
D. Nyadanu
Cocoa Research Institute of Ghana, Ghana
B. Adomako
Cocoa Research Institute of Ghana, Ghana
M. K. Assuah
Cocoa Research Institute of Ghana, Ghana
Yaw Opoku Asiama
Department of Crop Science, University of Cape Coast, Cape Coast, Ghana
E. Brenyah
Cocoa Research Institute of Ghana, Ghana
Y. Adu-Ampomah
Cocoa Research Institute of Ghana, Ghana
International Journal of Plant Breeding and Genetics
Year: 2011 | Volume: 5 | Issue: 4 | Page No.: 286-301







ABSTRACT
The objective of this study was to observe pattern of similarity of reaction to Phytophthora palmivora infection among 25 international cocoa genotypes in five laboratory experiments and a field observation at the Cocoa Research Institute of Ghana by hierarchical cluster analyses. The genotypes were clustered into two distinct populations in all the screening methods suggesting of genetic limitations of the cocoa genotypes studied. The hierarchical cluster analyses of all the screening tests sub-clustered the susceptible and resistant clones differently except in field infection where there was some inconsistency. This is indicative of the possibility of using any of the screening methods to observe similarity of reaction of cocoa genotypes to Phytophthora species. The inconsistencies in field infection are suggestive of the environmental factors influencing the genetic base of infection of Phytophthora species in the field. The resistant and susceptible genotypes were distributed among centres of origin and genetic groups showing that resistant and susceptible genotypes were widely dispersed among origins and genetic groups. The shapes of pods of resistant genotypes were made up of calabacillo and cundeamor while that of susceptible clones was Amelonado. The remaining of the clones, classified as moderately resistant were the most distantly related clones.
PDF Abstract XML References Citation
Received: August 05, 2010;
Accepted: May 17, 2011;
Published: July 09, 2011
How to cite this article
D. Nyadanu, B. Adomako, M. K. Assuah, Yaw Opoku Asiama, E. Brenyah and Y. Adu-Ampomah, 2011. Analysis of Reaction to Phytophthora palmivora Infection among the International Clone Trial Selections of Cocoa in Ghana. International Journal of Plant Breeding and Genetics, 5: 286-301.
DOI: 10.3923/ijpbg.2011.286.301
URL: https://scialert.net/abstract/?doi=ijpbg.2011.286.301
DOI: 10.3923/ijpbg.2011.286.301
URL: https://scialert.net/abstract/?doi=ijpbg.2011.286.301
INTRODUCTION
Ghana is the world’s second largest producer of cocoa after Cote’d’Ivoire and the crop occupies a key position in the country’s economy in terms of foreign exchange generation domestic, income and source of revenue for the provision of socio-economic infrastructure. The cocoa sector employs about 60% of the national agricultural labour force (Appiah, 2004). The chocolate industry in Europe and elsewhere depends heavily on imports of dry cocoa beans from West Africa. Even though there is an increasing demand for cocoa beans (Taylor, 2000), production of cocoa is limited by pests and diseases. In Ghana, production, based on smallholder farmers, has been fluctuating around 681, 000 and 740, 000 tonnes over a decade, with a maximum of 740, 000 tonnes recorded in 2005/2006 cocoa season (Research and Business Development (RBD), 2009). This stagnation is due to several factors, the most important being the pod rot disease caused by Phytophthora megakarya with losses of 60-100% of the crop being common in the absence of control measures. Adomako (2007) reported that yield loss in cocoa was largely due to black pod representing 64.1% of total yield loss. Farmers have continued to be compelled to apply frequent chemical control, with copper-based or systemic fungicides. These are damaging to the environment, increase significantly production cost compared to the limited income from cocoa sales and are therefore not available to all farmers. Cultural practices alone are not effective in controlling black pod disease (Akrofi et al., 1997). Studies conducted by Luterbacher (1994) on removal of leaf litter around cocoa trees and the application of fungicides to the ground showed that these practices had little or no effect on P. megakarya disease progress. This situation clearly explains the need to apply integrated disease control methods and to select materials with acceptable resistance levels against the disease.
Breeders and pathologists have long recognized that host plant resistance is likely to be the most sustainable means of combating the disease (Iwaro and Singh, 2004). During the 1990s, resistance to Phytophthora pod rot received considerable attention. This led to the search for resistant materials and consequently the development of several inoculation methods for the screening of locally collected germplasm. An early screening test based on leaf disc inoculations was developed by Nyasse et al. (1995) that proved to be correlated with field level of infection (Tahi et al., 2000; Nyadanu et al., 2009). Detached cocoa leaf and pod inoculation at penetration and post-penetration stages of infection were also developed by Iwaro et al. (1997a). Iwaro (1996) reported that inoculation of detached pods creates an opportunity for rapid assessment of pod resistance to Phytophthora infection.
