
Salma Sai Kachout
National Institute of Agronomic Research of Tunisia (INRAT), University Tunis Carthage, Tunisia
LiveDNA: 216.24597
Saleh Benyoussef
National Institute of Agronomic Research of Tunisia (INRAT), University Tunis Carthage, Tunisia
Aziza Zoghlami
National Institute of Agronomic Research of Tunisia (INRAT), University Tunis Carthage, Tunisia
Mohamed Chakroun
National Institute of Agronomic Research of Tunisia (INRAT), University Tunis Carthage, Tunisia
International Journal of Plant Breeding and Genetics
Year: 2019 | Volume: 13 | Issue: 1 | Page No.: 12-18







ABSTRACT
Background and Objective: Grass pea (Lathyrus sativus L.) is a multipurpose grain legume with an indeterminate growth habit. The objective of this study was to identify adaptation mechanisms of grass pea to water deficits. Materials and Methods: In the germination experiment, the used osmotic potential were: 0 (distilled water); -0.03, -0.1, -0.7 and -1.0 MPa, osmotic shock induced by increasing concentrations of polyethylene glycol (PEG). After treatment for 7 days, germination percentage, root and shoot length, seedling fresh and dry weight were measured. In the second experiment, water deficit was imposed on plants of grass pea by withholding water from first flowering. Results: Seeds of L. sativus were able to germinate at all concentrations of PEG treatments. Higher concentrations of PEG (-1.0 MPa) were less inhibitor effect on seedling shoot and root elongation. This study showed that under water deficit, grass pea avoids dehydration through a reduction in leaf area and fresh biomass. Water deficit reduced dry matter, seed yield, seed number, seed size, flowers number and harvest index, respectively, when compared with the controls. Conclusion: Conclusively, grass pea showed a better germination under osmotic stress; therefore, it seems to be a suitable candidate for breeding programs.
PDF Abstract XML References Citation
How to cite this article
Salma Sai Kachout, Saleh Benyoussef, Aziza Zoghlami and Mohamed Chakroun, 2019. Effect of Water Deficit during Germination and Flowering Period of Grass Pea (Lathyrus sativus L.). International Journal of Plant Breeding and Genetics, 13: 12-18.
DOI: 10.3923/ijpbg.2019.12.18
URL: https://scialert.net/abstract/?doi=ijpbg.2019.12.18
DOI: 10.3923/ijpbg.2019.12.18
URL: https://scialert.net/abstract/?doi=ijpbg.2019.12.18
INTRODUCTION
The environmental stress such as drought is serious obstacle for horticulture and field crops in all around the world, especially in arid and semiarid regions. Drought is considered as a worldwide problem currently which is expected to elaborate in the near future1. Environments with water restriction and salt excess produce harmful effects on seeds, such as a significant reduction in the germination percentage and rate in the root length and fresh mass and in the shoot of seedlings2; they may even cause a complete inhibition of the germination process of seeds3. In plant production, the most important factors limiting crop productivity are environmental stresses. Disruption of plant water management caused by drought, salinity or low temperature is a major yield decreasing factor4. The use of plants that are more resistant to environmental stresses, particularly in the germination and early seedling growth stage is gaining importance day by day5. Several Lathyrus species and in particular Lathyrus sativus (grass pea) have great agronomic potential as grain and forage legume, especially in drought conditions6. It can survive harsh environmental conditions, making the crop suitable for drought prone areas7. It is superior in yield, protein value, nitrogen fixation and drought, flood as well as salinity tolerance than other legume crops. Grass pea not only has potential as an agriculturally important crop for animal feed and human food, it can also be useful for studies of plant drought resistance8.
In Tunisia, the genus Lathyrus is cultivated mainly for grain production (food) and also used as fodder (animal feed) by farmers. Grass pea (Lathyrus sativus L.) is a legume crop with a unique adaptation features to abiotic stresses such as drought or poor soil conditions6 and as a result is considered a promising source of genes of interest. It is one of the relatively infrequent grain legumes. Also, high protein content makes this species interesting as a forage crop9-11. Although rich in protein, the utilization of grass pea grain is limited by the presence of water soluble, non-protein amino acid a-N-oxalyl diaminopropionic acid (a-ODAP) which causes lathyrism in animal12.
