. Three replications were maintained. The diameter of the mycelial growth was recorded by visual observation.
Nutritional studies
Utilization of carbon sources: The carbon requirement of the fungus was studied in the Czapek (Dox) medium. The quantity of carbon compounds added was determined based on their molecular weight. Carbon sources used in the study were carboxymethyl cellulose, glucose, fructose, lactose, mannitol, maltose, sucrose and starch. The sterilized melted warm medium was poured in the sterilized petri plate and allowed to solidify. The actively growing 10-day-old culture of 9 mm disc was placed on the centre of the petri plate. The plates were incubated at room temperature (28±2°C) for 10 days. Three replications were maintained. The diameter of the mycelial growth was recorded.
Utilization of nitrogen sources: The nitrogen requirement of the fungus was studied in the Czapek (Dox) medium. The quantity of nitrogen compounds added was determined based on their molecular weights so as to provide an equivalent amount of nitrogen, as that of potassium nitrate present on the basal medium. Nitrogen sources used in the study were ammonium nitrate, ammonium molybdate, ammonium oxalate, ammonium sulphate, potassium nitrate, sodium nitrate, urea and thiourea. The sterilized melted warm medium was poured in the sterilized petri plates and allowed to solidify. The Czapek Dox medium without nitrogen sources served as control. The actively growing 10 days old culture of 9 mm disc was placed on the centre of the petri plate. The plates were incubated at room temperature (28±2°C) for 10 days. The diameter of the mycelial growth was recorded. Three replications were maintained.
Physiological studies
Effect of pH on the growth of A. alternata: The effect of pH on the growth of the pathogen was studied as per the method followed by Kiryu (1939) using PDA medium. Different pH levels viz., 4.0, 4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5 and 9.0 were used. The pH levels of the medium were adjusted in a Digital pH meter using 0.1 N Hydrochloric acid and 0.1 N Sodium hydroxide. The media of different pH levels were sterilized, cooled and poured in the sterilized petri plates in 20 mL quantities and allowed to solidify. The actively growing 10-day-old culture of 9 mm disc was placed on the centre of the petri plates. The plates were incubated at room temperature (28±2°C) for 10 days. The diameter of the mycelial growth was recorded. Three replications were maintained. Thermal death point: The effect of temperature on growth of the pathogen was studied using spore suspension (5x105 spores mL-1) of A. alternata. The spore suspension was prepared by using sterile distilled water. The test tubes containing 1 mL of spore suspension were exposed to heat at different temperatures viz., 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 and 50°C for 10 min in a thermostatic water bath. These spore suspensions in each of tubes were separately poured into sterilized petri plates under aseptic condition and fifteen mL of sterilized PDA medium was poured in each petri plate. These plates were gently rotated to get uniform suspension and allowed to solidify. The plates were incubated at room temperature (28±2°C) for 5 days. Three replications were maintained. The growth of the pathogen was recorded in each treatment.
RESULTS
Isolation and identification of the pathogen: The process of isolation resulted in ten isolates of pathogen collected from different regions. All the isolates were confirmed by morphological and cultural characters as isolates of A. alternata.
Pathogenicity test: Initially, twelve days after inoculation the symptoms first appeared in the form of small whitish flecks on the leaf tip. These increased in size and became sunken lesions. The lesions gradually exhibited a grayish tint at the centre surrounded by a yellow halo.
Morphological characters: In all the ten isolates, the conidia were muriform shape and light brown colour. The length of their conidia was varied from 30.99 to 42.47 μm .The longest conidia was in I4 from Paavur Chathriam (42.47 μm) followed by I3 from Periasamy Puram (40.33 μm) and I8 from Kamanayakkanpatti (40.13 μm). The conidium was the shortest in I6 from Aruppukkottai (30.99 μm). The width of the conidia was varied from 11.90 to 17.37 μm. The I4 registered the maximum conidial width (17.37 μm) followed by I5 (15.54 μm) and I8 (14.83 μm), while the I2 were the narrow conidia (11.90 μm). Generally, all the isolates produced both beaked and unbeaked conidia. The beak length of conidia varied from 18.7 to 23.81 μm. The I4 had the longest beak (23.81 μm) followed by I3 (22.80 μm) and I5 (22.10 μm), while it was short in I2 (4.2 μm) I10 (8.0 μm). The spores of isolates I4, I5, I8 and I9 have been identified as bigger, I1, I2, I3, I4 and I10 as medium and I6 as small. The number of cells in each conidium varied from 2-9. Among all the isolates, the conidia of I1, I4 I5 and I6 had 2-9 cells respectively when I3 and I10 had 3-7, I2 and I7 possessed in 3-8 cells per conidium. All the isolates took 13-16 days for sporulation, while isolate I4 sporulated in 13 days. I5 took on 14 days to sporulate. The isolates I2, I3, I6 and I9 sporulated with in 15 days, while I1, I7, I8 and I10 required 16 days for sporulation (Table 1).
