
W. Manggoel
Department of Agriculture Technology, College of Agriculture, P.M.B. 001, Garkawa, Plateau State, Nigeria
M.I. Uguru
Department of Crop Science, Faculty of Agriculture, University of Nigeria, Nsukka, Nigeria
International Journal of Plant Breeding and Genetics
Year: 2012 | Volume: 6 | Issue: 1 | Page No.: 1-16







ABSTRACT
The Nigerian vegetable cowpea accessions differ in phenology with respect to their reactions to day-length. Some are Day Neutral (DN) while others are Short Day (SD) plants. Field experiments were therefore carried out to investigate possible influence of maternal effects on the inheritance of flowering time in cowpea. The distribution of the progenies with respect to days to flowering showed combine effect of both maternal and nuclear influence on the inheritance of flowering time in cowpea. The parental lines, F1, F2 and backcross populations differed significantly (p<0.05) in days to flowering and other yield traits assessed. Days to first and 50% flowering for SD accessions were delayed significantly (p<0.05) compared to DN accessions. The result of the principal component analysis revealed that the most effective characters for distinguishing the parents and progenies included days to first and 50% flowering, No. of peduncles plant-1, No. of flowers plant-1, No. of pods plant-1 and grain yield. The scatter plot and cluster means of the first two principal axes grouped the cowpea accessions into two main clusters. Cluster I comprised of early flowering (<45 days) DN cowpea accessions that were relatively poor in the yield traits assessed. Conversely, the cowpea accessions in cluster II were late flowering (>45 days) SD but prolific in grain and other yield attributes. The hybrids arising from these two groups of cowpea flowered irrespective of day-length, implying continuous podding and all-year-round harvest.
PDF Abstract XML References Citation
Received: February 28, 2011;
Accepted: August 24, 2011;
Published: October 29, 2011
How to cite this article
W. Manggoel and M.I. Uguru, 2012. Evidence of Maternal Effect on the Inheritance of Flowering Time in Cowpea (Vigna unguiculata (L.) Walp.). International Journal of Plant Breeding and Genetics, 6: 1-16.
DOI: 10.3923/ijpbg.2012.1.16
URL: https://scialert.net/abstract/?doi=ijpbg.2012.1.16
DOI: 10.3923/ijpbg.2012.1.16
URL: https://scialert.net/abstract/?doi=ijpbg.2012.1.16
INTRODUCTION
Flowering is an important physiological process in crop survival and assurance for continuity. Time of flowering is particularly of great importance in annual crops, including cowpea (Vigna unguiculata (L.) Walp), as it is a component of the adaptation of a variety to a particular environment and it also determines pod set and crop yield (Ishiyaku et al., 2005). Information on the inheritance of days to flowering will help breeders develop strategies for improvement of seed yields and their adaptation to various agroclimatological zones (Adeyanju et al., 2007). Plant growth and development, especially flowering, is dependent on the interaction of many complex processes which are influenced by both genetic and environmental factors (Uarrota, 2010). Craufurd et al. (1996) and Mukhtar and Singh (2006) reported that in West and Central Africa, photoperiod is the most important environmental variable affecting time of flowering and that most cowpea varieties under cultivation are unimproved, local types which are photoperiod sensitive. Photoperiod has been reported to influence plant growth characteristics, including flowering. Cha-um and Chalermpol (2007) reported that plant height, leaf length, leaf area and flag leaf area as well as flowering were highly regulated by short-day photoperiod in rice.
Earliness in cowpea (Vigna unguiculata (L.) Walp) is an important agronomic trait since it has been reported to yield the dividends of opening the possibility of successful sole cropping in areas with short rainy season, double/triple cropping in rice and/or wheat based systems. It is also beneficial in relay cropping in areas with relatively longer rainfall after millet, sorghum or maize as well as parallel multiple cropping with cassava, yam and cotton (Adeyanju and Ishiyaku, 2007). If the genetic basis of early and continuous flowering is understood, it can be exploited in the development of cowpea varieties that can flower and pod continually thereby ensuring all year round availability of cowpea for the teaming population in sub Saharan Africa.
Maternal effects have been implicated in the inheritance of quantitative and qualitative characters and responses to selection may be affected by this effect (Etterson and Galloway, 2002). This is because, for many characters, the genotype of the mother via maternal effect account for a considerable portion of the genetically based variation in progeny phenotype. Therefore, selection based on direct genetic effect may not be adequate, as it may lead to omission of potentially important source of genetic variance contributed by the cytoplasm of the maternal strain (Wolf et al., 2002). Mothers often provide much of the environment for their offspring. These maternal effects are predicted to result in unusual evolutionary dynamics in offspring traits if they are themselves heritable (McAdam and Boutin, 2003). A number of characters of an organism are determined in the unfertilized egg and with respect to characters of this type, a hybrid will appear similar to the maternal parent. This means that a mother can influence a trait in her offspring both by the genes she transmits (Mendelian inheritance) and by maternal attributes that directly affect that trait in the offspring (maternal effect). Maternal inheritance can alter the direction, rate and duration of adaptive evolution from standard Mendelian models and its impact on adaptive evolution has not been adequately explored in natural populations (Thiede, 1998).
