
B. M. Gichimu
Coffee Research Foundation, P.O. Box 4-00232, Ruiru, Kenya
C. O. Omondi
Coffee Research Foundation, P.O. Box 4-00232, Ruiru, Kenya
International Journal of Plant Breeding and Genetics
Year: 2010 | Volume: 4 | Issue: 4 | Page No.: 238-246







ABSTRACT
Five new lines of Arabica coffee and two existing commercial cultivars planted in the field were characterized morphologically. The objective of this study was to determine the level of morphological variation within and between new and existing coffee varieties in Kenya. Morphological characterization was done using a 25 character coffee descriptors. Both quantitative and qualitative morphological traits were recorded and the binary data were subjected to cluster and principal component analysis. Morphological variation among the genotypes was less than 25 percent. Four main groups were formed when the similarity index was considered for clustering. The improved cultivar, Ruiru 11 and the traditional cultivar SL28 were clustered alone in separate clusters, while two new lines CR8 and CR30 clustered together. The other three new lines CR22, CR23 and CR27 also clustered together in shared sub-clusters. As expected there was very little morphological variation within the varieties. In the principal component analysis, Ruiru 11 separated clearly from the rest and was located on the lower side of the PCA graph. SL28 was grouped with the new lines on the upper part of the PCA graph. The first two principal components accounted for a cumulative variance of 60%. The study demonstrated low morphological variation and hence low genetic variation among the varieties and emphasized the need to broaden the genetic base of Arabica coffee in Kenya. In addition, the study demonstrated minimal morphological variation within the varieties indicative of high genetic consistency.
PDF Abstract XML References Citation
Received: April 27, 2010;
Accepted: June 15, 2010;
Published: July 05, 2010
How to cite this article
B. M. Gichimu and C. O. Omondi, 2010. Morphological Characterization of Five Newly Developed Lines of Arabica Coffee as Compared to Commercial Cultivars in Kenya. International Journal of Plant Breeding and Genetics, 4: 238-246.
DOI: 10.3923/ijpbg.2010.238.246
URL: https://scialert.net/abstract/?doi=ijpbg.2010.238.246
DOI: 10.3923/ijpbg.2010.238.246
URL: https://scialert.net/abstract/?doi=ijpbg.2010.238.246
INTRODUCTION
Coffee belongs to the genus Coffea in the Rubiaceae family which contains some 640 genera and 10000 species. It is a biologically and morphologically diverse family consisting of varied life forms ranging from tiny herbs, epiphytes, lianas, shrubs to tall trees (Bremer, 1996). The genus Coffea consist of approximately 105 taxa and is distinguished from a closely related genus, Psilanthus, based on flowering and flower characteristics (Kumar et al., 2008). All Coffea species are native to the inter-tropical forest of Africa and Madagascar, while species belonging to the genus Psilanthus originate from either Asia or Africa. The genus Coffea L. has been reorganized into two subgenera: Coffea and Paracoffea (Bridson, 1987). Particular attention has been paid to the subgenus Coffea which includes two cultivated species of economic importance, Coffea arabica L. and Coffea canephora Pierre (Kumar et al., 2008).
C. arabica is tetraploid (2n = 4x = 44) and is self-fertile while other Coffea species are diploid 2n = 2x = 22) and generally self-incompatible (Masumbuko et al., 2003). C. arabica has two distinct botanical varieties C. arabica var. arabica (usually called Typica) and C. arabica var. bourbon (usually called Bourbon) (Hue, 2005). Historical data indicate that the Typica genetic base originated from a single plant from Indonesia which was subsequently cultivated in the Amsterdam botanical garden in the early 18th century, around 1715 (Hue, 2005). The Bourbon genetic base originated from a few coffee trees introduced from Mocha (Yemen) to the Bourbon Island (now La Reunion) at about the same time (Hue, 2005). The narrow geographic origin of C. arabica, along with its self-fertilising nature and the historical or selective bottlenecks in its agricultural adoption, have resulted in low genetic diversity of C. arabica varieties cultivated around the world (Chaparro et al., 2004). Another possibility could be a drastic loss of genetic diversity during glaciation phases of the quaternary period (Lashermes et al., 1993).
