
P. Samuel
Department of Biotechnology, Bharath College of Science and Management, Thanjavur, Tamilnadu, India
A. Raja
Department of Microbiology, Jamal Mohamed college, Tiiruchirapalli Tamilnadu, India
P. Prabakaran
Department of Microbiology, PRIST University, Thanjavur, Tamilnadu, India
International Journal of Oceanography and Marine Ecological System
Year: 2012 | Volume: 1 | Issue: 1 | Page No.: 1-10







ABSTRACT
Over billions of years, the ocean has been regarded as the origin of life on Earth. The ocean includes the largest range of habitats, hosting the most life-forms. This review deals with the research and development work investigating the occurrence and bioprocessing of marine microbial enzymes. Competition amongst microorganisms for space and nutrients in the marine environment is a powerful selective force which has led to evolution. The evolution prompted the marine microorganisms to generate multifarious enzyme systems to adapt to the complicated marine environments. In this review, starting from the analysis of these featuring habitat-related properties, important examples of marine enzymes in biocatalysis will be reported. Therefore, marine microbial enzymes can offer novel biocatalyst with extraordinary properties.
PDF Abstract XML References Citation
Received: May 25, 2011;
Accepted: June 28, 2011;
Published: November 14, 2011
How to cite this article
P. Samuel, A. Raja and P. Prabakaran, 2012. Investigation and Application of Marine Derived Microbial Enzymes: Status and Prospects. International Journal of Oceanography and Marine Ecological System, 1: 1-10.
DOI: 10.3923/ijomes.2012.1.10
URL: https://scialert.net/abstract/?doi=ijomes.2012.1.10
DOI: 10.3923/ijomes.2012.1.10
URL: https://scialert.net/abstract/?doi=ijomes.2012.1.10
INTRODUCTION
Enzymes have been isolated and purified from microorganisms, animals and plants; among them microorganisms represent the most common source of enzymes because of their broad biochemical diversity, feasibility of mass culture and ease of genetic manipulation (Lam, 2006). The world’s oceans cover more than 70% of our planet’s surface; countless marine microorganisms contain biochemical secrets which can provide new insights and understanding of enzymes. Marine microorganisms have been attracting more and more attention as a resource for new enzymes because the microbial enzymes are relatively more stable and active than the corresponding enzymes derived from plants or animals (Bull et al., 2000). A growing number of marine microorganisms are the sources of novel and potentially life-saving bioactive secondary metabolites. Here, we have discussed some of these novel antibacterial, antiviral, anticancer compounds isolated from marine-derived microbes and their possible roles in disease eradication and commercial exploitation of these compounds for possible drug development using many approaches (Raja et al., 2010).
With the recent advent of biotechnology, there has been a growing interest and demand for enzymes with novel properties. When compared with the terrestrial environment, the marine environment gives marine microorganisms, unique genetic structures and life habitats (Stach et al., 2003). The marine environment ranges from nutrient-rich regions to nutritionally sparse locations where only a few organisms can survive. The complexity of the marine environment involving high salinity, high pressure, low temperature and special lighting conditions, may contribute to the significant differences between the enzymes generated by marine microorganisms and homologous enzymes from terrestrial microorganisms. Hence, marine microbial enzyme technology boosted in recent years and resulting in the exploration of valuable products. These enzymes are used as pharmaceuticals, food additives and fine chemicals (De Miguel Bouzas et al., 2006). In recent years, researchers have isolated a variety of enzymes with special activities from marine bacteria, actinomycetes, fungi and other marine microorganisms and some products have already been used in industrial and medical applications. Mycothiol (MSH or AcCys-GlcN-Ins) is an unusual thiol compound found in the Actinobacteria including Actinomycetes and Mycobacterium. The enzymes involved in mycothiol biosynthesis are of interest as potential targets for new drugs directed against Mycobacterium tuberculosis. MSH-dependent enzymes that utilize MSH for their activity either as a cofactor or as a substrate indicate that MSH plays a key role in protecting cells against a variety of challenges. The enzymes of MSH metabolism could be possible targets for development of new drugs and vaccines directed against Mycobacterium tuberculosis and other infectious Actinomycetes (Vignesh et al., 2011). In particular, some marine microbial enzymes have yielded a considerable number of drug candidates. Marine microorganisms whose immense genetic and biochemical diversity is still in its infant stage, are of considerable current interest as a new promising source of enzymes with unsuspected application potentials (Debashish et al., 2005). This study is devoted to the analysis of novel chemical and stereochemical biodiversity offered by marine biocatalyst with particular emphasis on current or potential applications of these enzymes in chemical and pharmaceutical fields.
