
Manal M. Hefny
Not Available
International Journal of Plant Breeding and Genetics
Year: 2007 | Volume: 1 | Issue: 2 | Page No.: 54-66







ABSTRACT
Sixteen yellow maize inbred lines and their twelve single crosses, in addition to one check hybrid, single cross Pioneer 3062, were tested under two levels of nitrogen fertilizer: low (70 kg N f-1) and high (140 kg N f-1). The experiment was carried out at the experimental farm of Suez Canal University, Ismailia, Egypt to assess the genetic parameters and heterosis for nitrogen use efficiency related traits at each N level. In general, genetic variances and heritability estimates were higher at high N fertilizer than under stress conditions for inbred lines and hybrids. Grain yield traits of inbreds showed low heritabilities at both N fertilizer rates, except for 100-grain weight, however the expected genetic improvement was high at stress than under optimal conditions. On the contrary, the genetic improvement for nitrogen related-traits would be effective if selection is practiced under optimal fertilization except for nitrogen use efficiency for biomass production. Efficiency of the expected genetic improvement would increase for hybrid grain yield, N-uptake and nitrogen use efficiency for grain production if selection is carried out at high N levels. Genetic improvement could be practiced for inbred and hybrid for dry matter production under stress N environments. The cross (8x5) was the one demonstrated a attractive mid-parent heterosis for grain yield, yield plant-1, 100-grain weight, nitrogen use efficiency for grain production and biomass production, nitrogen harvest index, high protein, in addition it was early in days to 50% tasselling and silking. Midparent-hybrid correlation demonstrated high and positive coefficients at low N for all measured traits, except total dry matter, 100-grain weight and N-uptake, which is a predictor for hybrid performance under stress conditions based on inbred per se.
PDF Abstract XML References Citation
How to cite this article
Manal M. Hefny, 2007. Estimation of Quantitative Genetic Parameters for Nitrogen Use Efficiency in Maize under Two Nitrogen Rates. International Journal of Plant Breeding and Genetics, 1: 54-66.
DOI: 10.3923/ijpbg.2007.54.66
URL: https://scialert.net/abstract/?doi=ijpbg.2007.54.66
DOI: 10.3923/ijpbg.2007.54.66
URL: https://scialert.net/abstract/?doi=ijpbg.2007.54.66
INTRODUCTION
Drought and low soil fertility are the two among the most important a biotic stresses threatening maize production, limiting food security and economic growth (Bänziger and Diallo, 2001). The incidence of stress may increase, due partly to global climate changes, declines in soil organic matter, reducing soil fertility and water holding capacity (Bänziger et al., 2000).
Breeding for a biotic stress environments has been done for a number of crops like oat (Atlin and Frey, 1989), barley (Ceccarelli, 1987), alfalfa (Antolin et al., 1995), wheat (Ud-Din et al., 1992) and maize (Bolaños and Edmeades, 1993). One approach to reducing the impact of N deficiency on maize production may be to, select cultivars that are superior in the utilization of available N, either due to enhanced uptake capacity or because of more efficient use of absorbed N in grain production (Lafitte and Edmeades, 1994).
So, breeders need information about parameters, such as heritability, genetic and environmental variances under N supply. They also need to know the dynamics of these parameters under N increasing. In addition, adequate genetic variability for the traits that determine N efficiency is compulsory. Genotypic variation was reported for N uptake and/or N utilization efficiency in bread wheat (Ortiz-Monasterio et al., 1997) and maize (Moll et al., 1982). Despite the noticeable genotypic variation, May et al. (1991) recorded strong influence of environment on N use traits and concluded that it would be difficult to modify them by breeding.
While Rosielle and Hamblin (1981) stated that most crop breeding is conducted under high yielding conditions where heritability and genotypic variance for grain yield and therefore potential selection gains, are high. In contrast, Blum (1988) suggested the effectiveness of selection for yield in the target environment rather than selection for yield potential alone. However, such environments are not favored by maize breeders due to increased environmental variability as soil fertility declines resulting in a decline in heritability for grain yield (Lafitte and Edmeades, 1994). Bänziger and Lafitte (1997) evaluated maize germplasm adapted to lowland tropics under high and low N conditions. They mentioned that genotypic variance for grain yield under low N was about one third of the average genotypic variance for grain yield under high N, but the average error variance was similar at both low and high N levels. They found that among low N experiments, genotypic variance and error variance for grain yield tended to decrease with increasing relative yield reduction under low N, while heritability did not change.