There is a difficulty of identifying parents with higher and varied Phytophthora resistance factors. Resistance to Phytophthora species has been found to be largely polygenic (additive) (Tan and Tan, 1990; Akaza et al., 2009), indicating that it might be possible to increase resistance by accumulating the different resistant factors. There is a need to group or cluster parents that are genetically resistant to the disease. This could be done by clustering genetically related individuals in a population in relation to the trait of interest (Mohammadi and Prasanna, 2003; Ramdani et al., 2007). However, it is not known if the groupings of genetically similar cocoa genotypes in relation to resistance to black pod differ among the screening methods.
The objective of this study was to establish whether the genetic groupings of the 25 clones in resistance to black pod disease by hierarchical cluster analysis, would be similar among the screening methods.
MATERIALS AND METHODS
In this study, six screening methods were used to test for P. palmivora resistance in 25 cocoa genotypes. The six methods were: leaf disc test, punch inoculation method of leaf, multiple point inoculation method of cocoa pods, stab inoculation method of cocoa pods, natural field infection of cocoa pods. The phenotypic data from these screening methods were used to do hierarchical cluster analysis.
Experimental site: The experiment was conducted at Tafo in a cocoa clonal plot (K5 extension) (0.6 ha) at the Cocoa Research Institute of Ghana (CRIG) in the Eastern region of Ghana. The plot was established in 2001. Cocoa trees were planted in a Randomized Complete Block Design (RCBD) with 5 replications at 2.5x2.5 m spacing with shade trees and had uniform and optimal cover (approximately 50% of the solar radiation pass through the canopy). No fertilizers were applied. Regular pruning was performed on the exceeding plagiotropic and orthotropic branches, along with the removal of mistletoes and chupons. The soil at the experimental site belongs to the forest ochrosol. The region has a bimodal rainfall pattern with an average range between 1200 and 1930 mm. Relative humidity values of 99 to 100% are generally recorded at night and early morning. Relative humidity values drop to about 70% by mid-day especially on sunny days (CRIG Meteorological Station, 1990).
Cocoa genotypes: The genetically diverse set of 25 cocoa clones (PA 120, MOCORONGO, LECTEEN 37f, ICS 43, SPEC 54i, EQX 78, EET 59H, PA 107, LECTEEN 37f, SCA 6, LAF 1, T85/799, AMAZ 15-15, EQX 3360-3, IFC 5, IMC 47, PA 150, UF 676, VENC 4-4, ICS1, BE 10, MO 20, MAN 15-2, GU 225V and T79/501) used for this study represent the International Clone Trials supported by the CFC/ICCO/Biodiversity project being evaluated in Ghana. Each clone was represented by ten trees in each replication. The genetic group, origin and characteristics of the 25 international clones are presented in Table 1.
Isolation of Phytophthora species from black pod infected pods from the field: Isolation of Phytophthora species was done from naturally infected pods collected from the K-5 extension plot on which the 25 genotypes were planted. The detached diseased pods were washed and blotted dry. The pods were then surface sterilized with 70% ethanol. The infected parts of the pods were cut into 5 mm segments. The segments were then plated on carrot agar medium. All inoculated plates were incubated for 3 days on the laboratory bench. Plates were observed daily for fungal growth from 5mm segments.
| Table 1: | Genetic group, origin and characteristics of the 25 international clone selections |
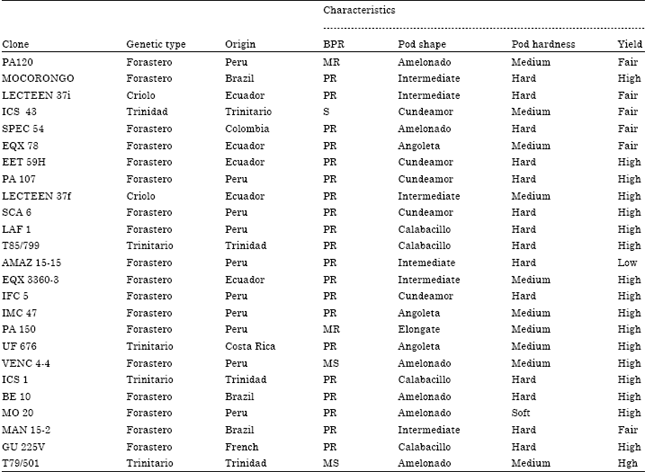 | |
| MR: Moderately resistant, PR: Potentially resistant, S: susceptible, MS: Moderately susceptible. Lower Amazon region: Brazil, Peru and Ecuador Upper Amazon region: Costa Rica, Mexico, Trinidad, Colombia and French | |
Fungal growths from tissue segments were sub-cultured onto another carrot agar medium to obtain pure culture of the isolated fungi. Emergent colonies were examined under light microscope and the Phytophthora specie was identified. Based on the growth of isolate on carrot agar medium, sporangial shape and size and pedicel length, the Phytophthora specie was identified. The isolate was grown on a carrot agar medium and from a ten-day-old culture, a zoospore suspension was obtained by inundating each culture plate (9 cm diameter) with 10 mL sterile distilled water. Such plates were then refrigerated for 25 min (5°C) and transferred into an incubator for 25 min. The plates were examined for zoospore release. The distilled water was poured into a beaker as a zoospore suspension. The concentration of 200,000 per mL zoospores required for the experiments were determined with a hemacyatometer. This concentration of inoculum was reported as optimum in similar inoculations conducted by Screenivasan (1985) and Okey (1996). The pathogenicity of the isolate was maintained by regular inoculation in the laboratory of green mature cocoa pods followed by re-isolation on carrot agar medium. Only one isolate of the Phytophthora specie was used in this study.