Protease: Total protease sales represent more than 60% of all industrial enzyme sales in the world. In modern society, the proteases are widely used. Proteases are used in the detergent industry, leather industry and also for pharmaceutical applications, such as digestive drugs and anti-inflammatory drugs (Kolodziejska and Sikorski, 1996). An alkaline protease, previously isolated from a symbiotic bacterium found in the Gland of Deshayes of a marine shipworm, was evaluated as a cleansing additive (Greene et al., 1996). A yeast strain (Aureobasidium pullulans) with a high yield of alkaline protease was isolated from sea saltern of the China Yellow Sea and the maximum production of enzyme was 623.1 U mg-1 protein (7.2 U mg-1) (Chi et al., 2007). In 2009, Bacillus mojavensis A21 producing alkaline proteases was isolated from seawater by Haddar et al. (2009) and they purified two detergent-stable alkaline serine-proteases (BM1 and BM2) from this strain. Both proteases showed high stability towards non-ionic surfactants. In addition, both of them showed excellent stability and compatibility with a wide range of commercial liquid and solid detergents (Haddar et al., 2009).
Lipase: Lipases are ubiquitous enzymes that catalyze the breakdown of fats and oils with subsequent release of free fatty acids, diacylglycerols, monoglycerols and glycerol. Besides this, lipases are also efficient in various reactions such as esterification, transesterification and aminolysis. Lipases have received increased attention recently, as evidenced by the increasing amount of information about lipases in the current literature. Also, many microbial lipases are available as commercial products, the majority of which are used in detergents, paper production, cosmetic production, food flavoring, organic synthesis and some other industrial applications (Chi et al., 2009). The enzyme detergent market share has currently reached 90% in Europe and in Japan around 80%. Lipases are valuable biocatalyst because they act under mild conditions and are highly stable in organic solvents, show broad substrate specificity (Davidson, 2006).
Microbial lipase was first found from Penicillium oxalicum and Aspergillus flavus in 1935 by Kirsh (David, 1935). Feller et al. (1990) screened four cold-adapted lipases secreted by Moraxella. These Moraxella were obtained from the Antarctic seawater with the optimum growth temperature of 25°C and the maximum secretion of lipases was supposed to occur at lower temperature conditions, the lowest secretion temperature can reach 3°C. Wang et al. (2007) screened out nine lipase producing strains from a total of 427 yeast strains. They belonged to Candida intermedia YA01a, Pichia guilliermondii N12c, Candida parapsilosis 3eA2, Lodderomyces elongisporus YF12c, Candida quercitrusa JHSb, Candia rugosa wl8, Yarrowia lipolytica N9a, Rhodotorula mucilaginosa L10-2 and Aureobasidium pullulans HN2.3. Some lipases could actively hydrolyze different oils, indicating that they may have potential applications in industry (Wang et al., 2007). In 2009, a novel extracellular phospholipase C was purified from a marine streptomycete which was selected from approximately 400 marine bacteria by Mo et al. (2009). Its enzyme activity was optimal at pH 8.0 at 45°C and it hydrolyzed only phosphatidylcholine (Mo et al., 2009).
POLYSACCHARIDE-DEGRADING ENZYMES
Chitinase and chitosanase: Chitin is widely distributed in nature as a biopolymer with non-toxic properties. After cellulose, it is the most common polysaccharide found in nature and is the major structural component of most fungi cell walls and also quite abundant in the crust of insects and crustaceans. In nature, annual generation of chitin is about 1.0x1010 t (Rinaudo, 2006). Chitin and chitosan have a similar chemical structure. Chitin is made up of a linear chain of acetylglucosamine groups and chitosan is obtained by removing enough acetylglucosamine residues. After hydrolysis, chitin and chitosan could enhance immune function, promote digestive function and eliminate toxins from the body, even inhibit tumor cell growth as well as being involved in other important physiological functions (Ngo et al., 2008). Therefore, hydrolysis of chitin and chitosan recently became a hot topic.