Heritability for grain yield averaged 65% both under high N and low N environments (Presterl et al., 2003). Atlin and Frey (1989) found the heritability of grain yield to be slightly greater in high N than in low N environments. Bänziger and Lafitte (1997) further reported that broad sense heritabilities of grain yield under low N were smaller than under high N. They recorded a positive genetic correlation between grain yield under low and high N. Lafitte and Edmeades (1994) evaluated different cycles of full-sib recurrent selection under low and high N conditions. They reported that realized heritability was generally larger for yield under low N than for yield under high N and that all traits evaluated had larger values of heritability when measured in cycle 2 versus cycle 0 of recurrent selection.
Heterosis is important in maize breeding and is dependent on level of dominance and differences in gene frequency. The manifestation of heterosis depends on genetic divergence of the two parental varieties (Hallauer and Miranda, 1988). Le Gouis and Pluchard (1996) in their experiment on a set of wheat hybrids at low and high N, reported a higher best- parent heterosis for grain yield at low N level (9.9%*) than at the high N level (1.1%ns)
There is little information in the literature on the estimation of genetic variance components and genetic advance under selection for yield and nitrogen use efficiency-related traits in corn at different levels of nitrogen fertilizer. Also information about heterosis for yield and nitrogen related traits under different N levels in maize are seldom.
The present study comprises three objectives, (1) estimate the genetic and environmental variances for yield and nitrogen use efficiency related-traits in an introduced set of yellow inbred lines and their crosses, (2) estimate the degree of heterosis on the basis of the deviation of cross mean from mid-parent and the check variety and (3) estimate the expected genetic advance within each nitrogen environment. The above-mentioned parameters are of major concern for breeders for further improvement of nitrogen use efficiency.
MATERIALS AND METHODS
The field experiment for this study was described in Hefny and Aly (2007) and only procedures essential for understanding this study are given below. The experiment consisted of sixteen exoticyellow maize inbred lines crossed in a hierarchal mating system to produce twelve single crosses.
| Table: 1: | List of variables measured on 16 inbred lines and their 12 single crosses and one check S.C. Pioneer 3062, cultivated at two levels of N fertilizer |
 | |
The single cross Pioneer 3062 was included for comparison. The experiment was cropped at the experimental farm of Suez Canal University, Ismailia, Egypt. during the 2006 summer season under two levels of N fertilizers (70 and 140 kg N f-1). A split-plot design with three replications was used, with fertility treatments as main-plots and genotypes as sub-plots. Each sub plot was made up of one row of 3 m long and 0.25 within row. Data were recorded on the following variables (Table 1).
Statistical Analysis
As the main objective of the present study is to quantify the effect of applied N on the estimation of genetic and environmental components, heritability, genetic advance and heterosis, each level of N fertilizer were statistically analyzed.
The variance components and broad-sense heritability were estimated from ANOVA table calculated for each N level according to Snedecor and Cochran (1980):
| σ2 g = (Mg-Me)/r | |
| σ2 e = Me/r | |
| σ2 p = Mg/r |
The standard error of genetic variabce was calculated (Becker, 1975) as:
| Where: | ||
| Msi | = | The mean square of effect i used to estimate variance component (g), |
| fi | = | The degrees of freedom for Msi, |
| k | = | The coefficient for σ2g in the expected mean squares. |
Broad sense heritability (h2) was estimated as
h2b = σ2 g / σ2 p
The standard error for heritability was calculated (Becker, 1975) as,
SE (h2) = [SE (σ2g)]/σ2p
| Where: | ||
| σ2p | = | The phenotypic variance. |
Expected Genetic Advance (GA) was calculated using the formula of Allard (1960):
GA = (k) (h2) (σph),
| Where: | ||
| h2 and σph | = | Heritability and square root of phenotypic variance. |
| k | = | Selection differential that varies depending on the selection intensity. i.c., k on considered 2.06 and is the value of 5% selection intensity. |
Heterosis was expressed as the percentage deviation from MP and the S.C.3062. For each hybrid and each character, the difference between the hybrid and the mean of its two parents was computed. A t-test was used to test whether the mean of these differences was different from 0, then if mid-parent heterosis was significant (Le Gouis et al., 2002).