The most popular C. arabica cultivars in Kenya are SL28 and Ruiru 11. The cultivar SL28 is a single tree selection from Tanganyika Drought Resistant, a variety selected in the northern province of Tanzania in 1931. It is a tall statured cultivar with long internodes. Ruiru 11 is a short and compact F1 hybrid cultivar derived from a cross between selected female and male parents (Omondi et al., 2001). Coffee Research Foundation has recently developed five new lines of Arabica coffee code named CR8, CR22, CR23, CR27 and CR30. The five new lines are outstanding selections from a multiple cross programme involving CBD resistant donor parents such as Rume Sudan (R gene), Hibrido de Timor (T gene), K7 (k gene) and the high yielding, good quality but susceptible cultivars such as N39, SL28, 34 and 4 (Gichimu and Omondi, 2010). Their unique features include tall stature, true breeding and resistance to the two major fungal diseases of coffee namely Coffee Berry Disease (CBD) and Coffee Leaf Rust (CLR). They are also high yielding with good bean and liquor quality that compares to Ruiru 11 and SL28 (Gichimu and Omondi, 2010).
Over the years, coffee breeders have tried to widen the genetic base of Arabica coffee by having more introductions and undertaking hybridisation programmes to create variability (Lashermes et al., 1999). As new coffee varieties are continuosly being developed through hybridization, there is need to determine the level and sources of morphological variation within and between new and existing coffee varieties. Genetic consistency within varieties is essential to quality assurance for any agricultural product. It is believed that morphological variability in coffee plantations is adverse to the product quality (Hue, 2005). While the Kenyan coffee industry targets high quality coffee, the observed morphological variation within coffee varieties could lower the quality. Therefore there is a need therefore to determine the causes of morphological variability within Kenyan coffee varieties. Due to some limitations, the ecological tests such as soil, sunlight and wind were not examined in this study. However, because the test plants were within a limited geographic area with nearly homogenous soil type, ecological contribution was assumed to be minimal.
MATERIALS AND METHODS
Study Site
The field trial was carried out at Tatu Estate in Ruiru District, Kenya from 2005 to 2009. The site lies within the upper Midland 2 agro-ecological zone (UM 2) at latitude 01°06’S and longitude 36°45’E and is approximately 1620 m above the sea level. The area receives a bimodal mean annual rainfall of 1063 mm and the mean annual temperature is 19°C (minimum 12.8°C and maximum 25.2°C). The soils are classified as a complex of humic nitisols and plinthic ferrasols. They are welldrained, deep reddish brown, slightly friable clays with murram sections occasionally interrupting. The soil pH ranges between 5 and 6 (Jaetzold and Schimidt, 1983).
Test Materials
The test materials included five advanced breeder’s lines coded CR8, 22, 23, 27 and 30 which were evaluated alongside two commercial Arabica cultivars, SL28 and Ruiru 11 as check cultivars. The five breeder’s lines have been developed by Coffee Research Foundation in Ruiru, Kenya. They are true breeding with tall stature and have been tested both in the lab and in the field and proven to be resistant to two major fungal diseases of coffee namely Coffee Berry Disease (CBD) and Coffee Leaf Rust (CLR).
Experimental Layout
The site was laid out in a Randomized Complete Block Design (RCBD) with twenty trees per plot planted on a spacing of 2x1.5 M and replicated three times. Field establishment was done in April/May 2005 and morphological data scored in October 2009.
Data Collection
Data were collected on morphological (both qualitative and quantitative) characters using a 25 character descriptors adopted from UPOV. Each qualitative descriptor was scored by observing twenty tagged trees per genotype taking six to seven plants from every block (replicate) and then calculating the mode to get an overall figure per replicate. Quantitative descriptors were taken as the mean value of three measurements made on six to seven trees per replicate and then calculating the mean to get an overall figure per replicate.
Data Analysis
The data was organized into a matrix and subjected to cluster analysis using R Statistical Software (Venables et al., 2006). Variables were segregated into discrete factors (e.g., Fruit colour - light red, pink, yellow); rank-ordered factors (e.g., fruit size - very small, small, medium, large and very large); integers (e.g., number of flowers per inflorescence) and numerical variable (e.g., average internode length). The clustering was done using DAISY (dissimilarity matrix calculation) function and unweighted pair-group method with arithmetic average [UPGMA] (Venables et al., 2006). Quantitative data were subjected to ANOVA using SAS version 9.1. Students-Newman-Keuls (SNK5%) was used to separate the means. The procedure PRINCOMP was then used to perform a principle component (PC) analysis using the quantitative variables. In this procedure, first a similarity matrix was calculated and was used to calculate eigen values and scores for the accessions. The accessions were then plotted on two dimensions using the first two principle components (PC1 and PC2).