Sampling of leaf from the field: The new flushes from bud break of the clones were tagged to establish the ages of the leaves for each experiment. For each of the inoculation series, leaves were collected from all of the 25 clones in each replication. In the field, 15 leaves were harvested from each clone in each replication. The ages of the leaves for each treatment were established by following the growth of young flushes from bud break in the field. After collecting the matured leaves, they were placed in labelled polyethylene bags into which a few drops of distilled water were sprayed beforehand. The bags were then kept in the dark till the next morning. This was done to minimize any effect of leaf sampling time that may occur with large time lapses between harvesting of leaves (Tahi, 2003). The leaves were washed thoroughly with tap water, blotted dry with Whatman number 3 paper and were then surface sterilized with 70% ethanol.
Leaf inoculation tests: The detached leaf inoculations used in this study were leaf disc test, multiple point inoculation of leaf and the punch inoculation of leaf.
Assessment of leaf resistance to P. palmivora by the leaf disc test: Leaf disc inoculation as described by Nyasse et al. (1995) was carried out. Sixteen leaf discs of 1.5 cm in diameter from each clone were made with a cork borer. The sixteen leaf discs were replicated five times, totaling 80 (5x16) discs per clone. Leaf discs were placed with their abaxial (upper) surface upwards on wetted plastic foam in five trays 70 cm long, 60 cm wide and 15 cm high. Discs belonging to the same replication were randomly arranged in groups of 25 within each tray, giving 16x25 = 400 discs per tray. Inoculation was carried out the same day, after preparation of all leaf discs. After the concentration of zoospores were determined with a hemacyatometer and adjusted to 200,000 per mL, droplets of 10 μL were placed on each disc. The discs were incubated at room temperature of 25°C in the plastic trays lined with moist plastic foam and covered with another plastic tray in the laboratory avoiding direct sunlight until observations were carried out. On the sixth day of incubation, disease severity symptom was recorded using a 0-5 assessment scale developed by Nyasse et al. (1995) and rated as 0 = absence of symptoms, 1 = very small necrotic spots, 2 = larger number and size of necrotic spots, 3 = coalescence of brown spots into medium-sized, 4 = large uniform brown lesions and 5 = very large brown lesions, often expanding outside the area covered by the inoculum droplet.
Assessment of leaf resistance to P. palmivora at the penetration level of infection: Leaf resistance at the penetration stage of infection was assessed with the tissue-paper mount method of inoculation as described by Iwaro et al. (1997a).
Mature leaves at the interflush-2 stages as described by Greathouse et al. (1971) were surface sterilized with 70% alcohol and rinsed in sterile distilled water once. The surface sterilized leaves were then placed in plastic trays lined with moist plastic foam, with the abaxial surface facing upwards. From each clone in a replication, 5 leaves were placed in one tray. Tissue paper-mount inoculation of the leaf samples was done by applying a 30 μL drop of zoospore suspension on the leaf surface at the centre over which a piece of tissue paper (area: 1 cm2, thickness: 0.23 mm) was placed to allow a uniform spread of the zoospore suspension within the area covered by the tissue paper. A leaf from each clone was inoculated with sterile distilled water as a control in each experiment. The 25 clones were arranged in a completely randomized design with five replications and incubated at room temperature of 25°C in plastic trays lined with moist plastic foam and covered with another tray. After 6 days of incubation, the tissue paper mounts were carefully removed and the penetration resistance was determined by counting the number of lesions. The experiment was repeated.
Assessment of leaf resistance to P. palmivora at the post-penetration level of infection: Assessment of leaf resistance at the post-penetration stage of infection was based on punch inoculation method (Iwaro et al., 1997a). The area of lesion formed was used as an indication of post-penetration resistance. A 4 mm-diameter hole was punched in the leaf lamina and the hole was covered at the adaxial surface with a spot plaster. The hole in the abaxial surface was filled with a 4 mm-diameter filter paper disc previously immersed in 200,000 zoospores per mL suspension. A leaf from each clone was inoculated with sterile distilled water in place of zoospore suspension as a control. Inoculated leaves were arranged in a completely randomized design with five replications and incubated at room temperature of 25°C in trays lined with moist plastic foam and covered with another tray. After incubation for six days, the lesion areas on leaves were cut and assessed for lesion size with a leaf area meter (MK2, Delta T services, Burnwell and Cambridge, England). This experiment was repeated to confirm the consistency of results obtained.
Sampling of pods from the field: Green mature unripe pods of sizes similar to those of ripe ones at approximately four months old were used as test samples. Pods were harvested with care and kept in labeled plastic bags. The pods were covered with cotton wool in order to avoid surface damage which may occur when many pods are kept together in close contact with each other. The harvested cocoa pods were washed thoroughly with tap water and blotted dry with Whatman Number 3 paper. They were then surface sterilized with 70% ethanol.