All statistical analyses were performed with GenStat Software Package, Release 4.24.
RESULTS AND DISCUSSION
Variance Components and Heritability Estimates
Variance components, genetic and environmental variances and heritability estimated for inbred lines and crosses at both N fertilizer levels are shown in Table 2. Ten characters recorded high genetic variance values under optimal N fertilizer compared to those under low N application. Regarding crosses, also the variance component due to genetic portion was higher for nine characters at optimal N fertilized compared to those for low N application. There was a general trend towards the higher average heritability value at optimal compared with low N fertilizer (1.11 and 1.10%, respectively). The low heritability values at low N relative to optimal condition is relate to the remarkable decrease in genetic variance more than environmental variance for inbred lines and their crosses, respectively. The previous results are confirmed by those obtained by Laperche et al. (2006), who recorded a general high heritability values for grain yield, HI and grain protein yield under N+ than under N- applications in wheat populations. In contrast Presterl et al. (2003) recorded an average of 43% increases in genetic variance at LN compared with HN when estimated for 21 field experiments with different breeding materials of corn selected under different N conditions. However, they added, heritability estimates did not show higher values at LN due to increasing environmental variance. Under optimal conditions (soil fertility), the genotypes reaction would not differ as much as would under stressed conditions, thus genetic variance and then heritability should increase under stressed cultivation (Laperche et al., 2006).
In the current study the opposite was obtained, due to two combined explanations. One is the decreased in general mean of the measured traits when exposed to stress environments and consequently the genetic variance reduction (Laperche et al., 2006). The second explanation is the different patterns genetic and environmental variances expressed: Increase of σ2g and σ2e (Sinebo et al., 2004); or decrease in σ2g and increase in σ2e (Brancourt-Hulmel et al., 2005; Makumbi, 2005). Also, Laperche et al. (2006) attributed the contradiction of such results to the characteristics of the population under study.
| Table 2: | Variance components, heritability and genetic advance for yield and nitrogen use efficiency related-traits estimated at two levels of nitrogen fertilizers (70 and 140 kg N f-1) for sixteen inbred line and F1 crosses |
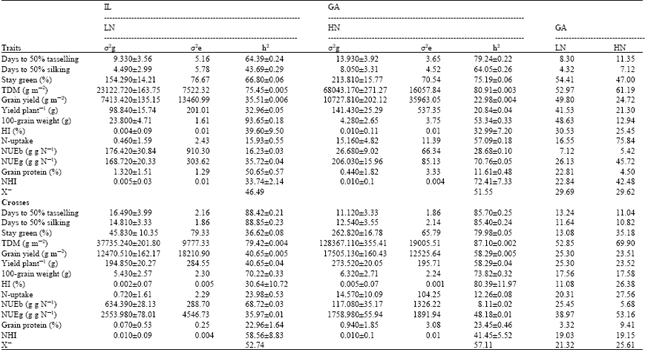 | |
| ±: SE | |
One explanation is that, the population’s complexity which is composed of lines originating from different sources (present study) should have resulted in, theoretically, higher σ2g and consequently higher h2 values. But they commented that, the impact of breeding is the main reason for genetic variance reduction, Presterl et al. (2003) different combinations from S2, S4 and S6 maize inbred lines, but in the current study S7 and S8 inbred lines were used.
When focus on the measured traits, generally, heritabilities were higher for the most traits at HN supplies for both parents and crosses. Although inbred lines showed decreased genetic variance with N stress intensity compared with HN, heritability estimates for yield traits demonstrated low values at both N levels, but still higher at low compared with high N, which is a consequence of higher environmental variance at HN compared with LN. Inbred lines recorded high to moderate h2 values for; flowering traits, stay green, dry matter yield and nitrogen traits (N-uptake, NUEb, NUEg and NHI) at HN compared with LN. In contrast, heritability for yield traits (yield m-2, yield plant-1 HI and protein %) were higher at LN relative to HN but still record low values except for 100-grain weight. Nitrogen traits were sensitive to low N supplies where recorded 43.41-72.10% (Table 1) reduction than those under HN fertilizer compared with yield traits. Laperche et al. (2006) found a higher sensitivity of nitrogen traits (22-30%) under N¯ compared with yield traits (10%). Similarly, Makumbi (2005) in his study on evaluation of 25 inbred lines of maize and their F1 crosses at stress and optimal levels of N recorded low (0.11) to relatively high (0.56) heritability estimates for grain yield. They affirmed that, the low repeatability for grain yield and other tested traits suggests that actual heritability estimates for these traits might be low and progress to be made might be slow.