As marine zooplankton are regularly supposed to shed, there is a large amount of abandoned chitin which could be a rich source of carbon and energy for growth and reproduction of chitin-degrading microorganisms. The total production of chitin in the whole marine biocycle is at least 2.3 million metric tons per year. Until now, researchers have found a wide range of microorganisms that can produce chitinase or chitosanase, including Aspergillus, Penicillium, Rhizopus, Myxobacter, Sporocytophaga, Bacillus, Enterobacter, Klebsiella, Pseudomonas, Serratia, Chromobacterium, Clostridium, Flavobacterium, Arthrobacter and Streptomyces (Xia et al., 2008).
Alginate lyases: The brown alga is one of the largest marine biomass resources. Alginate has a wide range of applications; further, the degraded low-molecular fragment shows more potential. Alginate lyases, characterized as either mannuronate or guluronate lyases, are a complex copolymer of α-L-guluronate and its C5 epimer β-D-mannuronate. They have been isolated from a wide range of organisms, including algae, marine invertebrates and marine and terrestrial microorganisms. In recent years, the marine microbial alginate lyases have been greatly developed. Discovering and characterization of alginate lyases will enhance and expand the use of these enzymes to engineer novel alginate polymers for applications in various industrial, agricultural and medical fields (Alkawash et al., 2006).
Agarases: Agar is a highly heterogeneous polysaccharide. Neutral agarose is an alternating polymer of D-galactose and 3,6-anhydro-L-galactose linked by alternating β1→4 and α1→3 bonds. Agar oligosaccharides have a wide range of applications in the food industry; it can be used for beverages, bread and some low-calorie food production. Japanese use agar-oligosaccharide as a moisturizing cosmetic additive and it also has good hair conditioning effects (Rasmussen and Morrissey, 2007). Now-a-days, the acid degradation of agar is replacing enzymatic degradation with the advantages of easy control and mild reaction. Agarase is an enzyme found in agarolytic microorganisms. Agar-degrading microorganisms can be divided into two groups: Bacteria soften the agar; the other violently liquefies the agar. Until now, researchers have found the presence of agarase from species within the genus Cytophaga, Bacillus, Vibrio, Alteromonas, Pseudoalteromonas, Streptomyces (Hosoda et al., 2003). A marine bacterium Vibrio sp. (JT0107) which can hydrolyze the α-l,3 glycosidic bond of agar by α-Neoagaro-oligosaccharide residues. Several agarase genes have been cloned and sequenced. This agarase hydrolyzes not only agarose but also neoagarotetraose to yield neoagarobiose. This is a unique characteristic for a β-agarase (Sugano et al., 1993). In 1994, from the same species of bacteria, a new β-agarase gene (aga B) was sequenced by the same group. A new β-agarase was purified from an agarolytic bacterium, Bacillus sp. MK03 in 2003; this enzyme could hydrolyze neoagarohexaose to produce neoagarotetraose and neoagarobiose. Researches also demonstrated that glucose can inhibit extracellular agarase secretion without transcription inhibition (Suzuki et al., 2003).
Carrageenases: Carrageenan and carrageenin are a family of linear sulfated polysaccharides which are extracted from red seaweeds. Eighty percent of the carrageenan is used in food and food-related industries and it can be used as a coagulant, adhesive, stabilizer and emulsifier. In addition, it has also been widely applied in the pharmaceutical and cosmetics industries. The oligosaccharides obtained from carrageenan degradation show a variety of specific physiological activities, such as anti-viral, anti-tumor, anti-coagulation, etc. (Roberts et al., 2007). As early as 1943, Mori extracted carrageenase from marine mollusc. Right now, Pseudomonas, Cytophaga, Alteromonas atlantica, Alteromonas carrageenovora and some unidentified strains have been found to possess the carrageenan-degrading enzymes (Mori, 1943). Sarwar et al. (1987) using carrageenan containing medium, cultured Cytophaga lk-C783 and obtained extracellular κ-carrageenase with a molecular weight of 10 kD. Mo et al. (2009) isolated an extracellular κ-carrageenase with a molecular weight of 30 kD from marine Cytophaga MCA-2. A distinct λ-Carrageenan-degrading Pseudoalteromonas bacterium (CL19) was isolated from a deep-sea sediment sample the molecular mass of this purified enzyme was approximately 100 kD (Ohta and Hatada, 2006).