While grass pea crops are known to be well adapted to drought but details of growth and yield responses to water deficit during germination and the reproductive period are not fully understood. The objective of this study was to determine the effect of water stress on germination and during reproductive period as well as on some morpho-physiological parameters of Lathyrus sativus due to osmotic effect and to optimize the best priming treatment for these stress conditions.
MATERIALS AND METHODS
Design of simulated drought conditions: Water stress conditions were simulated by polyethylene glycol-6000 (PEG) at one of four concentrations. Table 1 shows how osmotic potential decreases with increasing PEG-6000 concentration:
OP = (-1.18×10–2)–C-(1.18×10–4)×C+(2.67×10–4)× C×T+(8.39×10–7)×C2T
Where:| C | = | PEG concentration |
| T | = | Temperature |
Plant materials and growing conditions: In the first experiment, to determine the tolerance of the germination to the water stress, the seeds of grass pea were imbibed in the distilled water and in solutions of Polyethylene Glycol (PEG-6000). For each experiment, 20 seeds were soaked in PEG 6000 solution of different water potentials (-1.0, -0.7, -0.1 and -0.03 MPa) were prepared. The number of seeds germinated was counted daily for 7 days. Distilled water was used as a control (0 MPa). The PEG solutions were prepared dissolving different concentrations of PEG 6000 in deionized water. Seeds were germinated at a constant temperature of 25±1°C, in a controlled chamber. Samples of 80 seeds (4 replicates of 20 seeds each) were placed in Petri dishes containing a single filter paper moistened with 10 mL of PEG solution. After treatment for 7 days, all grass pea seedlings were sampled. Germination percentage, root and shoot length, water content, seedling fresh and dry weight and content of sugar were measured in the study.
In the second experiment, growth conditions and stress treatments of grass pea (Lathyrus sativus L.) were conducted in a greenhouse at the Experimental Station of INRAT in Tunisia (35°87 N, 9°96 E) in January, 2018. When plants reached flowering, drought was imposed to stress pots by withholding water for a week, while non-stressed plots continued receiving irrigation. The experiment design was split-plot with 6 replications.
Measurement of germination and agronomic parameters
Experiment 1: Germination: The seedling fresh and dry weight expressed as mg/plant and length of root and shoot (mm) were determined after 7th day of germination test in all the germinated seedlings.
| Table 1: | Amounts of polyethylene glycol used for producing of different levels of osmotic stresses |
| *According to Michel and Kaufmann13 at temperature of 25°C | |
The seedling dry weight was determined after drying at 80°C for 24 h.
The germination process was evaluated during 7 days; germinated seeds were counted every day (February, 2018):
| • | The final Germination Percentage (GP) was calculated in the 7th day by using the following equation: |
![]()
| • | Fresh weight of seedling after 24, 42, 48 and 92 h at different osmotic potentials |
| • | Dry weight of hypocotyl and radicle |
| • | Radicle length and hypocotyl length of seedling |
| • | Water content (WC) at different osmotic potentials, the WC percentage was calculated by using the equation14: |
![]()
| • | Determinations of total sugars: Total sugars were determined by phenol-sulfuric acid reaction, the analysis consisted in addiction of 2.5 mL of sulfuric acid and 0.5 mL 5% (w v-1) phenol solution over 0.5 mL of diluted sample. Afterwards, it was kept at room temperature for 20 min. After the reaction time, the optical density was determined at 480 nm in spectrophotometer15 |
Experiment 2: Flowering period: In the second experiment, observations on yield and yield attributing characters: Plant height (cm), number of pods per plant, number of seeds per pod, number of seeds per plant, fresh weight of biomass per plant (g), dry weight of biomass per plant (g), number of flowers per plant were recorded in May, 2018.
Statistical analysis: The data was statistically tested by a one-way ANOVA followed by a Tukey’s test. The analysis of the significance of difference for each parameter was performed on the basis of mean values and a level of significance at p<0.05. Each parameter was measured in at least five reiterations in control, PEG treatment and water deficit groups.
RESULTS
Germination of grass pea at different osmotic potentials of PEG
Seed germination: The result showed that germination percentage was not significantly decreased by PEG solutions.