Regarding the crosses, the data showed opposite direction where nitrogen traits, except for NUEg, showed higher heritabilities values at LN. Heritability estimates recorded 29.18-88.20% decrease at HN over the LN treatment. In contrast, yield traits demonstrated lower heritability estimates at LN and showed 2.09-61.88% decrease over HN supplies. In general, crosses showed higher genetic variance at high and low N fertilizer than inbred lines pool for almost all measured traits. Sinebo et al. (2004) obtained close heritability values estimated for barley genotypes when data was combined at two levels of nitrogen and phosphorus for NHI (0.74), lower values for NUEg (0.62) and higher values for N-uptake (0.70) than those reported in the present study. They attributed the large broad-sense heritability values to the large diversity of the studied genotypes and the larger role of genotypes in portraying the phenotype. Laperche et al. (2006) confirmed the present results in that h2 decrease with N stress intensity is related to both decreasing in σ2g and σ2e. But the authors recorded higher h2 values at both N+ and N¯ for grain yield (0.85 and 0.86%), HI (0.86 and 0.86%) and grain protein yield (0.65 and 0.80%) compared with our data. Also Presterl et al. (2003) obtained higher and similar grain yield h2 at high and low N supplies (65%) compared with the current findings and suggested that direct selection under low conditions is more efficient to improve N-use efficiency for grain yield than direct selection at HN. In their study on genetic variability in grain yield of 296 single cross of maize evaluated under 60 kg N f-1, Radwan et al. (2003) found significant genotypic variance among the tested hybrids for all measured traits, they recorded high heritability estimates ranges from 0.51-0.80% for all traits, except days to 50% silking and HI which recorded low values (0.48 and 0.27%, respectively). There was expected substantial genetic progress from selection for plant biomass, grain yield plant-1, stover yield (13.4, 13.7 and 13.8%, respectively), whereas for other traits the expected genetic gain was low (2.80-6.5%).
Genetic Advance (GA)
Although heritability estimates had been found to be helpful in conducting selection on the basis of phenotypic performance, heritability estimates along with genetic gain were usually more useful than heritability values alone in predicting the effect of selection. Consequently genetic gain was estimated as percent of the means. Table 2 shows the expected genetic advance for the measured traits if 5% of plants are selected at optimal and stress N supplies. The GA values were similar for high and low nitrogen when estimated for inbred lines, while, showed higher values at high N fertilizer compared with stress environments for crosses.
Regarding inbred lines at low nitrogen fertilizer, stay green and total dry matter production recorded the highest genetic improvement followed by a moderate improvement for yield traits (yield, yield plant-1, 100-grain weight, protein % and HI) relative to means. The nitrogen traits (N-uptake, NUEg and NHI) demonstrated low genetic advance of 16.55, 26.13 and 22.84% above the overall mean for those traits when selected at LN. In contrast, NUEb showed slow expected genetic improvement of 7.12 g dry weight g N-1 each cycle of selection at both levels of fertilization. At high N fertilizer, grain yield traits demonstrated low expected genetic improvement, whereas nitrogen traits showed high genetic improvement compared with optimal agriculture.
For crosses, total dry matter recorded the highest expected genetic advance (52.85 and 69.90 g dw m-2) if selection is carried out at both N supplies, while NUEg recorder high genetic improvement at HN (53.16 g dw g N-1) and moderate values (38.97 g dw g N-1) at LN. Tefera et al. (2003), confirmed that, high GA for grain yield as percent of the mean accompanied with high genetic variability and moderate heritability values indicate the amenability for genetic improvement. Regarding yield traits, the expected genetic improvement was close when selected practiced at both N levels except HI which showed higher improvement when selected at HN. Days to 50% tasselling and silking expressed low genetic improvement at both levels of fertilizer, however recorded high h2 at both levels. This finding suggests a possibility for genetic improvement in advanced generations. As high heritability estimate with low genetic advance indicate the influence of non-additive gene effects and more effective selection in late generations.