Pod inoculation tests: The pod inoculation tests carried out in this study were multiple point inoculation of pod and stab inoculation method of pod (Iwaro et al., 1997a).
Assessment of pod resistance to P. palmivora at the penetration level of infection: Resistance of pod at the penetration level was assessed with detached pods using the multiple-point inoculation (Iwaro et al., 1997a). The multiple point inoculation was performed on the pod surface, in which 10 μL drops of inoculum were placed at 3 points along the ridges with a micropipette. A distance of about 3 cm was maintained between inoculated points to avoid merging of adjacent lesions. A zoospore concentration of 200,000 per mL was used. A pod from each clone was inoculated with sterile distilled water in place of zoospore suspension as a control. The 25 clones were replicated five times (with appropriate controls) and arranged in a completely randomized design. The pods were incubated at 25°C in a 40x60 cm transparent polyethene bags. A beaker of water was kept in the bag with the mouth of the bag closed. After 6 days, pods were assessed for the number of established lesions. The number of established lesions on pods was used as an indication of penetration resistance. The experiment was repeated once.
Assessment of pod resistance to P. palmivora at the post-penetration level of infection: Assessment of pod resistance at the post-penetration stage of infection was based on stab inoculation method (Iwaro et al., 1997a). The area of lesion formed was used as an indication of post-penetration resistance. For the stab inoculation, a standard injury 4mm in size was created on the pod surface with a cork borer. The wounded spot was inoculated with a piece of cotton wool previously immersed in a 200,000 zoospores per mL suspension and covered with a spot plaster. Inoculated pods were arranged in a completely randomized design with five replications and incubated at room temperature of 25°C in a 40x60 cm transparent polyethylene bags. A beaker of water was kept in the bag with the mouth of the bag closed. After incubation for six days, the size of the established lesion was traced on a transparent paper. The lesion size was determined from brown paper cutouts trimmed to the size of each lesion and was measured with a leaf area meter. The experiment was repeated.
Assessment of natural field infection of black pod disease: Natural pod infection in the field has been collected from all the 25 clones in each replication monthly from July to December 2007 and 2008. The diseased, wilted, damaged (feeding) and ripened pods were removed each month. Pods infected by Phytophthora (Bp), Rodent damaged pods (R) and healthy ripe pods (H) were counted each month, with elimination of completely rotten pods after each harvesting round. The percentages (%) of pods affected by Phythophthora pod rot (Bp) were estimated in relation to the total number of pods produced by the tree at the end of the cocoa season:
Data analysis: The data on leaf disc scores, number and size of lesions of P. palmivora infection and field observations of black pod infection were analyzed using the Genstart statistical software version 10.0 to perform Analysis of Variance (ANOVA) after which residual plots were inspected to confirm data conformed to normality. The significance of mean differences among genotypes was evaluated at p<0.05, using DMRT. Hierarchical cluster analyses were carried out to examine similarities and dissimilarities in the reaction patterns of the clones with respect to the P. palmivora with which they had been inoculated. The clustering method of Ward (1963) was used, based on dissimilarity matrices representing absolute values squared Euclidean distances. The analyses were completed for the individual datasets separately. The statistical package SPSS version 14.0 was used (SPSS Inc, 1996).
RESULTS
Differential responses of cocoa genotypes to P. palmivora: Most cocoa genotypes displayed high variation in the range and mean of susceptible reactions when they were inoculated with P. palmivora (Table 2). Furthermore, the data indicated substantial differences among clones with respect to overall level of susceptibility and differential responses.
| Table 2: | Range and mean susceptible reactions of P. palmivora tested on cocoa differential genotypes (Theobroma cacao) by detached leaf and pod tests and field infection (FI) |
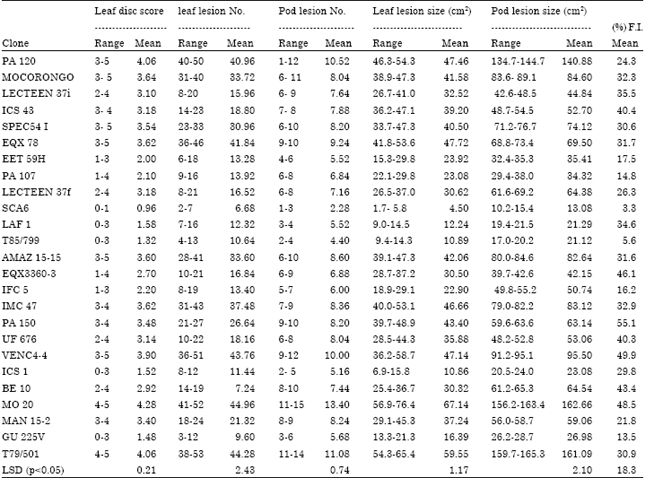 | |
Cultivars MO 20, VENC 4-4, PA 120 and T79/501 exhibited very little differential response, as they were susceptible to P. palmivora in the detached leaf and pod tests.
A low to intermediate infection of P. palmivora was observed on ICS 43, LCTEEN 37f, LCTEEN 37i, UF 676, EQX 3360-3, BE 10, MAN 15-2, IMC 47, MOCORONGO, AMAZ 15-15, SPEC 54i, PA 150 and EQX 78. These cultivars can be regarded as moderately resistant.