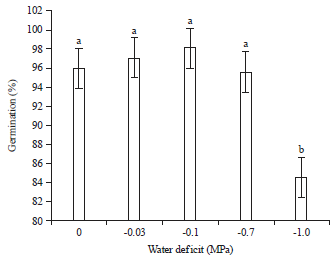 | |
| Fig. 1: | Germination percentages of grass pea at different osmotic potentials of PEG |
Values are Mean±SE (n = 5), one-way ANOVA was used to test differences among treatments, different letters indicate significant differences among concentrations of PEG 6000 at p<0.05 (Tukey’s test), control treatment was distilled water | |
Contrary, it increased considerably with decrease of osmotic potential. The maximum of germination was obtained after 3 days in all treatments. Decreasing osmotic potential from -0.7 to -1.0 MPa in the media caused a significant reduction in germination percentage. Seed germination was 96% under control and 84% under stress conditions (Fig. 1).
Seedling fresh weight: Decreasing water potential by PEG concentration caused a reduction in seedling fresh weight. Differences determined among the treatments were significant. When early seedlings were exposed 24 h from water stress to optimal conditions (-1.0 MPa), they were accompanied by a decrease in fresh weight of about 0.12 mg (from 0.36-0.24 mg) (Fig. 2).
Seedling dry weight: Dry weights of hypocotyl and radicle of L. sativus grown in different osmotic potential are presented in Fig. 3. Generally, drought conditions generated by PEG significantly influenced dry weight of radicle and hypocotyl of grass pea. The highest and lowest dry weight were observed at control treatment and at -1.0 MPa (Fig. 3). However, at the different osmotic potential the dry weight deceased significantly with increasing drought levels (p<0.05) compared to the control (Fig. 3).
Water content: The water content of Lathyrus shoots decreased under osmotic stress (Fig. 4). A significant reduction in water content of grass pea was observed at lower osmotic potential (-1.0 MPa), as far as in the experiments with different osmotic potentials of PEG (Fig. 4).
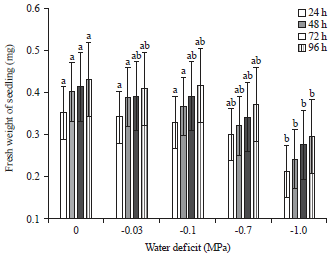 | |
| Fig. 2: | Seedling fresh weight of field pea at different osmotic potentials of PEG |
Values are Mean±SE (n = 5), one-way ANOVA was used to test differences among treatments, different letters indicate significant differences among concentrations of PEG 6000 at p<0.05 (Tukey’s test), control treatment was distilled water | |
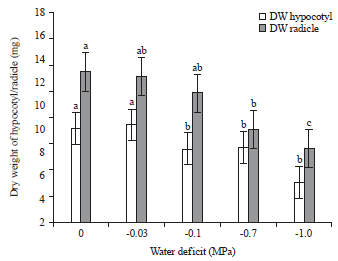 | |
| Fig. 3: | Dry weight of radicle and hypocotyl of grass pea at different osmotic potentials of PEG |
Values are Mean±SE (n = 5), one-way ANOVA was used to test differences among treatments, different letters indicate significant differences among concentrations of PEG 6000 at p<0.05 (Tukey’s test), control treatment was distilled water | |
Length of hypocotyl and radical: For the length of hypocotyl and radicle of grass pea at different osmotic potentials, the solutions of -0.7 and -1.0 MPa had a stronger negative effect than the -0.03 and -0.1 MPa solutions (Fig. 5). The highest radicle was recorded in control (7.3 cm), whereas the lowest one was noted at -1.0 MPa (2.9 cm) (Fig. 5).
Content of sugar: The results show that, along with a decrease in osmotic potential, the accumulation of sugar increased significantly in both the radicle and the hypocotyl of L. sativus (Fig. 6).
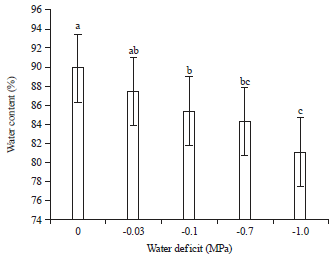 | |
| Fig. 4: | Water content of grass pea at different osmotic potentials of PEG |
Values are Mean±SE (n = 5), one-way ANOVA was used to test differences among treatments, different letters indicate significant differences among concentrations of PEG 6000 at p<0.05 (Tukey’s test), control treatment was distilled water | |
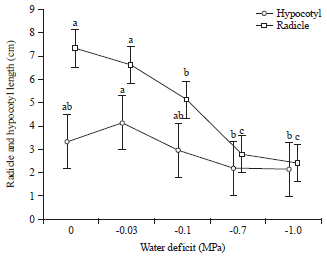 | |
| Fig. 5: | Length of hypocotyl and radicle of grass pea at different osmotic potentials of PEG |
Values are Mean±SE (n = 5), one-way ANOVA was used to test differences among treatments, different letters indicate significant differences among concentrations of PEG | |
The contents of sugar increased with decreasing of osmotic potential with that in the radicle being significantly higher than that in the hypocotyls. The maximum content of sugar in hypocotyl of grass pea seedling was 5.2 μmol g–1 DW observed at -1.0 MPa compared to the minimum 3.5 μmol g–1 DW at control. Decreasing of osmotic potential affected the content of sugar in radicle, the maximum was 15.2 μmol g–1 DW at -0.7 MPa treatment and the lowest was 5.8 μmol g–1 DW at control (Fig. 6).