Cellulose and hemicellulose hydrolase: Cellulose is an organic compound with the formula (C6H10O5) n; a polysaccharide consisting of a linear chain of several hundred to over ten thousand β (1→4)-linked D-glucose units. Hemicellulose is a polysaccharide related to cellulose and in contrast to cellulose, it can be derived from several sugars including glucose, xylose, mannose, galactose, rhamnose and arabinose. Hemicellulose consists of shorter chains of around 200 sugar units. Cellulose is the most available saccharide in nature and is about 50% of all plant matter and hemicellulose is ~20-30% while the remainder is mainly lignin (Klemm et al., 2005). Until now, it was found that bacteria can produce cellulase, including: Cytophaga, Cellulomonas, Vibrio and Clostridium, Nocardia, Streptomyces and for certain fungi it was found that Trichoderma, Aspergillus, Fusarium, Chaetomium, Phoma, Sporotrichum, Penicillium, etc. are also able to produce cellulase. Hemicellulase, generally refers to the hydrolase which can hydrolyze polysaccharides, for example, xylanase, galactanase, arabanase, among which xylanase has particular economic value.
Cellulase can be used for bio-textile auxiliaries, cotton and linen products processing and bio-fertilizer processing. With the rapid development of the seaweed industry, a mass of waste released into the environment led to very serious pollution problems. Cellulases degrade seaweed processing waste to low molecular fragments which can be easily absorbed by plants as bio-fertilizer. Xylanases are hydrolases depolymerize the plant cell component xylan, the second most abundant polysaccharide. Xylanases could be produced by fungi, bacteria, yeast, marine algae etc. but the principal commercial source is filamentous fungi. Xylanase could be used on semi-cellulose to produce products with high economic value, such as xylitol. In the paper and pulp industry, using xylanase improves the lignin dissolution rate and reduces the usage of Cl2 and ClO2, thereby reducing the pollution. Xylanase can also degrade some polysaccharides in juice or beer, thus it could contribute to beverage clarification (Maki et al., 2009). Indian researchers obtained several fungal isolates from marine habitat showed alkaline xylanase activity. The crude enzyme from NIOCC isolate No. 3 (Aspergillus niger) with high xylanase activity, cellulase-free and unique properties containing 580 UL-1 of xylanase. Yin et al. (2010) purified xylanase which had an optimal pH and temperature at 5.0 and 50°C from bacterium Bacillus sp. YJ6 (Yin et al., 2010). A novel cold-adaptive xylanolytic Penicillium strain FS010 was isolated from China Yellow sea sediments this fungus grew well from 4 to 20°C but a lower (0°C) or higher (37°C) temperature limited its growth. Compared with the mesophilic Penicillium chrysogenum, the cold-active xylanase showed high hydrolytic activities at low temperature (2-15°C) and high sensitivity to high temperature (>50°C) (Hou et al., 2006).
Other polysaccharide hydrolases: With the development of marine science and technology, researchers reported more and more microorganisms from marine habitats capable of producing amylase. The marine yeast strain Aureobasidium pullulans N13d, producing an extracellular amylase, was isolated from the deep sea sediments of Pacific Ocean (Li et al., 2007). Chakraborty et al. (2009) reported a novel α-amylase from marine Streptomyces sp. D1 by using media containing 2% sucrose, 0.35% peptone and 0.15% of malt extract (Chakraborty et al., 2009). Mohapatra et al. (1998) isolated a novel amylase from the Mucor sp. associated with the marine sponge Spirastrella sp., this enzyme has an optimum pH of 5.0 and an optimum temperature of 60°C.
EXTREMOZYMES
The marine environment is extremely complex, including low-temperature, high-temperature (-2 to ~113), high hydrostatic pressure (9.53 MPa), strong acid, strong alkali and very poor nutritional conditions. An extremophilic microorganism is a microorganism that thrives in physically or geochemically extreme conditions that are detrimental to the majority of life on Earth. Extremophilic microorganisms are adapted to survive in ecological niches, they must adaptively change their physiological structure and metabolism, in order to adapt to the extreme environmental conditions (Table 1).