Previous studies indicating considerable maternal and cytoplasmic effects in plants, especially with respect to inheritance of some quantitative and qualitative characters include non-genetic maternal effects on seed size reported by Weiner et al. (1997). The authors reported that the environment in which the maternal plant is growing influence seed size; with the mean weight of seeds being strongly influenced by the identity of the maternal plants. Similarly, nuclear gene and cytoplasmic factors were linked to the inheritance of petal colour in Brassica (Lu et al., 2004). When crosses were made between yellow-petal Brassica rapa and white-petal Brassica oleiracea, significant additive epistatic and cytoplasmic effects were found. Correns as cited by Mousseau and Fox (1998) also reported a classic instance of maternal inheritance in 4 o’clock plant. When crosses were made among the flowers associated with the three leaf colours: Green, variegated and white; the phenotype of the offspring took after their maternal strain.
Although, maternal effects, its evolutionary consequence and improvement in the fitness of progeny are well known in plants, there is almost no information about their expression in nature (Galloway, 2001a), especially on an important crop like cowpea. This is because most studies on maternal effects have been conducted on temperate crops and these studies were carried out under controlled conditions. While these works have led to an understanding of maternal influence on offspring phenotypic expression, less is known of their contribution to phenotypic variation in nature. For instance, a controlled-environment study found that both maternal and paternal environments influenced seed and germination characters (Galloway 2001b); however, it is not known whether these parental effects are expressed under field conditions. The objective of this study was therefore, to investigate possible influence of maternal effects on the inheritance of time of flowering in cowpea under field conditions.
MATERIALS AND METHODS
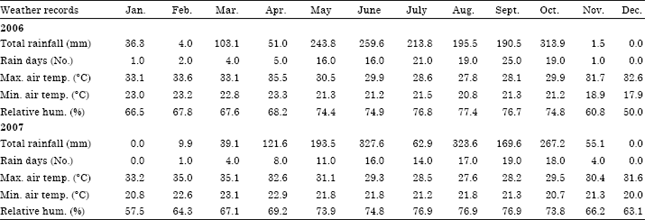
Experimental site and materials: The experiment was carried out in 2006 and 2007 cropping seasons at the Teaching and Research Farm of the Department of Crop Science, Faculty of Agriculture, University of Nigeria, Nsukka. Nsukka is characterized by low land humid tropical conditions and has a bimodal rainfall pattern with annual total of about 1500 mm. The relative humidity is about 70% during the rainy season and falls to 20% during the dry season. The mean annual temperature does not exceed 30°C. The weather data from the University of Nigeria, Nsukka is presented in Table 1. The experimental materials comprised ten cowpea accessions. Six accessions were obtained from Plateau state, north central Nigeria (Latitude 09°26’N; Longitude 9°08’E and altitude of 282.2 m above sea level) and four accessions were obtained from Nsukka, Enugu state, south eastern Nigeria (Latitude 06°52’N; Longitude 07o24’E and altitude of 447.2 m above sea level) (UNN, 2001). The source and description of the cowpea accessions used for the study are presented in Table 2.
| Table 1: | Meteorological records in 2006 and 2007 cropping seasons at Nsukka, Nigeria |
 | |
| Table 2: | Description of the Short Day (SD) and Day Neutral (DN) cowpea accessions used in the study |
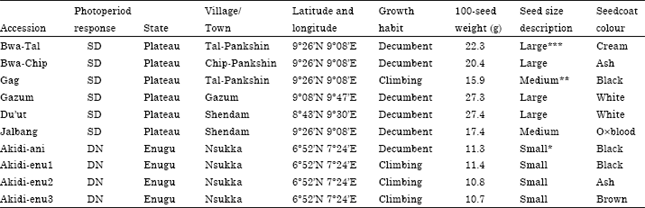 | |
| *Small: <15 g/100-seed weight, **Medium =15-19 g/100-seed weight, ***Large: >20 g/100-seed weight (Omoigii et al., 2006). SD: Short day, DN: Day neutral | |
Characterization and hybridization of the cowpea accessions: The characterization of the cowpea accessions was done using a Randomized Complete Block Design (RCBD) with the cowpea accessions as treatments replicated three times. Hybridization of cowpea accessions was carried out using the method of Utuk (1974). Reciprocal crosses were made between SD and DN cowpea accessions and the F1s generated were allowed to random mate to obtain the F2. Crossing each of the F1 to the respective seed parent (BCP1) and pollen parent (BCP2) were done to generate the backcrosses. The F1, F2, BCs and parental populations were evaluated using the Randomized Complete Block Design with three replications. Entries were in ten-row 15 m long plots. Planting using a 1x1 m intra and inter-row spacing were used. The plots were weeded manually to keep weed pressure low. Other recommended agronomic management practices were adopted for optimum crop growth and development.
Data collection and analysis: Data were collected on days to first and 50% flowering to estimate time of flowering of the parents, F1, F2 and BC populations of the cowpea accessions. Days to first flowering was recorded as the No. of days from sowing to first flower production, defined as corrolla color visible while days to 50% flowering was monitored as the No. of days from sowing to the time 50% of the plants flowered. Data were also collected on No. of flowers plant-1, No. of peduncles plant-1, No. of pods plant-1, pod length, No. of seeds pod-1, 100-seed weight and grain yield. Data for all variables quantified were subjected to Analysis of Variance (ANOVA), using Genstat software version 7.22. Fisher’s Least Significant Difference (F-LSD) was used to detect significant difference between treatment means (Obi, 2002). Test of significance was at 5% probability level. Principal component analysis was performed to identify the variability among the cowpea accessions and their progenies. Hierarchical clustering was performed to classify the cowpea accessions and populations, based on squared Euclidean distances. Cluster mean of a yield trait was computed as the mean of the yield trait of all the candidates in a cluster (Johnson, 1998; Manly, 1994). Variances of the F1 and F2 populations were calculated according to Little and Hills (1978). Bartlett’s test (Snedecor and Cochan, 1989) of homogeneity and Chi square test was done to ascertain the goodness of fit to the expected phenotypic ratios in the F2 and BC populations.