RESULTS
Variation in qualitative characters is presented in Table 1; there were minimal variations among all genotypes in most of the qualitative characters such as leaf shape, anthocyanin colouration and undulation of the margin, depth of the secondary veins, leaf domatia, fruit shape, fruit colour and adherence to the branch. Significant variations were found in plant shape and fruit size. CR8, 30 and SL28 were conical in shape while the rest were cylindrical. The fruit size ranged medium for CR27 to large for CR22, 23, SL28 and Ruiru 11 and very large for CR8 and CR30. The new lines were morphologically very similar to SL28 especially for plant height, canopy diameter and intensity of ramification. They were tall with large canopy diameter and medium intensity of ramification unlike Ruiru 11 which was short with medium canopy diameter and strong intensity of ramification.
| Table 1: | Variation of qualitative characters in the seven coffee genotypes |
 | |
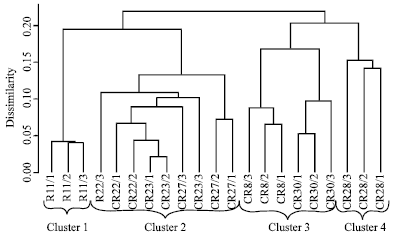 | |
| Fig. 1: | Cluster dendrogram illustrating morphological diversity between the seven coffee genotypes characterized using 24 morphological descriptors |
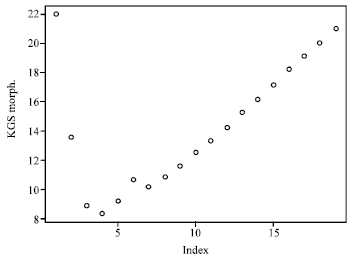 | |
| Fig. 2: | Kelley-Gardner-Sutcliffe (KGS) penalty function showing the similarity index for clustering |
Results of the cluster analysis are presented in Fig. 1. Degree of similarity varied from 25 to 20% and four main groups were formed when the similarity index was considered for clustering (Fig. 2). The improved cultivar, Ruiru 11 and the traditional cultivar SL28 were clustered alone in separate clusters, while two new lines CR8 and CR30 clustered together. The other three new lines CR22, 23 and 27 also clustered together in shared sub-clusters. The R command g clus was used to reorder the genotypes within a cluster keeping them contiguous to each other and therefore Ruiru 11 and SL28 were the most distantly related.
| Table 2: | Variation of quantitative characters in the seven coffee genotypes |
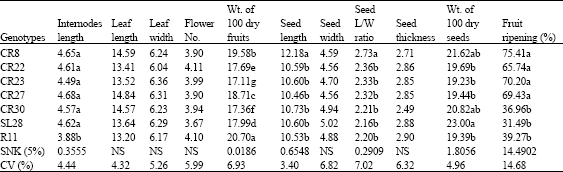 | |
| NB: Fruit ripening was scored when some of the tagged branches had attained 100% ripening | |
| Table 3: | The first two Principle Components (PC) of the morphological characters |
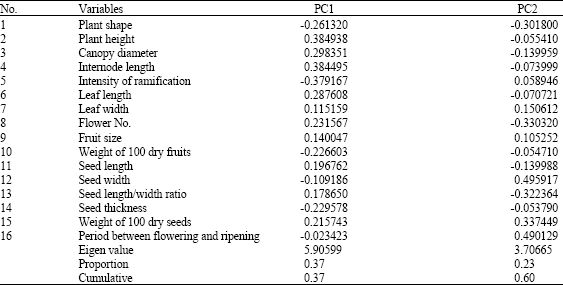 | |
The cultivar Ruiru 11 was found to be the most uniform recording intra-variation of less than 5%. On the other hand, SL28 was the most variable cultivar with an intra-variation of close to 15%. The new lines portrayed an intra-variation of less than 10%.
Quantitative morphological data are presented in Table 2, all the test materials were not significantly different in leaf length, leaf width, flower number, seed width and seed thickness. However, significant variations were obtained in internode length, weight of 100 dry fruits, seed length, seed lenth/width ratio, weight of 100 dry seeds and period between flowering and harvesting. Ruiru 11 had significantly shorter internode length than the rest of the test materials. On the other hand, CR8 had a bigger seed size as evident from its significantly bigger seed length and seed length/width ratio as compared to the rest of the genotypes. All the test materials were significantly different from each other in weight of 100 dry fruits while SL28 recorded significantly more weight of 100 dry seeds than Ruiru 11, CR22, CR 23 and CR27 though similar to CR8 and CR30. SL28, Ruiru 11 and CR30 took significantly longer time between flowering and harvesting than CR8, 22, 23 and 27.