Raja et al. (2011a, b) reported new antibiotics that are active against resistant bacteria. Bacteria have lived on Earth for several billion years.
| Table 1: | Extremophiles and living environments |
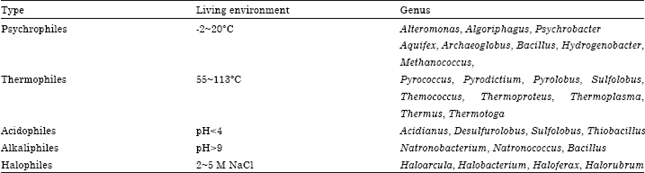 | |
During this time, they encountered in nature a wide range of naturally occurring antibiotics, in order to survive, bacteria developed antibiotic resistance mechanism (Hoskeri et al., 2010)
Therefore, extremophilic microorganisms screened from these environments, may have some specific physiological principles which can produce unique biocatalysts that function under extreme conditions comparable to those prevailing in various industrial enzymes. Because of this reason, in recent years, research programs investigating extremophilic microorganisms became a new area of interest in microbial research and are very popular (Niehaus et al., 1999).
Algoriphagus, Psychrotrophs and other low-temperature microorganisms have obvious advantages in the ecology. The low-temperature microorganisms cannot readily be involved in contamination. Their culture condition is simple and the enzymes from these microorganisms have advantages of high activity and high catalytic efficiency. Hence, with the assistance of low-temperature microbial enzymes, it can greatly shorten the processing time and save expensive heating/cooling systems, thus save considerable energy. Cold-adapted enzymes from marine microorganisms, especially, the lipases and proteases, have considerable potential, particularly in the cleaning industry. Many studies have shown that about 77% of Antarctic marine bacteria are resistant to cold environments and 23% are addicted to a cold environment. The unique geography and climate characteristics in Antarctica forms a dry, bitterly cold, strong radiation environment, in which microorganisms have to survive with corresponding unique molecular mechanism, physiological and biochemical characteristics. For these reasons, the Antarctic marine bacteria are thought to produce new bioactive substances with significant potential. Kolenc transferred TOLUENE plasmid pWWO of mesophilie Psychrotrophs putida PaW1 to the psychrotroph Psychrotrophs putida Q5. From expression of the genes, it was shown that the transconjugant had the capacity to degrade and utilize toluate (1,000 mg L-1) as a sole source of carbon at temperatures as low as 0°C (Kano et al., 1997). Transferring the useful gene from mesophilic microorganisms to psychrophilic ones was established to promote low-temperature microbial biological features which may have enormous potential in removing pollution in cold environments.
Near the deep sea volcanoes, some microorganisms can survive in extreme conditions, even over temperatures of 100°C. Therefore, these microorganisms are supposed to have unique enzyme systems which can work in these high temperature conditions. For example, the nucleic acid enzymes, such as DNA polymerase, ligase and restriction endonuclease, have a significant applicational value in molecular biology research. Iundberg’s group purified a thermostable DNA polymerase from thermophilic archaea (Pyrococcus furiosus) which has polymerizing and proofreading double functions and has high activity even at 100°C. Hence, this polymerase can be applied in high fidelity PCR experiments. In 2008, a novel thermostable non-specific nuclease from thermophilic bacteriophage GBSV1 was isolated by Song’s group, this non-specific nuclease can degrade various nucleic acids, including RNA, single-stranded DNA and double-stranded DNA (Song and Zhang, 2008). The deep-sea is regarded as an extreme environment with conditions of high hydrostatic pressure (9.53 MPa). Enzymes from deep-sea microorganisms are thought to have characteristic pressure-adaptation mechanisms in structure and function and they can be utilized in high hydrostatic pressure environments without disintegration. Microorganisms obtained from deep-sea environments appear to be an important source of modern enzyme industries. In 1979, the first barophilic bacteria was isolated from a deep-sea sample and has been found to grow optimally at about 500 bars and 2-4°C. Japanese scientists isolated multiple strains of bacteria addicted to pressure from the marine environment and found that the in vivo genes, proteins and enzymes in the deep-sea still have a high ability. The discovery and research of the marine barophilic microorganisms provide a good foundation for further development on extreme enzymes. There are some regions in oceans where microorganisms from these locales are commonly highly acidophilic or alkalophilic: They can live in conditions of pH 5, even below pH 1 or alternatively over pH 9 conditions. Extracellular enzymes secreted by these microorganisms are commonly acidophilic enzymes (optimum pH<3.0) or alkalophilic enzymes (optimum pH>9.0). Compared with the neutral enzymes, the extreme pH enzymes show good stability in the environment, due to the particular enzyme molecule containing high proportion of acidic or basic amino acids. The enzymes, produced by acidophilic or alkalophilic microorganisms, could have wide applications for compound synthesis in extreme pH conditions (Adams et al., 1995).