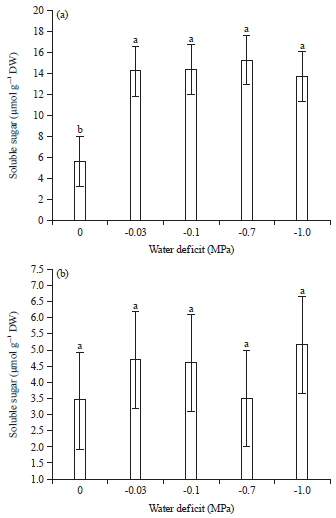 | |
| Fig. 6(a-b): | Effects of PEG induced water deficit on the content of sugar in (a) Radicle and (b) Hypocotyl of grass pea seedling |
Values are the means of 5 replicates means followed by different letters are significantly different at p<0.05 according to Duncan’s method | |
| Table 2: | Effect of water deficit on yield, harvest index and yield components in Lathyrus sativus |
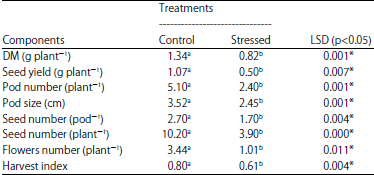 | |
| *Significant at 5%, values are means (n = 8) | |
Effect of drought during flowering stage: Generally, drought conditions generated by withholding water significantly influenced the vigor of grass pea plants. However, the water deficit caused a significant decrease in plant height, number of branches and leaf area of Lathyrus sativus (Fig. 7). The highest plant was obtained at control with 66 cm compared to stressed plant with 45 cm.
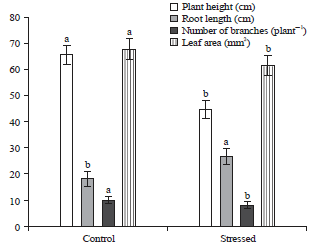 | |
| Fig. 7: | Variation of plant height, root length, number of branches and leaf area of Lathyrus sativus with the water deficit |
| Different letters indicate significant difference among levels of nitrogen treatment at p<0.05 | |
Contrary, application of water deficit stress during the flowering stage increased sharply root length of grass pea plants about 28 cm compared to their respective controls with 18 cm. The main effect of drought was significant for number of branches and leaf area by reduction of the mentioned traits (Fig. 7). Water deficit decreased flower production by 70 % compared with the control (Table 2). Under stress conditions, the harvest index was 0.61 instead of 0.80 in the control treatment.
DISCUSSION
In this study, all determined germination and seedling parameters (fresh and dry weight, radical length, hypocotyl length and water content) decreased significantly in response to water stress with PEG of grass pea. Seed germination and early seedling growth were potentially the most critical stages for water stress16. The results showed clearly that the pea seed germination and seedling growth were inhibited by osmotic effect during the seedling development. Contrary, Piwowarczyk et al.17 found that osmotic stress had no influence on seed germination of any of the accessions of Lathyrus tested. The depressive effects of an increased water deficit upon germination percentage and following seedling emergence have been extensively shown in many crops18-20. Jiang et al.21 showed that grass pea compared to the common garden pea has better adaptation to drought stress. This is a common behavior of plants and in fact, these early phases of the life cycle are generally the most sensitive to stress19,22,23.