RESULTS
The mean values of temperature in the study area in 2006 and 2007 were 28.7 and 29.8, respectively. This temperature is near optimum values for reproductive development of cowpea (Ishiyaku et al., 2005). The photoperiod ranged from 10 to 13.4 h day-1. The lower limit of this photoperiod (10 h day-1) which falls between the months of September and November provides the short day condition while the upper limit, 13.4 h day-1 recorded between April and August provides the long day condition.
Parental differences among the 10 cowpea accessions in days to first and 50% flowering for the two cropping seasons are presented in Table 3. The cowpea accessions differed significantly (p<0.05) in days to first flowering. The mean days to first flowering for DN cowpea accessions ranged from 40-42 days while the SD accessions had significant (p<0.05) delay, as it took between 56-66 days before they flowered. The earliest No. of days to first flower of 40 days was recorded by Akidi-ani and Akidi-enu 2. This value was statistically similar to those of other DN accessions (Akidi-enu1, 41 and Akidi-enu3, 42 days). The SD accession, Jalbang recorded the highest No. of days to first flower of 66 days which was equally similar to those of other SD accessions (Bwa-Tal, 61; Bwa-Chip, 58; Gag, 59; Gazum, 64 and Du’ut, 56 days) but differed significantly from days to first flowering of the DN accessions.
| Table 3: | Differences among 10 Short Day (SD) and Day Neutral (DN) cowpea accessions in days to flowering for two growing seasons |
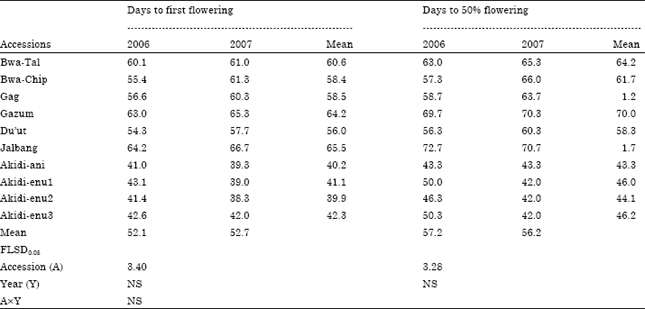 | |
| NS: Not significant | |
Days to 50% flowering followed the same trend as SD accessions took longer time (ranging from 58-72 days) to flower when compared to the DN accessions (43-46 days). For instance, the SD accession Jalbang recorded the lengthiest No. of days to 50% flowering of 72 days while a DN accession, Akidi-ani took 43 days for 50% of the plants to flower. Generally, cowpea accessions did not differed significantly in days to first and 50% flowering within a photoperiodic group. Significant differences in days to first and 50% flowering were however, observed between the two photoperiodic groups. There was however no significant difference between the two cropping years as well as the interactions between the cowpea accessions and years.
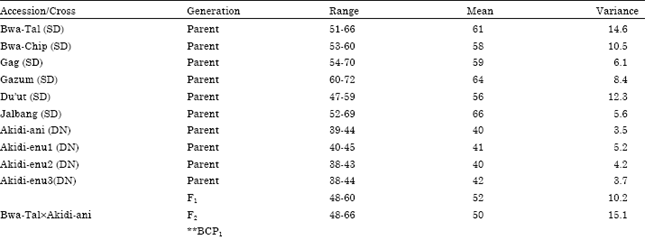
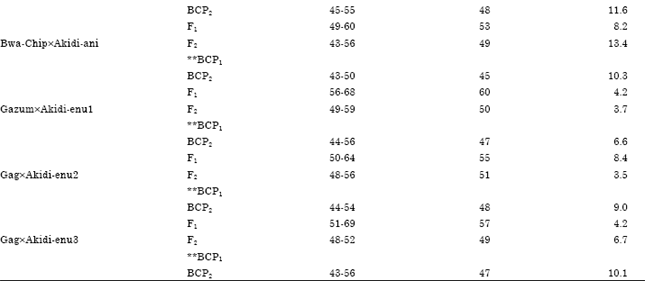
The mean, range and variance of days to first flowering of the F1, F2 and BC populations are presented in Table 4 and 5. The mean days to first flowering of the F1, F2 progenies differed significantly among the reciprocal crosses. The mean days to first flowering of the progenies were maternally influenced. When day neutral accessions were used as mothers in crosses the mean days to first flowering of the progenies took after their mothers. For instance, the cross Akidi-anixBwa-Tal (Table 4) produced F1 offspring that flowered early (42 days) and was statistically similar with the days to first flowering of its maternal line (Akidi-ani, 40 days). Similarly, the reciprocal of this cross, Bwa-TalxAkidi-ani (Table 5), produced progenies that flowered late (52 days) like its mother (61 days). Generally, photoperiod insensitive (DN) accessions used as the maternal parents produced day neutral F1 plants that flowered earlier (37-42 days) while Photoperiod Sensitive (SD) accessions produced offspring whose days to flowering were delayed (49-60 days), just like their mothers. The behaviour of the offspring followed a similar trend with respect to days to 50% flowering.