Grain yield, HI, NUEg and HI could be genetically improved when selected under normal fertilizer since they combine moderate to high h2 and GA over the grand means. Whereas, NHI is expected to give 19.03 and 19.15% increase over the grand mean at both N supplies in late generations. In general, it is expected to improve the nitrogen and yield-related traits if selection is based in normal agriculture within the crosses pool. The same results were confirmed by Laperche et al. (2006) based on their work on wheat. Where the authors established that, indirect selection under N+ to improve productivity under N¯ conditions would appear to be most suitable program because of higher heritabilities under N+. In contrast, Presterl et al. (2003) established that, direct selection under low N conditions is more efficient to improve N¯ use efficiency for grain yield in maize than direct selection at HN due to high h2 and genetic variance estimates.
Heterosis Estimates
Heterosis effects for flowering, stay green, yield and nitrogen traits were estimated relative to mid-parent values and the single cross Pioneer 3062 performance at each N level and shown in Table 3-6.
Heterosis Based on Mid-Parent
At low and optimal N fertilizer, significant negative mid-parent heterosis was observed for days to 50% tasselling and silking dates and N-uptake at high N fertilizer. On the other hand, other traits showed positive and non-significant mid-parent heterosis at low N level. Protein % and grain yield and N-uptake demonstrated positive and negative significant mid-parent heterosis at high N level, respectively.
Under stress and optimal N fertilizer, crosses recorded positive heterotic effects for stay green, grain and nitrogen traits except N-uptake. Whereas, for flowering traits, the majority of crosses were earlier than their mid-parent since, heterosis showed negative values.
| Table 3: | Heterosis effects relative to mid-parent and SC 3062 values for flowering and stay green traits estimated at two levels of N fertilizer applications |
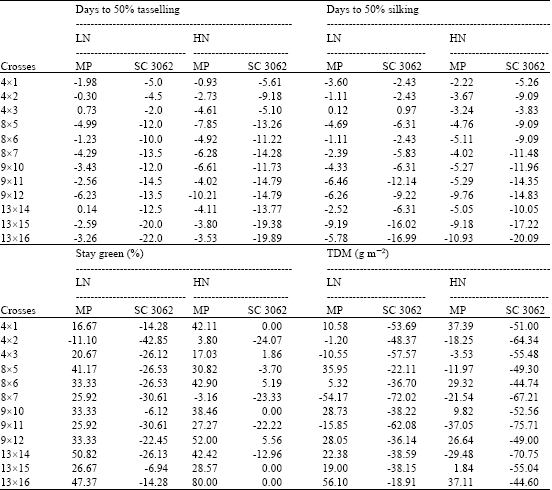 | |
The mean heterosis values were higher at stress compared with high N application (Table 3-6) for all measured traits except, NUEb and HI. Low heterosis values is an indicative for improving the performance of parent’s growth under increasing fertilizer consequently the mid-parents values increases, on the other hand the high mid-parent heterosis values at reduced level may attribute to the strong influence of non-additive gene effects which control the traits.
The presence of positive mid-parent heterosis values for the most traits is an indication for a non-additive gene effect with different degrees of dominance is portraying the inheritance of the concerned traits.
At N stress, there were obvious mid-parent heterotic effects and the best and highest heterosis values were demonstrated by the crosses, (4x1), (8x5), (8x6), (8x7), (9x11) and (9x12) for grain yield, yield plant-1, HI, NUEg and NHI. The crosses (8x5) and (9x12) recorded the highest mid-parent heterosis for protein% and NUEb. The cross (8x7) were the best one relative to its parental lines, where recorded-59. Thirty one percent reduction in N-uptake from the soil. For stay green trait, all crosses were superior to their mid-parents in leaf senescence delay, but three crossed {(13x14), (13x16) and (8x5)} showed the highest positive mid-parent heterosis. In contrast, days to 50% tasselling and silking showed negative desirable mid-parent heterosis, nearly all crosses recorded negative values, but the best were shown by the crosses, (13x15) and (9x12) (for days to 50% tasselling and silking, respectively).