DISCUSSION
The 21st century is the century of the ocean and the ocean is a vast treasure of human life. Recently, most countries face similar problems such as high population, resource consumption and pollution. Meanwhile, the marine biological progress and development gives a new source and options to humans. Marine microbial enzymes, especially marine extreme microbial enzymes, have become more and more important in applications.
Enzyme metabolism is a fundamental biological process that is vital for the survival of all species. Their specific function is to catalyze chemical reactions. Enzymes have found wide and diverse applications at which enzymes increase the rate of reactions which approach to equilibrium. Enzymes play critical role in the metabolic activities of all living organisms whether humans, animals, plants or microorganisms and are widely applied in microbial technology and their diagnosis processes (Raja et al., 2010).
Because enzymes have unequalled advantages, many industries are keenly interested in adapting enzymatic methods to the requirements of their processes. Clinical application of enzymes has been developing, For example, surgeons used proteolytic enzymes for debridement of wounds. Promising clinical results have been reported by injection of certain enzymes such as streptokinase, crystalline trypsin and chymotrypsin. Since the increased therapeutic use of enzymes, presently unpredictable, rapid advances in this field may be expected (Underkofler et al., 1958).
The complexity of the marine environment involving high salinity, high pressure, low temperature and special lighting conditions, may contribute to the significant differences between the enzymes generated by marine microorganisms and homologous enzymes from terrestrial microorganisms, leading to the boosted marine microbial enzyme technology in recent years and the resulting valuable products. These enzymes are used as pharmaceuticals, food additives and fine chemicals (Bernan et al., 1997).
Japan constantly increases its support to marine microbial enzymes research and from 1992, the Japanese government made a series of marine microorganisms, planned to discover and clone proteins or enzymes with some special activity. In addition, Canada, Spain, Finland and Russia and other countries have also stepped up on marine bio-enzyme research.
Collectively, due to marine biological diversity and the specificity of biological metabolism, the study on a global scale is still just beginning but it has huge potential for development and applications with industrial benefits.
REFERENCES
- Adams, M.W.W., F.B. Perler and R.M. Kelly, 1995. Extremozymes: Expanding the limits of biocatalysis. Nat. Biotechnol., 13: 662-668.
CrossRefDirect Link - Alkawash, M.A., J.S. Soothill and N.L. Schiller, 2006. Alginate lyase enhances antibiotic killing of mucoid Pseudomonas aeruginosa in biofilms. APMIS, 114: 131-138.
CrossRefPubMedDirect Link - Bernan, V.S., M. Greenstein and W.M. Maiese, 1997. Marine microorganisms as a source of new natural products. Adv. Applied Microbiol., 43: 57-90.
PubMedDirect Link - Bull, A.T., A.C. Ward and M. Goodfellow, 2000. Search and discovery strategies for biotechnology: The paradigm shift. Microbiol. Mol. Biol. Rev., 64: 573-606.
PubMedDirect Link - Chakraborty, S., A. Khopade, C. Kokare, K. Mahadik and B. Chopade, 2009. Isolation and characterization of novel α-amylase from marine Streptomyces sp. D1. J. Mol. Cataly. B: Enzym., 58: 17-23.
CrossRef - Chi, Z., Z. Chi, T. Zhang, G. Liu, J. Li and X. Wang, 2009. Production, characterization and gene cloning of the extracellular enzymes from the marine-derived yeasts and their potential applications. Biotechnol. Adv., 27: 236-255.