Concentrations of total soluble sugars increased sharply in PEG-treated grass pea seedling compared to their respective controls, which was consistent with some plants under water stress such as maize (Zea mays L.)24 and wheat (Triticum aestivum L.)25. The significant reduction of pod number and the growth inhibition of L. sativus caused by water deficit was due to interference with plant growth processes like cell division and cell enlargement, inhibition in nutrient uptake, reduction in dry matter production due to inhibition of metabolic processes such as photosynthesis and respiration26. The sensitivity of flowers and pods to water deficit varies between species and genotypes as well as growth environment27. Water deficit reduced flower production in an indeterminate soybean by reducing node numbers28,29. Mirzaei et al.30 reported that whatever drought stress is closer to the pod formation stage, its effect will be higher on the number of pods and results showed that part of the yield loss in stress conditions is related to the reduction of number of seeds per pods and number pods. Usually, when water deficit stress is applied after the flowering stage, it causes the reduction of pod number per plant by shortening the flowering period, the reproductive growth duration and finally their fertility of some flowers and their abscission31. Pandey et al.32 showed that during water deficit stress at flowering stage, the seed yield decreases due to reducing seeds weights. Moreover, Kage et al.33 demonstrated that dry matter partitioning and biomass disposition are strongly connected with plant productivity under drought stress conditions. Decrease of dry matter was noted in many plant species under drought conditions19,34. For this reason, L. sativus is gaining interest as grain legume crops in Mediterranean-type environments. However, grass pea as highly resistant crops may serve as potential sources of new and useful genetic traits, such as resistance genes.
CONCLUSION
Grass pea plants were found to be more susceptible during flowering as compared to drought stress at germination. Water deficit caused by PEG treatment resulted in significantly decreased dry weight of radicle and hypocotyl, water content and length of hypocotyl and radicle of grass pea. The yield component’s was also affected by drought stress as dry matter and number of pod and seed per plant declined. However, basic research on the mechanisms of drought resistance by grass pea is essential to understand how this plant combats water deficit stress.
SIGNIFICANCE STATEMENT
Despite the importance of legumes for sustainable production of plant proteins, the progress achieved in Tunisia remains modest. Considering the drought tolerance and resistance to biotic stress of grass pea are valuable traits that can be beneficial and become an added commercial value to many other crops. These results indicate that tolerance of L. sativus under water deficit could be used as an adaptive trait for improving drought-resistant crops.
REFERENCES
- Dai, A., 2013. Increasing drought under global warming in observations and models. Nature Climate Change, 3: 52-58.
CrossRefDirect Link - Yan, M., 2015. Seed priming stimulate germination and early seedling growth of Chinese cabbage under drought stress. S. Afr. J. Bot., 99: 88-92.
CrossRefDirect Link - Attia, H., N. Karray and M. Lachaal, 2009. Light interacts with salt stress in regulating superoxide dismutase gene expression in Arabidopsis. Plant Sci., 177: 161-167.
CrossRefDirect Link - Jaleel, C.A., P. Manivannan, A. Wahid, M. Farooq, H.J. Al-Juburi, R. Somasundaram and R. Panneerselvam, 2009. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol., 11: 100-105.
Direct Link - Fallahi, H.R., P. Rezvani-Moghaddam, M.B. Amiri, M. Aghhavani-Shajari and R.Y. Biuki, 2015. The study of nutritional management of mother plant and seed priming by biofertilizers on improve salinity tolerance of wheat (Triticum aestivum L.) cv. Sayonz at germination period. J. Agroecol., 6: 689-700.
Direct Link - Vaz Patto, M.C., B. Skiba, E.C.K. Pang, S.J. Ochatt, F. Lambein and D. Rubiales, 2006. Lathyrus improvement for resistance against biotic and abiotic stresses: From classical breeding to marker assisted selection. Euphytica, Vol. 147.
CrossRefDirect Link - Massawe, F., S. Mayes and A. Cheng, 2016. Crop diversity: An unexploited treasure trove for food security. Trends Plant Sci., 21: 365-368.
CrossRefDirect Link - Choudhary, A., P. Pandey and M. Senthil-Kumar, 2016. Tailored Responses to Simultaneous Drought Stress and Pathogen Infection in Plants. In: Drought Stress Tolerance in Plants, Hossain, M.A., S.H. Wani, S. Bhattacharjee, D.J. Burritt and L.S.P. Tran (Eds.)., Vol. 1, Springer International Publishing, Germany, pp: 427-438.
- Polignano, G., 2007. Conservation, analysis of genetic diversity and maintenance of grasspea and other species (Lathyrus spp.) in Italy. Plant Genet. Resour. Newslett. Bull. Ressour. Phytogenet. Noticiario Recur. Fitogenet., 152: 73-76.