| Table 4: | Heterosis effects relative to mid-present and SC3062 values for total dry matter, grain yield (m-2) and yield plant-1 estimated at two levels of N fertilizer applications |
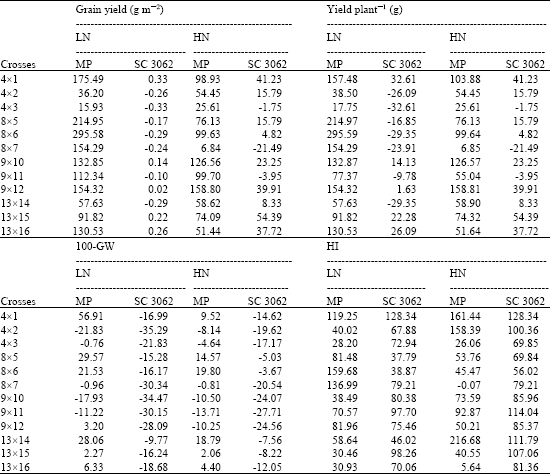 | |
Positive mid-parent heterosis for total dry matter at harvest was recorded for the crosses, (13x16) and (8x5), this result may be relate to the plant’s ability to capture more solar radiation, since, the same crosses showed mid-parent heterosis for stay green trait. The cross (13x16) combined the highest desirable heterotic effects for N-uptake and dry matter production.
At optimal N fertilizer, heterosis effects were not consistent for the different traits among different crosses. All F1 crosses were earlier in days to 50% tasselling and silking relative to their mean parent values (negative mid-parent heterosis). The best heterosis was recorded by the crosses, (9x12) for both traits. Regarding grain yield and the nitrogen traits (NUEg and NHI), crosses revealed desirable heterotic effects and recorded positive values. Specifically, the crosses, (9x12), (9x10), (9x11) and (4x1) showed the highest and positive heterosis for grain yield, NUEg, NUEb and NHI. Some crosses, showed positive mid-parent heterosis for total dry matter production, but the values were almost close. In contrast, the crosses, (9x11) were the best in N-uptake, since recorded the highest negative mid-parent value (-61.68%) or the lowest N-uptake.
Heterosis Against the Check SC 3062
Heterosis values relative to the check single cross for the measured traits are presented in Table (3-6). All F1 crosses surpassed the check variety for flowering traits, since recorded negative heterosis values (early flowering) at both N levels. In contrast, the Pioneer S.C. were superior to five crosses for stay green trait and less than three, while four crosses showed no heterosis relative to the check at high N application. Whereas, at low N, the single crosses were more senescence over all F1 crosses where recorded -42.85 to -6.12% decrease in greenness.
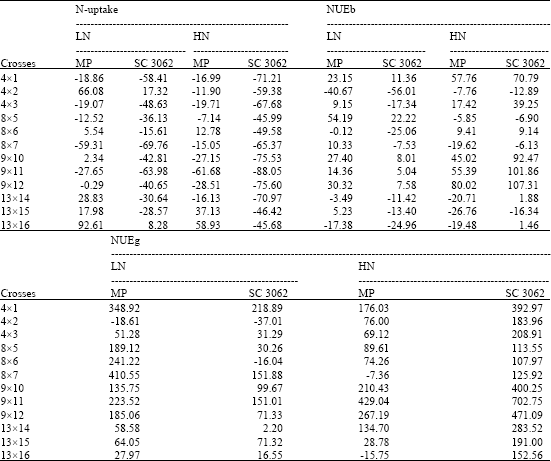
| Table 5: | Heterosis effects relative to mid-present and SC 3062 values for N-uptake and NUE traits estimated at two levles of N fertilizer applications |
 | |
| Table 6: | Heterosis effects relative to mid-present and SC 3062 values for NHI and protein % estimated at two levels of N fertilizer applications |
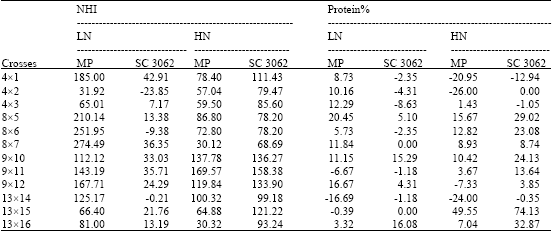 | |
The check variety showed inferior heterotic effects for total dry matter production at both N fertilizers, the reduction in means over the check ranged from -18.91 to -72.02% and from -44.60 to -67.21% at low and high N, respectively. On the contrary for N-uptake, all crosses showed best heterosis (negative values) over the check at high fertilizer, that is mean the check single cross is more exhausting to soil. Whereas under stress conditions only two crosses (4x2) and (13x16) showed higher N- uptake relative to the check.
