
Samuel E. Agarry
Biochemical Engineering Research Laboratory, Department of Chemical Engineering, Ladoke Akintola University of Technology, Ogbomoso, Nigeria
Bamidele O. Solomon
Biochemical Engineering Research Laboratory, Department of Chemical Engineering, Obafemi Awolowo University, Ile-Ife, Nigeria
Thomas O.K. Audu
Department of Chemical Engineering, University of Benin, Benin-City, Nigeria
International Journal of Chemical Technology
Year: 2011 | Volume: 3 | Issue: 1 | Page No.: 1-13







ABSTRACT
This study was carried out to estimate and evaluate the growth parameters of indigenous binary mixed culture of Pseudomonas aeruginosa and Pseudomonas fluorescence on phenol in an aerobic chemostat culture. The analysis was done by checking the obtained biodegradation data for consistency using carbon and available electron balances and estimating the growth parameters using two forms of growth models: reparameterized Pirt’s model based on Monod approach and a modified model based on substrate consumption rate being rate limiting. The results showed that the biodegradation data were consistent and the growth parameter estimates obtained using the two forms of models were similar. Using the reparameterized Pirt’s model coupled with covariate adjustment estimation technique (CAT), the best growth parameter estimates were the Maximum Likelihood Estimates (MLE) based on the complete data. The results also revealed that phenol could be oxidatively metabolized with a maximum of about 38-41% of the energy contained in it incorporated into binary mixed culture of P. aeruginosa and P. fluorescence biomass. The balance of 58-62% is mostly evolved as heat with little energy needed for maintenance of the cells. Thus, the study demonstrates that phenol supports the growth of Pseudomonas species.
PDF Abstract XML References Citation
Received: July 30, 2010;
Accepted: January 08, 2011;
Published: May 12, 2011
How to cite this article
Samuel E. Agarry, Bamidele O. Solomon and Thomas O.K. Audu, 2011. Bioenergetics of Binary Mixed Culture of Pseudomonas aeruginosa and Pseudomonas fluorescence Growth on Phenol in Aerobic Chemostat Culture. International Journal of Chemical Technology, 3: 1-13.
DOI: 10.3923/ijct.2011.1.13
URL: https://scialert.net/abstract/?doi=ijct.2011.1.13
DOI: 10.3923/ijct.2011.1.13
URL: https://scialert.net/abstract/?doi=ijct.2011.1.13
INTRODUCTION
Excellent design and operation of biological systems have the potential of being the most cost effective ways to treat toxic and hazardous chemicals because almost complete oxidation may be accomplished (Amer et al., 2008). The toxicity of phenol and the need to find ways of removing it from the environment have made the compound a prime candidate for study. Many microbes are capable of utilizing phenol as a source of carbon and energy provided it is not present in too high concentrations (Pawlowsky et al., 1973; Solomon et al., 1994; Ruiz-Ordaz et al., 2001; Paller et al., 1995; Hill and Robinson, 1975; Nikakhtari and Hill, 2006; Afzal et al., 2007; Udayasoorian et al., 2007; Agarry et al., 2008; Agarry et al., 2010). Several studies have been carried out on the kinetics of phenol degradation by various microorganisms and on its inhibitory effects (Hill and Robinson, 1975; Yang and Humphrey, 1975; Schroeder et al., 1997; Folsom et al., 1990; Oboirien et al., 2005; Agarry and Solomon, 2008; Li et al., 2010). The concept of material and energy balance in biotechnology has been identified and widely used in the analysis of experimental data concerning product formation, biomass formation and substrate consumption (Erickson, 1980; Layokun et al., 1985; Solomon et al., 1985, 1994). Also, the role of statistical techniques in data analysis and parameter estimation in biological system is widely gaining recognition (Yang et al., 1984; Solomon et al., 1984, 1985; Layokun et al., 1985; Beniwal and Chhokar, 2010). This is because measurement errors make accurate estimation of yield and maintenance parameters a difficult task (Layokun et al., 1985). Erickson and other researchers (Erickson, 1980; Solomon et al., 1984; Yang et al., 1984) have identified some multivariate statistical procedures which can be used to improve the quality of estimated parameters by making use of all the measured variables.
Most data in the literature on phenol biodegradation by mixed culture do not lend themselves to energetic analysis using the concept of carbon and available electron balances which have been widely used for data analysis (Erickson, 1980; Ferrer and Erickson, 1979; Oner et al., 1986; Solomon et al., 1984, 1985, 1994, 1995). The reason for this is that the data are incomplete as many variables required are either not measured or reported. Biomass concentration, substrate consumption rates, carbon dioxide production and oxygen uptake rates are considered as measured variables. For a complete data, all these variables ought to be measured to allow for a comprehensive energetic analysis. In spite of the extensive use of phenol biodegradation processes, surprisingly, no work has been published on the estimation of energetic parameters of phenol microbial degradation using a mixed culture and an influent phenol concentration of 100 mg L-1, a level that is lower than what has been investigated by previous studies.
The main objective of this study was to estimate and evaluate or analyze the growth parameters, true growth yields and maintenance coefficients (i.e., energetic parameters) from a complete biodegradation data obtained on the aerobic degradation of phenol by binary mixed culture of Pseudomonas aeruginosa and Pseudomonas fluorescence in chemostat culture. The data used for the estimation include parameters that were measured at various dilution rates: biomass concentration, substrate consumption rates and carbon dioxide production and oxygen uptake rates. In this work, the multivariate statistical technique which has been referred to as the covariate adjustment technique (Solomon et al., 1983, 1994, 1995) was employed in the estimation of true biomass energetic yield (ηmax) and maintenance coefficient (me) associated with the growth of mixed culture of P. aeruginosa and P. fluorescence on phenol. The consistency of the data obtained was examined using the concept of material and energy balance. The analysis should provide accurate estimates of the significant design and model growth parameters, true growth yields and maintenance coefficients. The parameters were estimated using two similar growth models that belong to two different classes. One is Pirt’s model (Pirt, 1965), which assumes that substrate uptake is a consequence of growth. The second model is a modified form of Pirt’s model (Solomon et al., 1994) which assumes that growth is a consequence of substrate uptake.