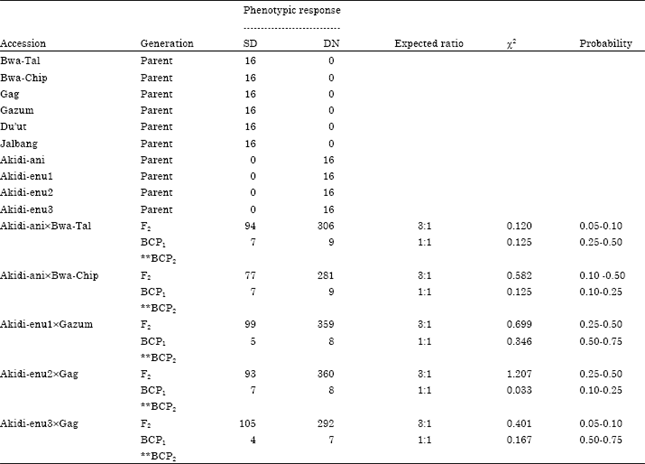
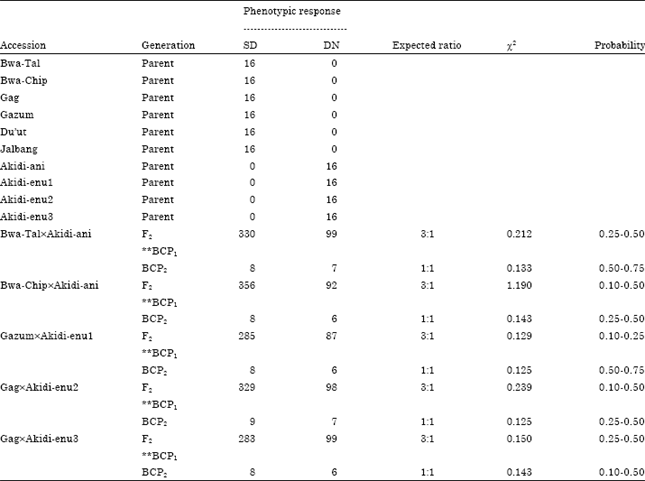
The Segregation patterns for days to first flowering in F2 and in the two backcrosses are presented in Table 6 and 7. The F2 phenotypic response gave a good fit to the 3:1 Mendelian ratio in favour of the maternal parent in the reciprocal crosses involving the two photoperiodic groups. For instance, in the cross Akidi-anixBwa-Tal, the F2 population of 306 DN: 94 SD is a good fit to a 3:1 ratio (χ2 = 0.120, p = 0.10-0.25).
| Table 4: | Mean, Variance and Range of days to first flowering for different populations of crosses between DN and SD cowpea accessions (DN is the maternal parent) |
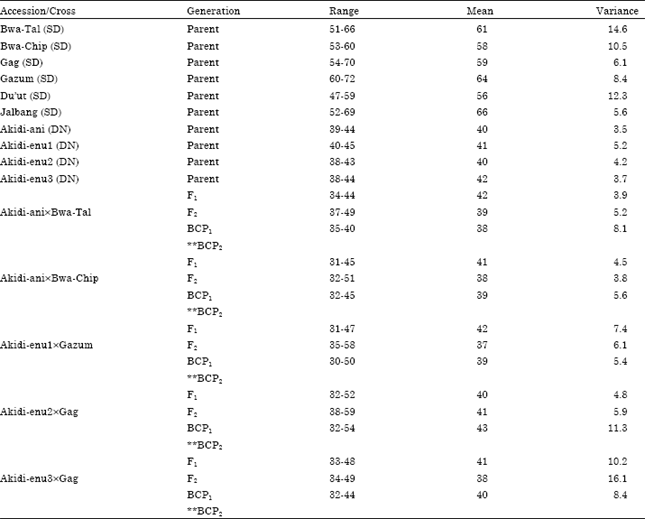 | |
| **BCP2: Not generated because the SD parent did not flower | |
| Table 5: | Mean, Variance and range of days to first flowering for different populations of crosses between DN and SD cowpea accessions (SD is the maternal parent) |
 | |
 | |
| **BCP1: Not generated because the SD parent did not flower | |
| Table 6: | Segregation pattern of days to first flowering of F2 and BC populations of reciprocal crosses between DN and SD cowpea accessions (DN is the maternal parent) |
 | |
| **BCP2: Not generated because the SD parent did not flower | |
| Table 7: | Segregation pattern of days to first flowering of F2 and BC populations of reciprocal crosses between DN and SD cowpea accessions (SD is the maternal parent) |
 | |
| **BCP1: Not generated because the SD parent did not flower | |
Similarly, the F2 population of the cross GagxAkidi-enu2 gave 329 SD plants: 98 DN plants, another good fit to 3:1 ratio (χ2 = 0.239, p = 0.10-0.50). When the F1 progenies were backcrossed to the parents, the phenotypes of the BCP1 and BCP2 progenies segregated at equal ratio of 1:1. However, backcross that required the use of SD parents were not generated due to failure of the parents to flower during the long-day period.