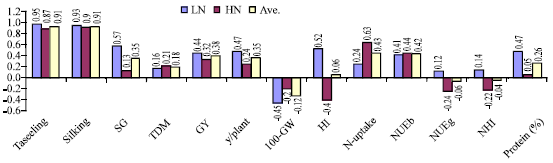 | |
| Fig. 1: | Midparent (inbreds)- offspring (crosses) correlation for flowering, yield and nitrogen traits at low, high and across fertilizer N rates, respectively in a nested mating design of 16 maize inbred lines |
HI, NUEg (except two crosses) and NHI (except three crosses) showed positive heterotic effects in the majority of crosses relative to the check at low N, except for grain yield, where crosses recorded negative and non-significant mean heterosis. The crosses, (9x11), (9x10), (9x12) and (4x1) recorded a positive and the highest heterosis values for HI, NUEg and NHI. For Protein%, only two crosses, (13x16) and (9x10) demonstrated an economic heterosis at low N fertilizer.
Correlation Between Inbreed Line per se and Cross Performance
The possibility of using inbred line information to indicate hybrid performance could reduce the need for hybrid evaluation was emphasized by Betrán et al. (2003). Therefore, correlation between mid-parent and cross performance was calculated at each N level and across levels and presented in (Fig. 1). Correlation between mid-parent and hybrid performance showed positive and greater values for low N fertilizer than under high N for all traits except, dry matter production, 100-grain weight, N-uptake and NUEb. The correlation showed significant and positive values for anthesis and silking dates and stay green traits, whereas for other traits the values were weak. The same results were established by many researchers (Lafitte and Edmeades, 1995; Gama and Hallauer, 1977), where they recorded positive and significant inbred-hybrid correlation at low and high N fertilizers. The present results were confirmed by Betrán et al., (2003), as they recorded a high (0.33*) and low (0.08) correlation values for grain yield between 17 maize inbred lines and their crosses in a diallel mating design evaluated at low and high N levels. Whereas under severe drought stress, the correlation was lower than under optimal stress. In addition, they obtained a low correlation (0.34) across environments. However in the present study the values were higher but non-significant than those obtained by the previous study (0.44 vs 0.32). Also correlation was not important under low N conditions as established by Lafitte and Edmeades (1995). The low correlation value is attributed to, as suggested by (Betrán et al., 2003), the inbreeding effect of the parents, since high stress intensity can be tolerated by early generation inbred lines. (Betrán et al., 2003) confirmed that, the weak correlation reported for grain yield might suggest that mid-parent heterosis would not be a good predictor of F1 hybrid performance under stress conditions.
CONCLUSIONS
The quantitative parameters estimated in the present study varied with the degree of applied stress. At both N rates there is a high opportunity for genetic improvement in inbred lines for the traits; stay green and total dry matter production because high h2 is accompanied with high genetic improvement. Whereas, for N-uptake, low heritability is accompanied with low GA indicating that selection is not effective under N stress cultivation and the non gene action in influencing those traits. In contrast, genetic improvement of 75.84 g N m-2 would achieve from one cycle of selection at high N fertilizer. NUEg and NHI would be more effective at high than at low N application (high h2 and GA). In contrast, grain yield and HI which combine low h2 and moderate GA give a predicted improvement of 50 and 30.5% over the grand mean at low N. Grain yield, NUEg, NUEb and N-uptake are expected to be genetically improved simultaneously under low N fertilizer cultivation within the hybrid pool.
The mid-parent heterosis responded differently according to the applied fertilizer rate. Crosses demonstrated desirable negative heterosis for days to 50% tasselling and silking and positive trend towards greenness at both N rates. The cross (8x7) is preferred under low agriculture because it reveal low N-uptake, but on the other hand recorded the lowest dry matter accumulation relative to its parents.. The crosses, (9x11) and (9x12) could be utilized for cultivation under a range of N fertilization, since they showed positive desirable heterosis for grain yield, yield plant-1, HI, NUEg and NHI.