| Table 1: | Morphological characters of conidia of A. alternata isolates |
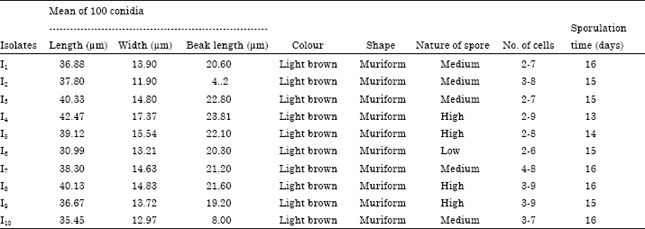 | |
Cultural characters
Effect on growth of A. alternata on different solid media: All the twelve media supported differently the growth of all the ten isolates of A. alternata (Fig. 1). The diameter of mean mycelial growth of the pathogen was maximum in host leaf extract agar medium (9.00 cm) followed by modified Czapek (Dox) agar (9.00 cm), potato dextrose agar (8.14 cm) and carrot agar medium (7.94 cm), while it was very less in water agar medium (0.92 cm) and less in glucose phosphate agar (5.14 cm). The isolate I4 from Paavur Chathiram had maximum mean mycelial growth (6.92 cm) on all the twelve media followed by I5 from Kulippatti (6.89 cm) and I7 from R. Combai (6.81 cm). The minimum mean mycelial growth was recorded in I8 of Akkanayakkanpatti (6.58 cm).
The mycelial growth of the ten isolates differed significantly on different growth media, The mycelial growth was maximum in I4 on host leaf agar medium, modified Czapek's Dox agar, potato dextrose agar, nutrient agar and carrot agar medium were 9.00, 9.00, 8.36, 8.12 and 8.05 cm respectively, while the growth of the pathogen less in water agar (1.17 cm) and glucose phosphate agar (5.27 cm).
Nutritional studies
Effect on growth of A. alternata isolates on different carbon sources: All the eight carbon sources supported differently on the growth of all the ten isolates of A. alternata (Table 2). The maximum mean mycelial growth was recorded in maltose (8.82 cm) followed by glucose (8.67 cm), sucrose (8.29 cm) and fructose (7.83 cm), while the carboxymethyl cellulose had the minimum mycelial growth (7.39 cm) compared to control respectively. Both isolates I4, I5 recorded highest mean mycelial growth of 8.00 cm followed by I7 (7.98 cm) and I9 (7.94 cm). I8 had a minimum mean mycelial growth of 7.72 cm. The following carbon sources maltose, glucose and sucrose recorded the maximum mycelial growth of 9.00, 8.88 and 8.51 cm respectively. All are significantly different from each other.
Effect on growth of A. alternata isolates on different nitrogen sources: Eight nitrogen sources were tested to assess their growth of the pathogen of all the ten isolates of A. alternata (Table 3). Potassium nitrate produced maximum mean mycelial growth (9.00 cm) followed by sodium nitrate (7.32 cm) and ammonium molybdate (6.18 cm), while thiourea had minimum mean mycelial growth (2.63 cm).
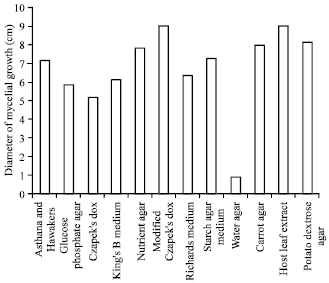 | |
| Fig. 1: | Effect of different solid media on the growth of Alternaria alternata |
| Table 2: | Effect of different solid media on the growth of A. alternata isolates (in vitro) |
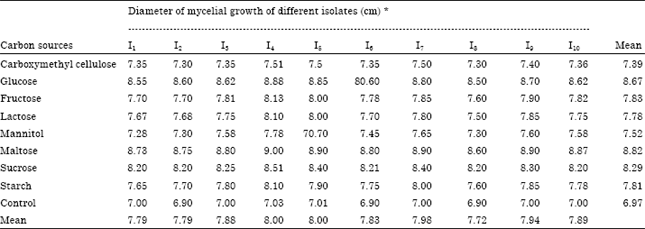 | |
| CD (p = 0.05). Media: 0.02148. Isolates: 0.01961. Mediaxisolates: 0.06793 * Mean of three replications | |
| Table 3: | Effect of different Nitrogen sources on the growth of A. alternata isolates (in vitro) |
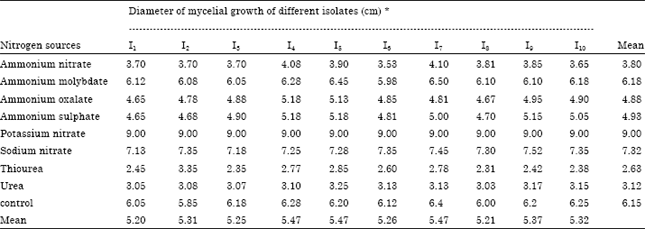 | |
| CD (p = 0.05). Nitrogen sources: 0.1003. Isolates: 0.10576. Nitrogen sources x Isolates: 0.31727. * Mean of three replications | |
Isolates I4, I5 and I7 had recorded the same level of mycelial growth (5.47 cm) respectively. The least value was recorded in isolate I1 (5.2 cm).