Results of the principle component analyses indicated that the first two PCS explained 37 and 23% (a total of 60%) of the total variation (Table 3). The most principle component was weight of 100 dry seeds followed by the period between flowering and ripening, plant height and internode length in that order (Table 3). Plant height and internode length contributed most to PC1 while seed width and period between flowering and ripening contributed most to PC2.
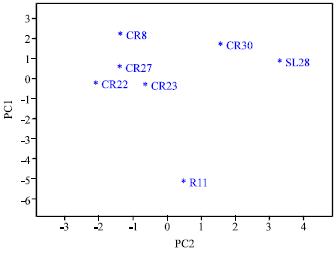 | |
| Fig. 3: | Diversity among genotypes as determined by morphological variables |
The traits leaf shape, anthocyanin colouration, undulation of the margin, depth of the secondary veins, presence or absence of leaf domatia, fruit shape, fruit colour and adherence to the branch did not contribute at all to both PC1 and 2.
The two dimensional presentation of the seven genotypes is presented in Fig. 3. Improved commercial cultivar, Ruiru 11, separated clearly from other genotypes and was located on the lower part of the PCA graph (Fig. 3). On the other hand, the traditional commercial cultivar, SL28 and the five advanced breeder’s lines were located on the upper part. Among the advanced breeder’s lines, CR22, 23 and 27 were plotted closely together, a distance from CR8 and 30 which were plotted on the uppermost part of the PCA graph. CR30 was plotted closer to SL28 than other advanced breeder’s lines.
DISCUSSION
Genetic variation in Arabica coffee has previously been characterised using morphological and yield-related traits and phylogenetic relationships established (Lashermes et al., 1996). Morphological markers are a classical method to distinguish variation based on the observation of the external morphological differences such as the size and shape of the leaf and of the plant form, the colour of the shoot tip, the characteristics of the fruit, the angle of branching and the length of the internodes (De Vienne et al., 2003). In this study, the cluster analysis depicted less than 25% morphological variation between the test materials indicating a narrow genetic base. This contradicted the report of Kumar et al. (2008) that coffee trees differ greatly in morphology, size and ecological adaptation. Ruiru 11 and SL28 were the most distantly related. The observed difference could be attributed to hybridization programme used in the development of Ruiru 11 unlike in SL28 where single tree selection procedures were used.
There was a high degree of similarity in qualitative traits of SL28 and the five new lines. The principle component analysis also produced SL28 as being closely related to the five new lines than to Ruiru 11. However, considering that the five new lines were selected from the male parents of Ruiru 11, it was expected that the new lines would be more genetically similar to Ruiru 11 than to SL28. The otherwise results could be attributed to inefficiency of morphological markers in assessing genetic variation. This concurs to the report of De Vienne et al. (2003) that assessing genetic variation with morphological markers can be inefficient as they are generally dominant traits, they often exhibit epistatic interactions with other genetic traits and can also be influenced by the environment. Lashermes et al. (1996) reported that genetic factors are more accurately tested by molecular markers. The observed similarity was also attributed to the single tree selection procedures that were used to develop these varieties. Agwanda et al. (1997) also reported that single tree selection procedures used to develop most Arabica coffee cultivars have contributed to high level of uniformity among them.
The divergence of Ruiru 11 from the other varieties studied could have resulted from its female parent Catimor, which has also been shown (Agwanda et al., 1997) as being genetically divergent from other Arabica genotypes owing to the interspecific origin of one of its progenitor, Timor Hybrid. Ruiru 11 being an F1 hybrid of the variety Catimor, as the female parent and outstanding male selections most of which has Timor Hybrid in their pedigree must contain a considerable amount of Timor Hybrid genes. In addition all Ruiru 11 male progenitors contain the semi-wild variety, Rume Sudan in their parentage which further accentuates its divergence from other varieties. Agwanda et al. (1997) reported that the use of semi-wild varieties such as Rume Sudan, or interspecific hybrids such as Catimor, as resistance donors results into the associated introgression of other un-targeted and agronomically undesirable genes. Silveira et al. (2003) also reported that coffee genotypes derived from Timor Hybrid demonstrate notable genetic diversity. Although both Rume Sudan and Timor Hybrid are contained in the pedigree of the five new lines, the magnitude of their genes may be smaller than in Ruiru 11.