The correlation between midparent and hybrid performance give interpretation for the hybrid performance under N stress conditions based on mid-parent performance for all measured traits, except total dry matter, 100-grain weight and N-uptake.
REFERENCES
- Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Antolin, M.C., J. Yoller and M. Sachez-Diaz, 1995. Effects of temporary drought on nitrate-fed and nitrogen-fixation alfalfa plants. Plants Sci., 107: 159-165.
CrossRef - Banziger, M. and H.R. Lafitte, 1997. Efficiency of secondary traits for improving maize for low-nitrogen target environments. Crop Sci., 37: 1110-1117.
CrossRefDirect Link - Banziger, M., G.O. Edmeades, D.L. Beck and M.R. Bellon, 2000. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice. CIMMYT, Mexico, ISBN: 970-648-46-3, Pages: 68.
Direct Link - Banziger, M. and A.O. Diallo, 2001. Progress in developing drought and N stress tolerant maize cultivars for Eastern and Southern Africa. Proceedings of the 7th Eastern and Southern Africa Regional Maize Conference, February 11-15, 2001, Nairobi, Kenya, pp: 189-194.
Direct Link - Betran, F.J., D. Beck, M. Banziger and G.O. Edmeades, 2003. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Sci., 43: 807-817.
CrossRefDirect Link - Blum, A., 1988. Plant Breeding for Stress Environments. CRC Press, Boca Raton, USA., ISBN-10: 0849363888, Pages: 223.
Direct Link - Hallauer, A.R. and J.B. Miranda, 1988. Quantitative Genetics in Maize Breeding. 2nd Edn., Iowa State University Press, Ames, Iowa, ISBN: 9780813815220, Pages: 468.
Direct Link - Hefny, M.M. and A.A. Aly, 2008. Yielding ability and nitrogen use efficiency in maize inbred lines and their crosses. Int. J. Agric. Res., 3: 27-39.
CrossRefDirect Link - Koutroubas, S.D. and D.A. Ntanos, 2003. Genotypic differences for grain yield and nitrogen utilization in Indica and Japonica rice under Mediterranean conditions. Field Crops Res., 83: 251-260.
Direct Link - Lafitte, H.R. and G.O. Edmeades, 1995. Association between traits in tropical maize inbred lines and their hybrids under high and low nitrogen. Maydica, 40: 259-267.
Direct Link - Laperche, A., M. Brancourt-Hulmel, E. Heumez, O. Gardet and J. Le Gouis, 2006. Estimation of genetic parameters of a DH wheat population grown at different N stress levels characterized by probe genotypes. Theor. Applied Genet., 112: 797-807.
Direct Link - Le Gouis, J. and P. Pluchard, 1996. Genetic variation for nitrogen use efficiency in winter wheat (Triticum aestivum L.). Euphytica, 92: 221-224.
Direct Link - Le Gouis, J., D. Beghin, E. Heumez and P. Pluchard, 2002. Diallel analysis of winter wheat at two nitrogen levels. Crop Sci., 42: 1129-1134.
Direct Link - Moll, R.H., E.J. Kamprath and W.A. Jackson, 1982. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agron J., 74: 562-564.
CrossRefDirect Link - Presterl, T., G. Seitz, M. Landbeck, E.M. Thiemt, W. Schmidt and H.H. Geiger, 2003. Improving nitrogen-use efficiency in European maize: Estimation of quantitative genetics parameters. Crop Sci., 43: 1259-1265.
Direct Link - Rosielle, A.A. and J. Hamblin, 1981. Theoretical aspects of selection for yield in stress and non-stress environment. Crop Sci., 21: 943-946.
CrossRefDirect Link - Sinebo, W., R. Gretzmacher and A. Edelbauer, 2004. Genotypic variation for nitrogen use efficiency in Ethiopian barley. Field Crops Res., 85: 43-60.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Tefera, H., K. Assefa, F. Hundera, T. Kefyalew and T. Teferra, 2003. Heritability and genetic advance in recombinant inbred lines of tef (Eragrostis tef). Euphytica, 131: 91-96.
CrossRef - Ud-Din, N., B.F. Carver and A.C. Clutter, 1992. Genetic analysis and selection for wheat yield in drought-stressed and irrigated environments. Euphytica, 62: 89-96.
CrossRefDirect Link