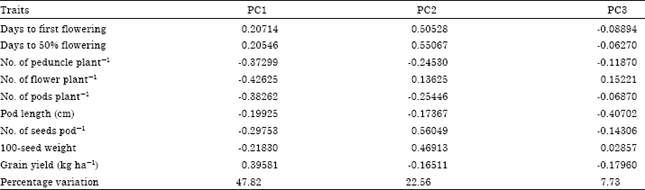
Principal Component Analysis (PCA): The result of the principal component analysis for the 10 cowpea accessions averaged over two cropping seasons are presented in Table 8. The PC1, PC2 and PC3 accounted for 47.82, 22.56 and 7.73% of the variation, respectively. The contribution of the traits towards the diversity of the cowpea accessions revealed that in the first principal component axis, the traits with the highest loadings were No. of peduncles plant-1, No. of flowers plant-1, No. of pods plant-1 and grain yield. This axis is regarded as productivity and yield axis since it loaded high for the yield component traits assessed. The second principal component axis weighed highest in days to first flowering, days to 50% flowering, No. of seeds pod-1 and 100-seed weight. The third principal component axis loaded high for pod length. The two-dimensional scatter plot analysis conducted on the yield traits apparently grouped the cowpea accessions into two clusters (Fig. 1) with the accession Jalbang as an outlier that did not belong to any of the clusters.
| Table 8: | Eigenvector values for principal components using yield traits for 10 cowpea accessions averaged over two cropping seasons |
 | |
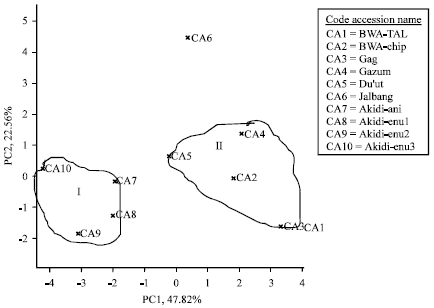 | |
| Fig. 1: | Cluster diagram showing the distribution of SD and DN cowpea accessions averaged over two cropping seasons |
Cluster I comprised the DN cowpea accessions: Akidi-ani, Akidi-enu1, Akidi-enu2 and Akidi-enu3; while cluster II consist of the SD accessions, Bwa-Tal, Bwa-Chip, Gag, Gazum and Du’ut. The cowpea accessions in the different specific groups appear to maintain some level of distance from one another.
The cluster mean (mean of a yield trait of all candidates in a cluster) values for the yield traits of the 10 cowpea accessions evaluated are presented in Table 9. The cluster means show that cluster I comprised early flowering cowpea accessions (Akidi-ani, Akidi-enu1, Akidi-enu2 and Akidi-enu3) that were poor in all other yield traits assessed. Conversely, the cowpea accessions in cluster II (Bwa-Tal, Bwa-Chip, Gag, Gazum and Du’ut) were late flowering and were excellent in the production of peduncles plant-1, flowers plant-1, pods plant-1, pod length, seeds pod-1, 100-seed weight and grain yield. Although, the cowpea accession, Jalbang did not belong to any of the clusters, it flowered late and had very poor record of the assessed agronomic traits.
The principal component analysis and total percentage variation of yield traits for the first three principal axes of the F1 and F2 progenies are presented in Table 10.
| Table 9: | Clusters means for nine yield traits in 10 cowpea accessions averaged over two cropping seasons |
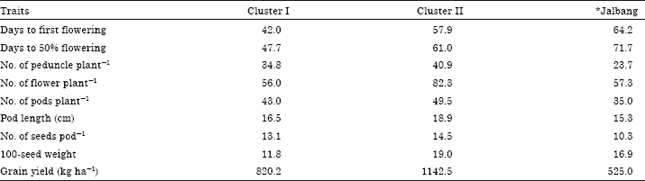 | |
| *Jalbang: Outlier | |
| Table 10: | Eigenvector values for principal component using nine yield traits in the F1 and F2 progenies |
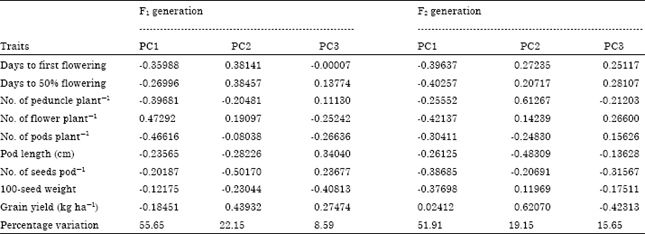 | |
In the F1 progenies, the PC1, PC2 and PC3 accounted for 55.65, 22.15 and 8.59% of the variation, in that order; while the F2 progenies recorded 51.91, 19.15 and 15.65% for the first, second and third principal components, respectively.
In the F1 offspring the first principal component axis had high loadings for No. of peduncles plant-1, No. of flowers plant-1 and No. of pods plant-1. The second principal axis weighed highest in days to first flowering, days to 50% flowering, No. of seeds pod-1 and grain yield. The third principal component axis loaded high for pod length and 100-seed weight. In the F2 progenies, PC1 loaded high for days to flowering, No. of flowers plant-1, No. of pods plant-1, No. of seeds pod-1 and 100-seed weight. The PC2 had high values for No. of peduncles plant-1 and pod length; while PC3 had high loadings for No. of seeds pod-1 and grain yield (Table 10).
The intra-population variability evaluated by cluster analysis conducted on the yield traits of the progenies effectively grouped the genotypes into two clusters (Fig. 2, 3), except for the outlier, GazumxAkidi-enu1 which did not belong to any of the clusters. In both the F1 and F2 offspring, cluster I comprised crosses with DN as their maternal parent; while cluster II consist of progenies whose maternal parents were SD plants. The members of cluster 1 were offspring from the following crosses: Akidi-anixBwa-Tal, Akidi-anixBwa-Chip, Akidi-enu1xGazum, Akidi-enu2xGag and Akidi-enu3xGag; while cluster II comprised of progenies from Bwa-TalxAkidi-ani, Bwa-ChipxAkidi-ani, GagxAkidi-enu2 and GagxAkidi-enu3.