Physiological studies
Effect on growth of A. alternata isolates at different pH levels: All of the eleven pH levels, pH 4.5 was favoured to be ideal and produced the maximum mean mycelial growth (8.87 cm) followed by pH 4.0 (8.66 cm) and pH 5.0 (8.46 cm). The mean mycelial growth was minimum in pH 9.0 (6.10 cm). The isolate I4 registered maximum mycelial growth in pH 4.5 (8.97 cm) followed by I5 and I6, which registered 8.92, 8.91 cm respectively (Table 4).
Thermal death point of A. alternata isolates: The conidia of all the ten isolates of A. alternata germinated up to a temperature of 46°C. The conidia of the isolates I4, I5 and I7 had germinated at 48°C, but they failed to germinate at 49°C indicates as a thermal death point for their isolates, while the rest had their at 48°C and 47°C (Table 5).
| Table 4: | Effect of different pH on the growth of A. alternata isolates (in vitro) |
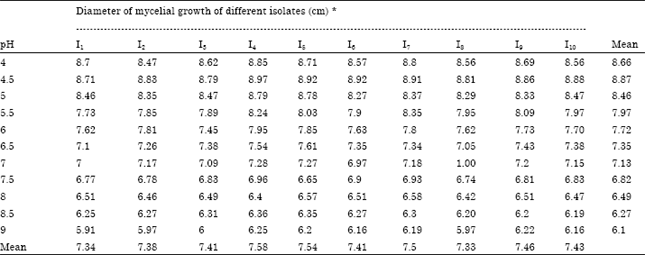 | |
| CD (p = 0.05). pH: 0.04583. Isolates: 0.04370. pHxIsolates: 0.14494. * Mean of three replications | |
| Table 5: | Thermal death point of A. alternata isolates (in vitro) |
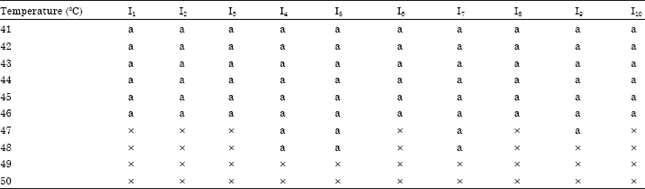 | |
| a: Present; x: Absent | |
DISCUSSION
Morphological characters are important tools in identification and classification of the fungus. In the present study, spore size, septation of conidiophore and conidia and shape of conidia were used for identifying the fungus. The characteristics of the fungus in question were compared with Alternaria species reported on the host and characters agreed very closely with those of A. alternata (Fr.) Keissler with minor variation in shape dimension which may be either, due to host or environmental factor and hence, were considered to fall with in the limits for species (Abubakar and Ado, 2009).
All the ten isolates produced light brown and muriform shape conidia measuring about 30.99-42.47 μmx11.9-17.37 μm. The beak length of the isolates varied from 18.7 μm to 23.81 μm. The number of cells varied from 2-9 among the isolates. The character agreed with the original descriptions given by Alternaria alternata. However, Nolla (1927) reported the length of conidia (105-320x12-24 μm) in Alternaria alli, 105-220-17.5-26 μm in A. porri (Nolla, 1927), 58.8-184.8x21-46.2 μm in Alternaria cepulicola (Rao, 1974), 38-146x10-73 μm in Alternaria cepulae (Ponnappa, 1974) and 10.26-77.52x4.56-14.82 μm in Alternaria alternata (Keissler). Neergard (1945) reported 1-9 cells in Alternaria alternata. Muthulakhsmi (1990) and Cuervo-Parra et al. (2011) reported A. alternata produced both beaked and unbeaked conidia. Mohan (1996) also reported such variations among the isolates in number of cells of conidia ranging from 2-9 in A. palandui. Sumathi (1997) reported that the isolates of A. sesami also had beaked and unbeaked conidia. Karthikeyan (1999) also reported that both beaked and unbeaked conidia were observed on ten out of seven isolates of A. palandui. Pandey and Vishwakarma (1999) reported that 4-6 septation observed in A. alternata.