The lowest mean infection of P. palmivora across populations and testing methods was observed on cocoa genotypes SCA 6, LAF 1, T85/799, GU 225V, ICS 1, EET 59H, PA 107 and IFC 5 indicating a high level of resistance in these cultivars.
Similarity of reactions of cocoa genotypes to P. palmivora: Similarities and dissimilarities of the cocoa genotypes and their patterns of reaction to P. palmivora were examined by means of hierarchical cluster analysis (Ward, 1963) (Fig. 1 a-f). The cluster analyses were based on similarity matrices representing absolute values squared Euclidean distances. The closer the values in the similarity distances in Fig. 1(a-f) are to 1, the more similar are the respective genotypes.
The dendrograms of the cluster analyses shown in Fig. 1(a-f) reflect the ‘hierarchical structure of relative closeness’ (Peneder, 1995) of the reaction patterns of the cultivars, allowing a more comprehensive view of the similarity of response of the cocoa genotypes to P. palmivora.
 | |
| Fig. 1a: | Cluster analysis of different genotypes based on leaf test |
 | |
| Fig. 1b: | Cluster analysis of different genotypes based on leaf inoculation at pentration level |
 | |
| Fig. 1c: | Cluster analysis of different genotypes based on leaf inoculation at post-penetration level |
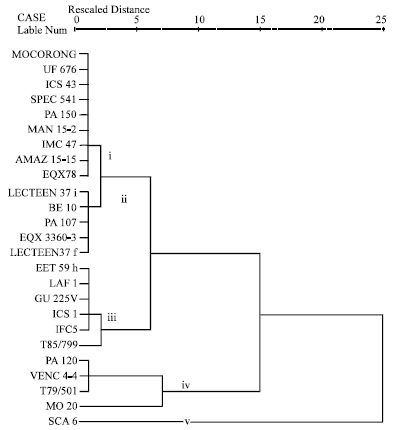 | |
| Fig. 1d: | Cluster analysis of different genotypes based on pod inoculation at pentration level |
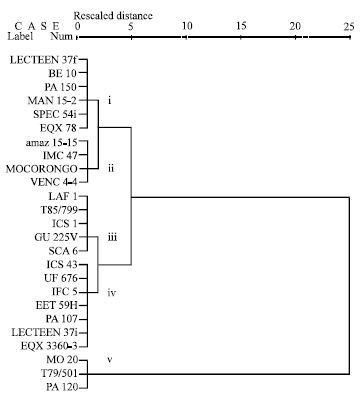 | |
| Fig. 1e: | Cluster analysis of different genotypes based on pod inoculation at post-penetration level |
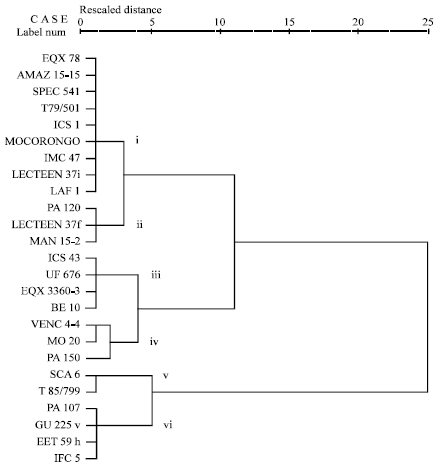 | |
| Fig. 1f: | Cluster analysis of different genotypes based on pooled percentage field infection in 2007 and 2008 |
Two major genotypes clusters were evident in the entire population of clones used in all the tests. It can be seen that the two distinct populations are made up of different genetic groups. The dendrogram shows that the genetic divergence between the two major clusters and is wide with a similarity distance of 25. It is clear from the dendrogram that the clusters were not mere groupings of the genotypes with similar morphological characters but also as distinct genetic groups. Within the sub-clusters of these two clusters, susceptible clones MO 20, VENC 4-4, PA 120 and T79/501 and the resistant clones SCA 6, LAF 1, T85/799, GU 225V, ICS 1, EET 59H, PA 107 and IFC 5 were more similar and most closely related clones. For example, the low squared Euclidean distances for the susceptible clones, when compared with each other, indicate that they reacted similarly to P. palmivora in leaf disc test, leaf inoculation at penetration stage, pod inoculation at penetration stage and pod inoculation at post-penetration stage (Fig. 1a-e). Genotypes MO 20 and T79/501 were also very similar in their reaction patterns but they reacted very differently from Venc4-4 and PA 120 in leaf inoculation at the post-penetration stage (Fig. 1c). The cluster grouping of the susceptible clones in field infection test was somewhat inconsistent; however, genotypes VENC4-4 and MO 20 were grouped to the same sub-cluster whilst T79/501 and PA 120 were grouped in another two distinct sub-clusters (Fig. 1f). Based on leaf disc score and leaf lesion number, five sub-clusters of the cocoa genotypes could be discerned (Fig. 1a, b). SCA 6, LAF 1, T85/799, GU 225V, ICS 1, EET 59H, PA 107 and IFC 5 were generally most similar to each other and very different from most other clones. However, SCA 6 sub-clustered differently from the rest of the clones in both tests.