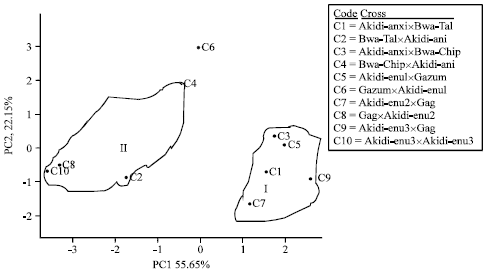 | |
| Fig. 2: | Cluster diagram showing the distribution of F1 progenies in crosses between SD and DN cowpea accessions |
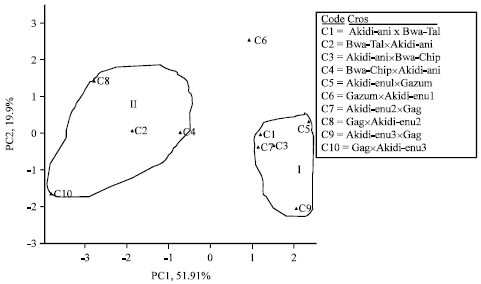 | |
| Fig. 3: | Cluster diagram showing the distribution of F2 offspring in crosses between SD and DN cowpea accessions |
The cluster means for the yield traits in the F1 and F2 progenies of the reciprocal crosses between SD and DN cowpea accessions are presented in Table 11. The cluster means (mean of yield traits of all candidates in a cluster) show that cluster I comprised early flowering F1 and F2 progenies. The offspring in cluster II were late flowering with prolific potentials for the production of peduncles plant-1, flowers plant-1, pods plant-1, pod length, seeds pod-1, 100-seed weight and grain yield. Although, the cross, GazumxAkidi-enu1 did not belong to any of the clusters, it flowered late and was prolific in the No. of peduncles plant-1 and flower production. Based on the cluster means, the F1 and F2 progenies with DN as their maternal parent (cluster I) had significantly higher mean No. of peduncles plant-1, flowers plant-1, pods plant-1, pod length, seeds pod-1, 100-seed weight and grain yield, compared to their parental lines.
| Table 11: | Cluster means for nine yield traits in F1 and F2 progenies of reciprocal crosses between SD and DN cowpea accessions |
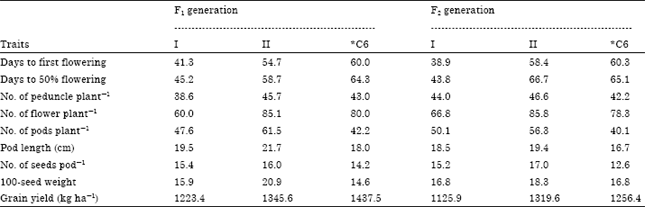 | |
| *C6: GazumxAkidi-enu1 | |
DISCUSSION
The number of days from sowing to flowering is of great importance in cowpea, as it affects pod set and crop yield. In this study, days to first flowering in SD cowpea accession was prolonged significantly (p<0.05) when compared to those of DN accessions. For instance, the No. of days from sowing to first flowering in Bwa-Tal averaged 61 days while that of Akidi-ani was 40 days. The 21 days difference between these accessions in this study is greater than the 11 days reported by Ehlers and Hall (1997) for some cowpea genotypes grown under inductive photothermal environments, with temperature of 24-30°C and photoperiod of 11 h day-1. The wide gap in days to first flowering between the SD and DN cowpea accessions can be linked to differences in factors other than Mendelian inheritance. The differences in days to flowering could be a reflection of innate attributes that are associated with the different photoperiodic groups. The range is equally not close to that reported by Craufurd et al. (1996) for genotypes grown under inductive photoperiods. The authors reported a narrow genetic variation in terms of days to flowering in cowpea. The early flowering in DN accessions (<45 days) and the delayed flowering in SD accessions (>45 days) obtained in this study is in agreement with an earlier photoperiodic grouping by Singh (1993) that genotype whose days to first flowering is greater than 45 are photoperiod sensitive while those that flower in less than 45 days are photoperiod-insensitive or day neutral. The No. of days to 50% flowering in SD cowpea accessions ranged from 58-72 Days after Planting (DAP) is at variance with the report of Oyekanmi and Sangodoyin (2007). The authors evaluated two SD cowpea genotypes and reported that the average No. of days to 50% flowering ranged from 52-56 DAP.