Fungi secure food and energy from the substrate upon which they live in nature. In order to culture fungus in the laboratory, it is necessary to furnish essential elements and compounds in the medium for their growth and other life processes. All media are not equally good for all fungi, nor there can a universal substrates or artificial media upon which all fungi grow. So, different media including both synthetic and non synthetic media were tried for A. alternata in the present investigation. The isolates exhibited variations of growth in different solid media tested. The growth was highest in modified Czapek (Dox) agar medium (9.0 cm), host leaf extract agar medium (9.0 cm) followed by potato dextrose agar (8.24 cm), while the lowest growth was recorded in water agar medium (0.92 cm).
Potato dextrose agar was the best medium for the growth of A. carthami inciting leaf blight of safflower (Awadhuya, 1991), A. solani causing leaf blight of tomato (Babu, 1994), A. alternata causing leaf blight of tobacco and jasmine (Li, 1994; Kamalalakshmi, 1996), A. palandui causing leaf blight of onion (Mohan, 1996; Karthikeyan, 1999). Oat agar medium supported maximum growth of A. palandui and A. alternata inciting leaf blight disease of onion (Ahmed, 1960; Kannan, 1992). Czapek's medium was the best for the growth of A. porri causing leaf blight of onion (Gupta et al., 1987), A. alternata causing leaf blight of brinjal (Pandey and Vishwakarma, 1998), Chilli (Anand and Bhaskaran, 2009) and Noni (Hubballi et al., 2010). Among the ten isolates I4 from Paavur Chathiram recorded significantly the highest mycelial growth, while isolate I8 from Akkanayakkanpatti recorded the minimum mycelial growth. Maximum mycelial growth was recorded in virulent isolates of A. alternata (Kannan, 1992), A. sesami (Sumathi, 1997), A. palandui by Mohan (1996) and Karthikeyan (1999).
Carbon occupies a unique position among the essential elements required by the living organisms. Carbon utilization is speculated to be dependent upon enzyme system. The utilization of various carbon compounds may depend either on the activity of the fungus to utilize certain simpler forms or on its power to convert the complex carbon compounds into simpler forms which may be easily utilized. As a component of both structural and functional constituents carbon comprises about 50% of the total mycelial dry weight in fungi (Bilgrami and Verma, 1978). Chaturvedi (1966) reported that sucrose, lactose, maltose and raffinose supported the maximum growth of A. alternata. Among the various carbon compounds tested, maltose supported the maximum growth (8.82 cm) of the fungus followed by glucose (8.67 cm) and sucrose (8.27 cm). Padmanabhan and Narayanaswamy (1977) have reported different species of Alternaria which utilized the sucrose most efficiently as a carbon source. Bilgrami and Verma (1978) opined the sucrose, being the major sugar component of photosynthetic plants, is generally utilized as a good carbon source by most of the plant pathogenic fungi.
The fungi for their structural and functional processes use nitrogen, like carbon. Nitrogen is very important element in the protein synthesis. But all the sources of nitrogen are not equally good for the growth of all fungi (Lilly and Barnett, 1951). In the present study, among the various nitrogen sources tested, potassium nitrate supported the maximum growth (9.0 cm) of the fungus followed by sodium nitrate (7.32 cm). Isolate I4 showed the maximum mycelial growth of 5.47 cm. In general, inorganic nitrogen supported better growth of the fungus than the organic nitrogen sources. Mallikarjunaiah and Rao (1972) found that A. tenuissima made a good growth in potassium nitrate and sodium nitrate. Patil (1987) observed that potassium nitrate and L-arginine were the best inorganic and organic nitrogen sources for the growth of A. tenuissima. In the present study, least growth was observed in thiourea. Inferiority of ammonia salts as source of nitrogen by some fungi has been discussed by Cochrane (1958). According to him, urea break down into ammonia on autoclaving and ammonia in high concentration is toxic to fungi. Ammonia may be accumulated to an inhibitory level and contribute to the so called staling effect. In the present study also, least growth on the thiourea might have occurred, because of breakdown of urea to ammonia which has inhibited the growth of the fungus (Mishra et al., 2011).