PubMedDirect Link - Chi, Z., C. Ma, P. Wang and H.F. Li, 2007. Optimization of medium and cultivation conditions for alkaline protease production by the marine yeast Aureobasidium pullulans. Bioresour. Technol., 98: 534-538.
CrossRefDirect Link - David, K., 1935. Lipase production by Penicillium Oxalicum and Aspergillus flavus. Bot. Gazzette, 97: 321-333.
Direct Link - Davidson, M.H., 2006. Mechanisms for the hypotriglyceridemic effect of marine omega-3 fatty acids. Am. J. Cardio., 98: 27-33.
PubMedDirect Link - Debashish, G., S. Malay, S. Barindra and M. Joydeep, 2005. Marine enzymes. Adv. Biochem. Eng. Biotechnol., 96: 189-218.
CrossRefPubMedDirect Link - Feller, G., M. Thiry, J.L. Arpigny, M. Mergeay and C. Gerday, 1990. Lipases from psychrotrophic antarctic bacteria. FEMS Microbiol. Lett., 66: 239-244.
CrossRef - Greene, R.V., H.L. Griffin and M.A. Cotta, 1996. Utility of alkaline protease from marine shipworm bacterium in industrial cleansing applicat. Biotechnol. Lett., 18: 759-764.
CrossRef - Haddar, A., R. Agrebi, A. Bougatef, N. Hmidet, A. Sellami-Kamoun and M. Nasri, 2009. Two detergent stable alkaline serine-proteases from Bacillus mojavensis A21: Purification, characterization and potential application as a laundry detergent additive. Bioresour. Technol., 100: 3366-3373.
CrossRefDirect Link - Hoskeri, H.J., V. Krishna and C. Amruthavalli, 2010. Effects of extracts from lichen Ramalina pacifica against clinically infectious bacteria. Researcher, 2: 81-85.
Direct Link - Hosoda, A., M. Sakai and S. Kanazawa, 2003. Isolation and characterization of agar-degrading Paenibacillus spp. associated with the rhizosphere of Spinach. Biosci. Biotechnol. Biochem., 67: 1048-1055.
CrossRefPubMedDirect Link - Hou, Y.H., T.H. Wang, H. Long and H.Y. Zhu, 2006. Novel cold-adaptive Penicillium strain FS010 secreting thermo-labile xylanase isolated from Yellow Sea. Acta Biochim. Biophys. Sin, 38: 142-149.
CrossRefPubMedDirect Link - Kolodziejska, I. and Z.E. Sikorski, 1996. Neutral and alkaline muscle proteases of marine fish and invertebrates a review. J. Food Biochem., 20: 349-364.
CrossRefDirect Link - Kano, H., S. Taguchi and H. Momose, 1997. Cold adaptation of a mesophilic serine protease, subtilisin, by in vitro random mutagenesis. Applied Microbiol. Biotechnol., 47: 46-51.
PubMedDirect Link - Lam, K.S., 2006. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol., 9: 245-251.
CrossRefPubMedDirect Link - Klemm, D., B. Heublein, H.P. Fink and A. Bohn, 2005. Cellulose: Fascinating biopolymer and sustainable raw material. Angewandte Chemie Int. Edn., 44: 3358-3393.
CrossRefPubMedDirect Link - Li, H., Z. Chi, X. Wang and C. Ma, 2007. Amylase production by the marine yeast Aureobasidium pullulans N13d. J. Ocean Univ. Chin., 6: 60-66.
CrossRefDirect Link - Maki, M., K.T. Leung and W. Qin, 2009. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int. J. Biol. Sci., 5: 500-516.
CrossRefDirect Link - Rinaudo, M., 2006. Chitin and chitosan: Properties and applications. Prog. Polym. Sci., 31: 603-632.
CrossRefDirect Link - Mo, S.J., J.H. Kim and K.W. Cho, 2009. Enzymatic properties of an extracellular phospholipase C purified from a marine Streptomycete. Biosci. Biotechnol. Biochem., 73: 2136-2137.
CrossRefPubMedDirect Link - Mohapatra, M.A., U.C. Banerjee and M. Bapuji, 1998. Characterization of a fungal amylase from Mucor sp. associated with the marine sponge Spirastrella sp. J. Biotechnol., 60: 113-117.