The maternal inclination with respect to No. of days to flowering in the offspring implied that progenies from crosses with SD accessions as maternal parents did not flower until a certain critical photoperiod (short day) was attained. The F1 progenies of the cross, Akidi-anixBwa-Tal in this study recorded mean days to first flowering of 42 which corresponded with the mean No. of days that its maternal parent (Akidi-ani) flowered. The reciprocal cross (Bwa-TalxAkidi-ani) produced F1 progenies with mean days to first flowering of 52 days which was similar to its DN mother (Bwa-Tal). Similarly, when DN accession, Akidi-enu1, was crossed with a SD accession, Gazum with the former as the maternal parent, the F1 progeny had days to first flowering within the range that the day neutral parent flowered. These observations have two possible explanations: First, is the obvious interference of the maternal parent in the behavior of the progeny. The female gamete is usually larger than the male gamete and provides the cytoplasm for the developing embryo. Within the maternal cytoplasm are factors that are released by nuclear genes of the female. These factors produce specific effects upon the developing embryo that eventually result in the different responses of the progenies to day length. Secondly, the female cytoplasm has two important organelles, the mitochondria and the chloroplast that contain DNA. Phenotypes that are controlled by such nuclear factors found in the cytoplasm of the female exhibit maternal inheritance. The distribution of the F1 progenies showed clear evidence of such cytoplasmic nuclear factors in full operation in the inheritance of time to flowering in crosses between DN and SD cowpea accessions. Galloway (2001a), Ishiyaku and Singh (2001), Mosjidis and Yermanos (2004), Thiede (1998), Mousseau and Fox (1998) and Schmitz and Kowallik (1986) have stressed on the significant contributions of the maternal parent in the inheritance of traits. This justifies why Wolf et al. (2002) reported that selection based on direct genetic effect alone is insufficient and misleading as it may lead to omission of potentially important source of genetic variance contributed by the cytoplasm of the maternal strain.
Mendel’s pioneer experiments on the inheritance of monogenic traits in crosses involving a pair of contrasting character showed that the F2 progenies would segregate at a ratio of 3:1. The 3:1 F2 segregation ratio of a monohybrid cross was obtained in this study. The segregation in the F2 is however at variance with the postulation of Galloway (2001b), who suggested that maternal effects may sometimes cause delay in the expression of the phenotype by one generation (from F2 to F3). The involvement of both maternal influence and Mendelian inheritance, in this study, agrees with the report of Lu et al. (2004) in Brassica spp. The authors reported that both nuclear gene and cytoplasmic factors were linked to the inheritance of petal colour in that plant. The 1:1 segregation ratio obtained in the phenotypes of the backcross progenies implied that using the backcross breeding procedure the DN accessions could be used as the adapted recurrent parent on to which desirable traits (e.g., high grain yield) from the SD non-recurrent (donor) parent can be inculcated. However, backcross that required the use of SD parents were not generated due to their failure to flower during the long-day period; thus confirming their photoperiod sensitivity.
The PCA is perhaps the most useful statistical tool for screening multivariate data with significant correlations (Johnson, 1998). Information obtained through Principal Component Analysis (PCA) may assist the plant breeders to identify a limited No. of highly differentiated population for use in hybridization and selection programs. In the present study, the first three principal components accounted for 89.25% of the variability among the 10 cowpea accessions evaluated. The most effective characters for distinguishing among the cowpea accessions included days to first flowering, days to 50% flowering, No. of peduncles plant-1, No. of flowers plant-1, No. of pods plant-1 and grain yield. All these characters showed maximum contribution towards total divergence among the cowpea accessions. The variability of some cowpea cultivars have been described based on the aforementioned traits (Ubi et al., 2001; Ishiyaku et al., 2005; Omoigii et al., 2006). Using the PCA some of these characters have also been used extensively to characterize bambara groundnut (Oyiga et al., 2010) and Cucurbita (Aruah et al., 2010) accessions in Nigeria.
The distribution of the cowpea accessions along the first two principal axes revealed a reasonable agreement with the hierarchical cluster. The intra-population variability evaluated by cluster analysis conducted on the quantitative traits grouped the accessions into two clusters. However, the accession, Jalbang alienated itself from the clusters thereby suggesting that the accession is genetically independent from the other cowpea accessions. The cluster means show that cluster I comprised early flowering cowpea accessions that recorded low yield of all other quantitative traits assessed. Conversely, the cowpea accessions in cluster II were essentially late flowering but prolific in the production of yield determining traits such as peduncles plant-1, flowers plant-1, pods plant-1 and seeds pod-1. These had direct bearing with grain yield as the late flowering genotypes were endowed with higher grain yield than the early flowering genotypes. These results also indicated that the reproductive traits assessed had significant contributions towards grain yield.
The result of PCA of the F1 and F2 generation showed that the most effective characters for distinguishing among the progenies are similar to those of the parental lines. They include days to first flowering, days to 50% flowering, mean No. of peduncles plant-1, flowers plant-1, pods plant-1, pod length, seeds pod-1, 100-seed weight and grain yield. The PCA also effectively grouped the F1 and F2 offspring into two main clusters. The cluster means showed that cluster I comprised the early flowering F1 and F2 progenies. The offspring in cluster II were essentially the late flowering and were prolific in the production of yield determining traits. This study has provided a good foundation for cowpea breeding; pure line selection method could be adopted by plant breeders to isolate the promising varieties from these local cowpea accessions for cowpea growers. This will provide a new research latitude in cowpea breeding in the future. The selected genotypes will serve as veritable materials for hybridization to improve the poorly but agronomically good exotic cowpea lines.
CONCLUSION
The study established the accessions from Plateau state as Short Day (SD) plants and those from Enugu state as Day Neutral (DN) plants. The DN cowpea accessions were poor yielding in contrast to the SD accessions with delayed flowering but better yielding potentials. The production of hybrids between these two groups of cowpea would make a mark in the current drive for sustainable crop yield for the teaming population in the sub Saharan Africa. The emerging progenies would flower irrespective of day length with inbuilt endowments for enhanced yield determining attributes.
REFERENCES
- Adeyanju, A.O. and M.F. Ishiyaku, 2007. Genetic study of earliness in cowpea (Vigna unguiculata L. Walp) under screen house condition. Int. J. Plant Breed. Genet., 1: 34-37.