In the present study the maximum mycelial growth of A. alternata was recorded in pH 4.5 (8.47 cm) followed by pH 4.0 (6.67 cm), while minimum was recorded at higher levels of pH 9.0. Ahmed (1960) reported that good growth of A. palandui was observed at pH levels 3.0, 4.0 and 5.0. Saeed et al. (1995) observed that isolates of A. alternata grew best at pH 5.5. The isolates of A. palandui recorded the maximum mycelial growth at pH 4.5 followed by pH 4.0 and the growth was retarded at higher pH levels (Mohan, 1996; Karthikeyan, 1999).
Temperature differs almost every function of the fungi and each fungi has its temperature range for growth and sporulation .The thermal death point of the isolates of A. alternata differed with a narrow range of 47-49°C. Mohan (1996) and Karthikeyan (1999) observed that the conidia of all A. palandui isolates germinated up to 46°C.
CONCLUSION
The pathogen identification, morphological, cultural and environmental effects are very important for the production of conidia. A. alternata produced light brown conidia with muriform shape. Host leaf extract agar and modified Czapek Dox medium favored for the growth of mycelium. In nutritional requirements maltose and potassium nitrate amended medium was highly suitable for the growth A. alternata, while pH 4.5 was found to be an ideal growth of A. alternata.
ACKNOWLEDGMENT
The authors wish to express their appreciation to Tamil Nadu Agricultural University, Coimbatore, India for the providing facility of this doing work.
REFERENCES
- Abubakar, L. and S.G. Ado, 2009. Genotypex environment interaction for resistance to purple blotch (Alternaria porri L. (ellis) Cif.) in onion (Allium cepa L.) in Nigeria. Asian J. Crop Sci., 1: 15-25.
CrossRefDirect Link - Anand, T. and R. Bhaskaran, 2009. Exploitation of plant products and bioagents for ecofriendly management of chilli fruit rot disease. J. Plant Protect. Res., 49: 195-203.
Direct Link - Chaturvedi, C., 1966. Utilization of oligosaccharides by three imperfect fungi. Mycopathologia, 29: 323-330.
CrossRefDirect Link - Cuervo-Parra, J.A., V. Sanchez-Lopez, M. Ramirez-Suero and M. Ramirez-Lepe, 2011. Morphological and molecular characterization of Moniliophthora roreri causal agent of frosty pod rot of cocoa tree in tabasco, Mexico. Plant Pathol. J., 10: 122-127.
CrossRefDirect Link - Elwakil, M.A., I.M. El-Refai, O.A. Awadallah, M.A. El-Metwally and M.S. Mohammed, 2009. Seed-borne pathogens of faba bean in Egypt: Detection and pathogencity. Plant Pathol. J., 8: 90-97.
CrossRefDirect Link - Hubballi, M., S. Nakkeeran, T. Raguchander, T. Anand and R. Samiyappan, 2010. Effect of environmental conditions on growth of Alternaria alternate causing leaf blight of noni. World J. Agric. Sci., 6: 171-177.
Direct Link - Mishra, V.K., S.K. Biswas and M. Rajik, 2011. Biochemical mechanism of resistance to alternaria blight by different varieties of wheat. Int. J. Plant Pathol., 2: 72-80.
CrossRefDirect Link - Sallam, N.M.A., 2011. Control of tomato early blight disease by certain aqueous plant extracts. Plant Pathol. J., 10: 187-191.
CrossRefDirect Link - Padmanabhan, P. and P. Narayanaswamy, 1977. Growth studies of Alternaria macrospora Zimm. an incitant of leaf spot disease of cotton. Madras Agric. J., 64: 258-261.
Direct Link - Ponnappa, K.M., 1974. Leaf blight of onions (Allium cepa L.) caused by Alternaria cepulae Ponnappa and Deshpande sp. nov. 1. Comparative morphology and physiology. Beihfte Zur Nova Hedurigia, 47: 547-564.
Direct Link - Ramjegathesh, R., E.G. Ebenezar and M. Muthusamy, 2011. Management of onion leaf blight by Alternaria alternata (FR.) keissler by botanicals and bio-control agents. Plant Pathol. J., 10: 192-196.
CrossRefDirect Link - Jain, S., K.S. Verma and M.D. Shah, 2005. Pathological studies on Alternaria alternata (Fr.) Keiss. Causing leaf blight of pear. Plant Pathol. J., 4: 51-53.
CrossRefDirect Link - Tripathi, M.K., S. Tiwari and U.K. Khare, 2008. In vitro selection for resistance against purple blotch disease of onion (Allium cepa L.) caused by Alternaria porri. Biotechnology, 7: 80-86.
CrossRefDirect Link
dugasa hinsermu Reply
morphology of onion leaf blight