Direct Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Wu, Q.S. and R.X. Xia, 2006. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J. Plant Physiol., 163: 417-425.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Ahmad, S., F. Anwar, A.I. Hussain, M. Ashraf and A.R. Awan, 2007. Does soil salinity affect yield and composition of cottonseed oil? J. Am. Oil Chem. Soc., 8: 845-851.
CrossRefDirect Link - Piwowarczyk, B., I. Kaminska and W. Rybinski, 2014. Influence of PEG generated osmotic stress on shoot regeneration and some biochemical parameters in Lathyrus culture. Czech J. Genet. Plant Breed., 50: 77-83.
Direct Link - Murillo-Amador, B., R. Lopez-Aguilar, C. Kaya, J. Larrinaga-Mayoral and A. Flores-Hernandez, 2002. Comparative effects of NaCl and polyethylene glycol on germination, emergence and seedling growth of cowpea. J. Agron. Crop Sci., 188: 235-247.
CrossRefDirect Link - Okcu, G., M.D. Kaya and M. Atak, 2005. Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. For., 20: 237-242.
Direct Link - Kaya, M.D., G. Okcu, M. Atak, Y. Cikili and O. Kolsarici, 2006. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron., 24: 291-295.
CrossRefDirect Link - Jiang, J., M. Su, Y. Chen, N. Gao and C. Jiao et al., 2013. Correlation of drought resistance in grass pea (Lathyrus sativus) with reactive oxygen species scavenging and osmotic adjustment. Biologia, 68: 231-240.
CrossRefDirect Link - Vicente, O., M. Boscaiu, M.A. Naranjo, E. Estrelles, J.M. Belles and P. Soriano, 2004. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ., 58: 463-481.
CrossRefDirect Link - Franco, J.A., S. Banon, M.J. Vicente, J. Miralles and J.J. Martinez-Sanchez, 2011. Root development in horticultural plants grown under abiotic stress conditions-A review. J. Hortic. Sci. Biotechnol., 86: 543-556.
CrossRefDirect Link - Mohammadkhani, N. and R. Heidari, 2008. Water stress induced stomatal closure in two maize cultivars. Res. Biol. Sci., 3: 750-754.
Direct Link - Al Hakimi, A., P. Monneveux and G. Galiba, 1995. Soluble sugars, proline and relative water content (RCW) as traits for improving drought tolerance and divergent selection for RCW from Triticum polonicum into Triticum durum. J. Genet. Breed., 49: 237-244.
Direct Link - Yilmaz, H. and A. Kina, 2008. The influence of NaCl salinity on some vegetative and chemical changes of strawberries (Fragaria x ananssa L.). Afr. J. Biotechnol., 7: 3299-3305.
Direct Link - Turner, S.J. and A.M. Schwarz, 2006. Biomass development and photosynthetic potential of intertidal Zostera capricorni in New Zealand estuaries. Aquat. Bot., 85: 53-64.
CrossRefDirect Link - Desclaux, D., T.T. Huynh and P. Roumet, 2000. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci., 40: 716-722.
CrossRefDirect Link - Westgate, M.E., 1994. Water status and development of the maize endosperm and embryo during drought. Crop Sci., 34: 76-83.
CrossRefDirect Link - Mirzaei, A., R. Naseri and R. Soleimani, 2011. Response of different growth stages of wheat to moisture tension in a semiarid land. World Applied Sci. J., 12: 83-89.
Direct Link - Parker, W.C. and S.G. Pallardy, 1985. Drought-induced leaf abscission and whole-plant drought tolerance of seedlings of seven black walnut families. Can. J. For. Res., 15: 818-821.
CrossRefDirect Link - Pandey, P.K., J.W. Maranville and A. Admou, 2001. Tropical wheat response to irrigation and nitrogen in a Sahelian environment. I. Grain yield, yield components and water use efficiency. Eur. J. Agron., 15: 93-105.
CrossRefDirect Link - Kage, H., M. Kochler and H. Stutzel, 2004. Root growth and dry matter partitioning of cauliflower under drought stress conditions: Measurement and simulation. Eur. J Agron., 20: 379-394.
CrossRefDirect Link - Zeid, I.M. and Z.A. Shedeed, 2006. Response of alfalfa to putrescine treatment under drought stress. Biol. Plant, 50: 635-640.
CrossRefDirect Link