Likewise, leaf inoculation at the post-penetration stage and pod inoculation at the penetration stage grouped the cocoa genotypes into sub-clusters (Fig. 1c, d). SCA 6, LAF 1, T85/799, GU 225V and ICS 1 were similar to each other, however in pod inoculation at the penetration stage, Sca6 grouped differently from all the other clones (Fig. 1d). In pod inoculation at the post-penetration stage, LAF1, T85/799, ICS 1, GU 225V and SCA 6 resembled each other and were more similar to ICS43, UF 676, IFC 5, EET 59H, PA 107, LCTEEN 37i and EQX 3360-3 than to any other and very different from most other cultivars (Fig. 1e).
Results of field infection formed another two distinct clusters. The second cluster (which is the smaller population) composed of SCA 6, T85/799, GU 225V, PA 107, EET 59H and IFC 5.
SCA 6 and T85/799 sub-clustered from Pa 107, GU 225V, EET 59H and IFC 5 whilst LAF 1 and ICS 1 grouped in another distinct sub-cluster (Fig. 1f).
Among the remaining genotypes, LCTEEN 37f, LCTEEN 37i, ICS 43, UF 676, EQX 3360-3 and BE 10 appeared most similar to each other and always belonged to the same sub-cluster in the screening tests.
DISCUSSION
Using specific resistance properties of individual genotypes is a biologically appropriate and relevant approach for examining the composition, diversity and evolution of cultivars and populations and therefore, widely used (Robinson and Jalli, 1996; Afanasenko et al., 2007).
The reactions of these clones showed a substantial genetic variability for Phytophthora resistance. The clones exhibited significant variation for both the frequency and size of lesions as well as percentage infection in the field. The screening methods were carried out under the same environmental conditions and as such the observed variations in the range and mean of the cocoa genotypes could probably be genetic. The study supports the results of Tan and Tan (1990) and Simmonds (1994) who observed similar variations in their studies on the resistance of cocoa progenies to P. palmivora. The findings also agrees with the observations of Djocgoue et al. (2006) and Akaza et al. (2009) who also reported significant differences in levels of resistance between the genotypes of cocoa progenies. Ahmed et al. (2006) also observed similar findings in reaction of chickpea to Ascochyta blight following detached leaf and whole-plant inoculation with an aggressive isolate of Ascochyta rabiei.
The grouping of the 25 genotypes into two distinct populations is suggestive of genetic limitations of the cocoa genotypes studied. This agrees with the findings of Gizlice et al. (1994) who also observed genetic limitations in their genotypes and stated that the narrow genetic base of modern cultivars has been caused by a limited initial base and several decades of intensive breeding and selection. They further stated that greater than 85% of the genes present in modern cultivars could be traced to a collection of 18 ancestors and their initial progeny. Since genetic variability is necessary for genetic progress, this limitation of genetic diversity may impede further advances in cocoa breeding for resistance to Phytophthora species. Introduction of new sources of genetic variability into breeding programmes may be necessary. Reed and Frankham (2003) stated that genetic diversity have a positive correlation in improvement of crops.
The results of the cluster analyses revealed the susceptible, resistant and moderately resistant clones. The grouping of resistant clones differently from the susceptible clones indicates the resistant clones resembled each other with respect to their black pod reaction patterns when inoculated with zoospores of P. palmivora in all the screening tests used except in field infection where there was somewhat some inconsistency. For example: Fig. 1b, cluster ii- PA 120, T79/501, VENC4-4, Mo 20- all susceptible ; Cluster iv- ICS 1, GU 225V, LAF 1, T85/799, SCA 6, PA 107, IFC 5, EET59H- all generally quite resistant. Cultivars within a particular cluster are likely to share common resistance genes. This is indicative of the possibility of using any of the screening methods to observe similarity of reaction of cocoa genotypes to Phytophthora species. The inconsistencies in field infection are suggestive of the environmental factors influencing the genetic base of infection of Phytophthora species in the field.
Among the resistant clones, ICS 1, GU 225V and T85/799 originated from the Upper amazon region and LAF 1, SCA 6, EET 59H and PA 107 originated from Lower Amazon region. Also, ICS 1 and T85/799 belong to Trinitario genetic group and GU 225V, LAF 1, SCA 6, EET 59H, PA 107 and IFC 5 belong to the Forastero genetic group. However, among the susceptible clones, MO 20, PA 120 and VENC 4-4 originated from Peru in the Lower amazon region and belong to the Forastero genetic group while T79/501 originated from Trinidad in the Upper amazon region and belongs to the Trinitario genetic group. This supports the findings of Adomako (2006) who reported that similarity of resistance factors of progenies could be inferred from the origin of their parents. Chaveerach et al. (2007) also observed genetic diversity and similarity in 15 individuals of Nelumbo nucifera from NE and C Thailand collected according to their geographical areas and the morphological differences among individuals. Parents from the same area of the Amazon Basin in South America are more likely to be closely related. The distribution of resistant and susceptible genotypes within origins and genetic groups showed that resistant and susceptible genotypes were widely dispersed among origins and genetic groups. This finding is in agreement with results of Iwaro (1996) who suggested that an appropriate strategy would therefore permit accumulation of resistance genes from different genetic populations and origins.