CrossRefDirect Link - Adeyanju, A.O., M.F. Ishiyaku and L.O. Omoigui, 2007. Inheritance of time to first flower in photo-insensitive cowpea (Vigna unguiculata (L.) Walp.). Asian J. Plant Sci., 6: 435-437.
CrossRefDirect Link - Aruah, C.B., M. I.Uguru and B.C. Oyiga, 2010. Variations among some Nigerian Cucubita landraces. Afr. J. Plant Sci., 4: 374-386.
Direct Link - Cha-um, S. and K. Chalermpol, 2007. An alternative protocol for flowering induction of Thai Indica rice (Oryza sativa L. sp. indica) using a short-day photoperiod treatment. J. Agron., 6: 68-74.
CrossRefDirect Link - Ehlers, J.D. and A.E. Hall, 1997. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res., 53: 187-204.
CrossRefDirect Link - Etterson, J.R. and L.F. Galloway, 2002. The influence of light on paternal plants in Campanula Americana (Campanulaceae): Pollen characteristics and offspring traits. Am. J. Bot., 89: 1899-1906.
Direct Link - Galloway, L.F., 2001. Parental environmental effects on life history in the herbaceous plant Campanula americana. Ecology, 82: 2781-2789.
CrossRefDirect Link - Galloway, L.F., 2001. The effects of maternal and paternal environments on seed characters in the herbaceous plant Campanula Americana (Campanulaceae). Am. J. Bot., 88: 832-840.
Direct Link - Ishiyaku, M.F., B.B. Singh and P.Q. Craufurd, 2005. Inheritance of time to flowering in Cowpea (Vigna unguiculata (L.) Walp.). Euphytica, 142: 291-300.
CrossRefDirect Link - Little, T.M. and F.J. Hills, 1978. Agricultural Experimentation Design and Analysis. 1st Edn., John Wiley and Sons, New York, USA., pp: 42-45.
Direct Link - Lu, C.M., L. Zhang, L. Liu and M. Kato, 2004. Genetic analysis of petal size through genomic manipulation in Brassica. Plant Breed., 123: 495-496.
CrossRefDirect Link - McAdam, A.G. and S. Boutin, 2003. Maternal effects and the response to selection in red squirrels. Proc. R. Soc. Lond., 271: 75-79.
CrossRef - Mosjidis, J.A. and D.M. Yermanos, 2004. Maternal effects and cytoplasmic inheritance of oleic and linoleic acids content in sesame. Euphytica, 33: 427-432.
CrossRefDirect Link - Mousseau, T.A. and C.W. Fox, 1998. The adaptive significance of maternal effects. Trends Ecol. Evol., 13: 403-407.
CrossRef - Mukhtar, F.B. and B.B. Singh, 2006. Influence of photoperiod and gibberellic acid (GA3) on the growth and flowering of cowpea [Vigna unguiculata (L.) Walp]. J. Food Agric. Environ., 4: 201-203.
Direct Link - Omoigui, L.O., M.F. Ishiyaku, A.Y. Kamara, S.O. Alabi and S.G. Mahammed, 2006. Genetic variability and heritability studies of some reproductive traits in cowpea (Vigna unguiculate (L.) Walp.). Afr. J. Biotechnol., 5: 1191-1195.
Direct Link - Oyekanmi, A.A. and O.S. Sangodoyin, 2007. Evaluation of advanced lines of cowpea (Vigna unguiculata (L.) Walp) for agronomic traits and grain yield in the transition zone of Nigeria. Asian J. Plant Sci., 6: 163-167.
CrossRefDirect Link - Oyiga, B.C., M.I. Uguru and C.B. Aruah, 2010. Studies on the floral traits and their implications on pod and seed yields in bambara groundnut (Vigna subterrenea (L.) Verdc). Aust. J. Crop Sci., 4: 91-97.
Direct Link - Schmitz, U.K. and K.V. Kowallik, 1986. Plastid inheritance in Epilobium. Curr. Genet., 11: 1-5.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1989. Statistical Methods. 8th Edn., Iowa State University Press, Ames, IA, USA, ISBN-13: 978-0813815619, Pages: 524.
Direct Link - Thiede, D.A., 1998. Maternal inheritance and its effects on adaptive evolution: A quantitative genetic analysis of maternal effects in natural plant population. Evolution, 13: 908-1015.
Direct Link - Uarrota, V.G., 2010. Response of cowpea (Vigna unguiculata L. Walp.) to water stress and phosphorus fertilization. J. Agron., 9: 87-91.
CrossRefDirect Link - Ubi, B.E., H. Mignouna and G. Obigbesan, 2001. Segregation for seed weight, pod length and days to flowering following a cowpea cross. Afr. Crop Sci. J., 9: 463-470.
Direct Link - Weiner, J., S. Martinez, H. Muller-Scharer, P. Stoll and B. Schmid, 1997. How important are environmental maternal effects on plants? A study with Centaurea maculosa. J. Ecol., 85: 133-142.
Direct Link - Wolf, J.B., T.T. Vaughn, L.S. Pletscther and J.M. Cheverud, 2002. Contribution of maternal effect QTL to genetic architecture of early growth in mice. Heredity, 89: 300-310.
CrossRefDirect Link
Dasbak, Madang Ayuba Reply
A good article worth publishing in a reputable Journal
Paul Longmil Reply
The work is a masterpiece, published in a relevant rep[table international Journal.