CrossRefDirect Link - Raja, M.M.M., A. Raja, M.M. Imran, A.M.I. Santha and K. Devasena, 2011. Enzymes application in diagnostic prospects. Biotechnology, 10: 51-59.
CrossRefDirect Link - Ngo, D.N., M.M. Kim and S.K. Kim, 2008. Chitin oligosaccharides inhibit oxidative stress in live cells. Carbohydr. Polym., 74: 228-234.
CrossRef - Niehaus, F., C. Bertoldo, M. Kahler and G. Antranikiar, 1999. Extremophiles as a source of novel enzymes for industrial application. Applied Microbiol. Biotechnol., 5: 711-729.
PubMedDirect Link - Ohta, Y. and Y. Hatada, 2006. A novel enzyme, λ-carrageenase, isolated from a deep-sea bacterium. J. Biochem., 140: 475-481.
CrossRefPubMedDirect Link - Rasmussen, R.S. and M.T. Morrissey, 2007. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res., 52: 237-292.
CrossRefPubMedDirect Link - Raja, A., P. Prabakaran and P. Gajalakshmi, 2010. Isolation and screening of antibiotic producing psychrophilic actinomycetes and its nature from rothang hill soil against viridans Streptococcus sp. Res. J. Microbiol., 5: 44-49.
CrossRefDirect Link - Vignesh, S., A. Raja and R.A. James, 2011. Marine drugs: Implication and future studies. Int. J. Pharmacol., 7: 22-30.
CrossRefDirect Link - Raja, M.M.M., A. Raja, M.M. Imran and E.V. Ugin, 2011. Anti mycobacterial activity of actinomycetes producing mycothiol. Singapore J. Sci. Res., 1: 144-148.
Direct Link - Roberts, J.N., C.B. Buck, C.D. Thompson, R. Kines and M. Bernardo et al., 2007. Genital transmission of HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat. Med., 13: 857-861.
CrossRefPubMedDirect Link - Song, Q. and X. Zhang, 2008. Characterization of a novel non-specific nuclease from thermophilic bacteriophage GBSV1. BMC Biotechnol., 8: 43-43.
CrossRefPubMedDirect Link - Stach, J.E.M., L.A. Maldonado, A.C. Ward, M. Goodfellow and A.T. Bull, 2003. New primers for the class Actinobacteria: Application to marine and terrestrial environments. Environ. Microbiol., 5: 828-841.
CrossRefPubMedDirect Link - Sugano, Y., T. Matsumoto, H. Kodama and M. Noma, 1993. Cloning and sequencing of agaA, a unique agarase 0107 gene from a marine bacterium, Vibrio sp. strain JT0107. Applied Environ. Microbiol., 59: 3750-3756.
PubMedDirect Link - Suzuki, H., Y. Sawai, T. Suzuki and K. Kawai, 2003. Purification and characterization of an extracellular β-agarase from Bacillus sp. MK03. J. Biosci. Bioeng., 95: 328-334.
CrossRefPubMedDirect Link - Underkofler, L.A., R.R. Barton and S.S. Rennert, 1958. Production of microbial enzymes and their applications. Applied Microbiol., 6: 212-221.
PubMed - Wang, L., Z.M. Chi, X.H. Wang, Z.Q. Liu and J. Li, 2007. Diversity of lipase-producing yeasts from marine environments and oil hydrolysis by their crude enzymes. Ann. Microbiol., 57: 495-501.
CrossRefDirect Link - Xia, W., P. Liu and J. Liu, 2008. Advance in chitosan hydrolysis by non-specific cellulases. Bioresour. Technol., 15: 6751-6762.
ASCIDirect Link - Yin, L.J., H.H. Lin, Y.I. Chiang and S.T. Jiang, 2010. Bioproperties and purification of xylanase from Bacillus sp. J. Agric. Food Chem., 58: 557-562.
CrossRefPubMedDirect Link - De Miguel Bouzas, T., J. Barros-Velazquez and T.G. Villa, 2006. Industrial applications of hyperthermophilic enzymes: A review. Protein Pept. Lett., 13: 645-651.
PubMed