In addition, it is interesting to note that the shapes of pods of resistant genotypes were made up of Calabacillo and Cundeamor while that of susceptible clones was Amelonado. Thresh et al. (1988) reported the ‘West African Amelonado’ to be vulnerable to cocoa swollen shoot virus. Van Hall (1932) described Amelonado as broad, short fruits sometimes with a brunt, sometimes acuminated apex. He described Calabacillo as short fruits with smooth pod walls and shallow furrows. The apexes shortly acuminate and sometimes blunt. Ciferri (1929) described Cundeamor as bottle necked pods which are small with a smooth surface. Angoleta is a term used to describe regularly formed fruits with a thick fruit wall. The shape of the pods might explain in part the differences in the reaction of the cocoa genotypes to P. palmivora. Cocoa pods which keep moisture on the pod surface for a longer duration are more prone to Phytophthora species than pods which quickly dispose off moisture. This supports the observations of Agrios (1997) who stated that the moisture level has an important influence on the growth and reproduction of Phytophthora species. The high amount of moisture requirement is satisfied by high relative humidity and free water on the plant surface for species such as Phytophthora infestans and P. palmivora that are pathogens of leaves or pods. This finding is also in agreement with findings of Okaisabor (1974) who observed that a film of water on the leaf for 6-8 h, depending on the temperature, allows infection, penetration, sporangial formation and dispersal of Phytophthora infestans and P. palmivora to occur at high relative humidities. The smooth surface of Calabacillo and Cundeamor pods could be components of factors conferring resistance on the resistant clones. This agrees with the results of Iwaro et al. (1997b) who reported a strong relationship between surface wax, hardness of pod husk and stomatal frequency and resistance to lesion establishment (lesion frequency). Cocoa pods with high deposition of surface wax are smooth.
The remaining of the clones, classified as moderately resistant were the most distantly related clones. This was evidenced by their wide distribution into very different genetic groups and sources of origin. They may likely be sources of rare alleles or rare combinations of alleles which could be useful for cocoa breeders. This supports findings of Tanksley and McCouch (1997) who stated that genetically diverse genotypes are sources of important genes needed for genetic progress. Other researchers have suggested that accessions with phenotypes intermediate between resistant and susceptible genotypes may be due to hybridization between the two (Hymowitz, 1970; Broich and Palmer, 1981). The only additional recognizable pattern among the moderately resistant clones was the clones LCTEEN 37f and LCTEEN37i grouped in the same cluster in all the screening tests. They belong to same genetic group, Criollo and their relationship is augmented by common features, such as their characteristic pink cotyledon colour which sets them apart from the other clones. This agrees with the observations of Bartley (1964) who also reported similar cotyledon colour of genetically similar genotypes of cocoa. The LCTEEN clones were collected from Ecuador in an area bounded by the Lower amazon region.
CONCLUSION
Breeders of cocoa often consider resistance to diseases in addition to yield and interest in how improved germplasm relates to resistance to diseases. Results from this study can aid germplasm development efforts in addressing those perspectives. Understanding how screening methods used in breeding for resistance to Phytophthora species cluster different genetic groups would increase the likelihood of selecting resistant materials resulting in improved germplasm.
The resistant Trinitario clones from Upper amazon and Forastero clones from Lower amazon, (ICS1 and T85/799) and (LAF 1, SCA6, EET 59H, PA 107), respectively might have similar resistance factors and therefore may be good parents for breeding against black pod disease resistance in cocoa.
ACKNOWLEDGMENTS
We thank the Common Fund for Commodities (CFC) for financial support. Our sincere gratitude goes to the technical staff of Plant Pathology and Plant Breeding Divisions, CRIG, for help in this study. This research article was published with kind permission of the director of Cocoa Research Institute of Ghana.
REFERENCES
- Adomako, B., 2006. Combining ability analysis of black pod disease incidence in cocoa genotypes in Ghana. Trop. Sci., 46: 201-204.
CrossRef - Adomako, B., 2007. Causes and extent of yield losses in cocoa progenies. Trop. Sci., 47: 22-25.
CrossRef - Afanasenko, O., N. Mironenko, O. Filatova, D. Kopahnke, I. Kramer and F. Ordon, 2007. Genetics of host-pathogen interactions in the Pyrenophorateres f. teres (net form)-barley (Hordeumvulgare) pathosystem. Eur. J. Plant Pathol., 117: 267-280.
CrossRef - Ahmed, H.U., K.F. Chang, S.F. Hwang, B.D. Gossen, R.J. Howardand and T.D. Warkentin, 2006. Components of disease resistance in desi and kabuli chickpea varieties against ascochyta blight. Plant Pathol. J., 5: 336-342.