As expected there was very little morphological variation within varieties. The mean dissimilarity values between varieties were higher than within varieties which underscores the low variability and the inbreeding nature of cultivated Arabica coffee. The results concurs with those of Anthony et al. (2001), who also demonstrated low genetic variation within Arabica coffee genotypes. Masumbuko and Bryngelsson (2006) also found similar results when comparing diploid coffee species and cultivated Coffea arabica L. from Tanzania. The low genetic variability within varieties further strengthened the evidence of the narrow genetic base of Arabic coffee. The observed minimal intra-variation within varieties was attributed to ecological instability as the trees were less than five years old. This is in line with the report of Hue (2005) that variability within varieties may result from environmental and/or management factors such as mis-labelling or mis-planting. However, other hypothesis, such as gene introgression from a gene pool alien to C. arabica, cannot be excluded.
CONCLUSION
The study demonstrated minimal morphological variation among the cultivars that were tested indicating low genetic variation and narrow genetic base. There is therefore need to widen the genetic base of Arabica coffee in Kenya by having more introductions, initiate hybridisation programmes to create variability and use of diploid species as a source of desirable genes. The study also demonstrated minimal morphological variation within varieties indicative of high genetic consistency within Kenyan Arabica coffee varieties.
ACKNOWLEDGMENT
This research was co-financed by Coffee Research Foundation (CRF) and the Common Fund for Commodities (CFC) through the Coffee Leaf Rust Project (CFC/ICO/40). Additional financial support was provided the European Union through the Quality Coffee Production and Commercialization Programme (QCPCP). Thanks are due to Messrs S.M. Njeruh and M.M. Musembi of Plant Breeding section who were responsibile for the management of experimental site and data collection. This work is published with the permission of the Director of Research, CRF, Kenya.
REFERENCES
- Agwanda, C.O, P. Lashermes, P. Trouslot, M.C. Combes and A. Charrier, 1997. Identification of RAPD markers for resistance to coffee berry disease, Colletotrichum kahawae, in Arabica coffee. Euphytica, 97: 241-248.
CrossRefDirect Link - Anthony, F., B. Bertrand, O. Quiros, A. Wilches, P. Lashermes, J. Berthaud and A. Charrier, 2001. Genetic diversity of wild coffee (Coffea arabica L.) using molecular markers. Euphytica, 118: 53-65.
CrossRef - Bremer, B., 1996. Combined and separate analyses of morphological and molecular data in the plant family Rubiaceae. Cladistics, 12: 21-40.
CrossRef - Chaparro, A.P., M.A. Cristancho, H.A. Cortina and A.L. Gaitan, 2004. Genetic variability ofCoffea arabica L. accessions from Ethiopia evaluated with RAPDs. Genet. Resour. Crop Evol., 51: 291-297.
CrossRef - Gichimu, B.M. and C.O. Omondi, 2010. Early performance of five newly developed lines of Arabica Coffee under varying environment and spacing in Kenya. Agric. Biol. J. North Am., 1: 32-39.
Direct Link - Lashermes, P., J. Cros, P. Marmey and A. Charrier, 1993. Use of random amplified DNA markers to analyse genetic variability and relationships of Coffea species. Genet. Resour. Crop Evol., 40: 91-99.
CrossRefDirect Link - Lashermes, P., M.C. Combes, J. Robert, P. Trouslot, A. D'Hont, F. Anthony and A. Charrier, 1999. Molecular characterisation and origin of the Coffea arabica L. genome. Mol. Gen. Genet., 261: 259-266.
PubMed - Lashermes, P., P. Trouslot, F. Anthony, M.C. Combes and A. Charrier, 1996. Genetic diversity for RAPD markers between cultivated and wild accessions of Coffea arabica. Euphytica, 87: 59-64.
CrossRef - Masumbuko, L.I., T. Bryngelsson, E.E. Mneney and B. Salomon, 2003. Genetic diversity in Tanzanian Arabica coffee using random amplified polymorphic DNA (RAPD) markers. Hereditas, 139: 56-63.
CrossRefDirect Link - Masumbuko, L.I. and T. Bryngelsson, 2006. Inter simple sequence repeat (ISSR) analysis of diploid coffee species and cultivated Coffea arabica L. from Tanzania. Genet. Resourc. Crop Evolut., 53: 357-366.
CrossRef - Omondi C.O., P.O. Ayiecho, A.W. Mwang'ombe and H. Hindorf, 2001. Resistance of Coffea arabica cv. Ruiru 11 tested with different isolates of Colletotrichum kahawae, the causal agent of coffee berry disease. Euphytica, 121: 19-24.
Direct Link - Silveira, S.R., P.M. Ruas, C.F. Ruas, T. Sera, V.P. Carvalho and A.S.G. Coelho, 2003. Assessment of genetic variability within and among coffee progenies and cultivars using RAPD markers. Genet. Mol. Biol., 26: 329-336.
Direct Link