CrossRefDirect Link - Akaza, M.J., J.A.K.N. Goran, S.P.A.N. Guetta, B.I. Kebe, G.M. Tahi and A. Sangare, 2009. Resistance to Phytophthora palmivora (Butler) butler assessed on leaf discs of cacao (Theobroma cacao L.) hybrid trees. Asian J. Plant Pathol., 3: 106-118.
CrossRefDirect Link - Chaveerach, A., R. Sudmoon, T. Tanee, P. Mokkamul and A. Tanomtong, 2007. Genetic relationships in a population of Nelumbo nucifera Gaertn. (Nelumbonaceae). J. Biol. Sci., 7: 1388-1393.
CrossRefDirect Link - Broich, S.L. and R.G. Palmer, 1981. Evolutionary studies of the soybean: The frequency and distribution of alleles among collections of Glycine max and G. soja of various origin. Euphytica, 30: 55-64.
CrossRef - Djocgoue, P.F., T. Boudjeko, D.J. Nankeu, M.I.B. Efombagn, S. Nyasse and D.N. Omokolo, 2006. Comparative assessment of the resistance of cocoa (Theobroma cacao L.) progenies from SNK10 x SNK413; ICS84 x ICS95 to Phytophthora megakarya in cameroon by measuring size of necrotic lesion along the midrib. Plant Pathol. J., 5: 329-335.
CrossRefDirect Link - Gizlice, Z., T.E. Carter and J.W. Burton, 1994. Genetic base for North American public soybean cultivars released between 1947 and 1988. Crop Sci., 34: 1143-1151.
Direct Link - Greathouse, D.C., W.M. Laetsch and B.O. Phinney, 1971. The shoot-growth rhythm of a tropical tree (Theobroma cacao L.). Am. J. Bot., 58: 281-286.
Direct Link - Hymowitz, T., 1970. On the domestication of the soybean. Econ. Bot., 23: 408-421.
CrossRefDirect Link - Iwaro, A.D., 1996. Strategies of variety improvement in cocoa with emphasis on durable disease resistance. An external review prepared for INGENIC. Proceedings of the International Workshop on the Contribution of Disease Resistance to Cocoa Variety Improvement, Nov. 24-26, Salvador, Bahia, Brazil, pp: 23-32.
- Iwaro, A.D., T.N. Screenivasan and P. Umaharan, 1997. Phytophthora resistance in cacao (Theobroma cacao): Influence of pod morphological characteristics. Plant Pathol., 46: 557-565.
CrossRef - Mohammadi, S.A. and B.M. Prasanna, 2003. Analysis of genetic diversity in crop plants-salient statistical tools and considerations. Crop Sci., 43: 1235-1248.
CrossRefDirect Link - Nyadanu, D., M.K. Assuah, B. Adomako, Y.P. Asiama, I.Y. Opoku and Y. Adu-Ampomah, 2009. Efficacy of screening methods used in breeding for black pod disease resistance varieties in cocoa. Afr. Crop Sci. J., 17: 175-186.
CrossRefDirect Link - Nyasse, S., C. Cilas, C. Herail and G. Blaha, 1995. Leaf inoculation as an early screening test for cocoa (Theobroma cacao L.) resistance to Phytophthora black pod disease. Crop Prot., 14: 657-663.
CrossRefDirect Link - Peneder, M., 1995. Cluster techniques as a method to analyze industrial competitiveness. Int. Adv. Econ. Res., 1: 295-303.
CrossRef - Reed, D.H. and R. Frankham, 2003. Correlation between fitness and genetic diversity. Conserv. Biol., 17: 230-237.
CrossRefDirect Link - Robinson, J. and M. Jalli, 1996. Diversity among Finnish net blotch isolates and resistance in barley. Euphytica, 92: 81-87.
CrossRef - Tahi, M., I. Kebe, A.B. Eskes, S. Ouattara, A. Sangare and F. Mondeil, 2000. Rapid screening of cacao genotypes for field resistance to Phytophthora palmivora using leaves, twigs and roots. Eur. J. Plant Pathol., 106: 87-94.
CrossRefDirect Link - Tanksley, S.D. and S.R. McCouch, 1997. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science, 277: 1063-1066.
CrossRefPubMedDirect Link - Thresh, J.M., G.K. Owusu, A. Boamah and G. Lockwood, 1988. Ghanaian cocoa varieties and swollen shoot virus. Crop Prot., 7: 219-231.
CrossRef - Ward, Jr. J.H., 1963. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc., 58: 236-244.
CrossRefDirect Link - Tan, G.Y. and W.K. Tan, 1990. Additive inheritance of resistance to pod ret caused by Phytophthora palmivora in cocoa. Theor. Applied Genet., 80: 258-264.
CrossRef - Ramdani, M., O. Rached, T. Lograda and A. Aggoun, 2007. Genetic diversity in foliar terpinoids among natural populations of Cupressus dupreziana in Tassili n'Ajjer (Algeria). Asian J. Plant Sci., 6: 1211-1216.
CrossRefDirect Link - Iwaro, A.D., T.N. Sreenivasan and P. Umaharan, 1997. Foliar resistance to Phytophthora palmivora as an indicator of pod resistance in Theobroma cacao. Plant Dis., 81: 619-624.
CrossRefDirect Link