MATERIALS AND METHODS
Materials: The microorganisms, Pseudomonas aeruginosa and Pseudomonas fluorescence being indigenous bacteria strains isolated from an oil-polluted area in Niger-Delta region of Nigeria were procured from the Department of Microbiology, Obafemi Awolowo University, Ile-Ife, Nigeria. The microorganisms were maintained on nutrient agar slant and stored at 4±1°C for further use. The study was conducted in the Biochemical Engineering Research Laboratory, Department of Chemical Engineering, Obafemi Awolowo University, Ile-Ife, in 2008.
Methods
Preparation of culture medium and inoculum: The mineral salt medium used was a modified form of the one used by Bettmann and Rehm (1984). The mineral salt medium was composed of the following per litre: 700 mL deionized water, 100 mL buffer solution A, 100 mL solution B, 50 mL solution C and 50 mL solution D. Composition of each solution was as follows: Buffer solution A composition: K2HPO4 1.0 g; KH2PO4 0.5 g; (NH4)2SO4 0.5 g; Solution B composition: NaCl 0.5 g; CaCl2 0.02 g; MnSO4 0.02 g; CuSO4.5H2O 0.02 g; H3BO3 0.01 g; Solution C composition: Mg SO4.7H2O 0.5 g; Solution D composition: FeSO4 0.02 g; Molybdenum powder 0.02 g. To prevent the precipitation of CaSO4 and MgSO4 in storage, the medium was prepared by autoclaving water, buffer solution A, solution B, Solution C and Solution D. After cooling the four solutions was mixed together and kept as stock solution from which known quantities was taken for the cultivation of microorganisms.
A primary culture of P. aeruginosa and P. fluorescence was each prepared by transferring two loop full of microorganisms from an agar slant culture (using an inoculating loop which has been sterilized by heating to redness in a spirit flame and allowed to cool by oscillating briefly in air) into 100 mL of feed medium (Nutrient medium) containing 20 mL of mineral salt medium and 80 mL of phenol solution (50 mg L-1) in a two different 250 mL Erlenmeyer conical flask. Each flask was then placed in a New Brunswick gyratory incubator shaker (G25-R model, N.J.S.A) and incubated for 48 h at a temperature of 30°C and agitated with a speed of 120 rpm.
A secondary culture was prepared by transferring 10 mL of each primary culture into 100 mL of feed medium (made up of 20 mL mineral salt medium and 80 mL of phenol solutions) in a two different 250 mL Erlenmeyer conical flask. Each flask was placed in the gyratory incubator shaker and the incubation process was repeated. The secondary culture was used as the inoculum for the cultivation or fermentation experiments as this ensures that the microorganism had fully adapted to growth on the phenol as sole source of carbon and energy. Each of the secondary culture (P. aeruginosa and P. fluorescence) was then mixed in the ratio 1:1 (v:v) which was used as the binary mixed culture for the studies.
Continuous cultivation in bioreactor: The continuous culture was cultivated in a 7½ L capacity bioreactor (New Brunswick Microferm Twin Fermentor, N.J., USA) with a working volume of 4 L. The feed medium was fed into the bioreactor by 101U/R peristaltic pump (Watson-Marlow, England). And the effluent (culture broth) was collected through the side arm into a reservoir or harvest vessel. The culture volume in the bioreactor was constantly maintained by means of a constant level overflow which is located at the side of the fermentor growth vessel. The mass flow of the feed medium was controlled by a balance.
To start the continuous runs, a batch culture was initiated by addition of 200 mL of the inoculum into the fermentor growth vessel containing 800 mL of mineral salt medium and 3 L of phenol solutions (50 mg L-1). Just after the exponential growth has ended, continuous pumping of feed medium containing 100 mg L-1 of phenol was started and smoothly increased for several hours until the required dilution rate was reached. All cultivations were carried out at a temperature of 30°C while the pH was maintained at 7.0 being adjusted by the addition of 1.0 M sodium hydroxide solution and 0.5 M sulphuric acid solution when necessary. Aeration was done with compressed air at a flow rate of 3.0 vvm and the stirrer speed (agitation) was set at 300 rpm. Fifty milliliter samples were withdrawn from the harvest vessel for phenol and biomass determination only when steady state has been established and this occurred after the system has operated for several hydrodynamic residence times (τ = D-1). In order to establish a steady state, the fermentor was left to equilibrate over three to five hydrodynamic residence times and steady state was assumed when the absolute difference in consecutive measurements of biomass and phenol concentrations differed by less than 3%. The exhaust gas from the reactor was analyzed for carbon dioxide and oxygen content using the Infra Red (IR) absorption for CO2 and the paramagnetic properties of oxygen for O2 measurement. The dilution rate was varied between 0.01 h-1 and the washout point and the corresponding steady state data was recorded. The undegraded phenol was estimated quantitatively by measuring its absorbance at a wavelength of 510 nm using UV-visible Spectrophotometer (Lambda 35, Perkin-Elmer, USA) and 4-aminoantipyrene as colour indicator (Yang and Humphrey, 1975). The biomass concentration was estimated using the dry weight method (Agarry et al., 2008).
Methods of data analysis
Consistency tests of experimental data: When phenol is oxidatively converted to biomass with concomitant carbon dioxide and water production as the only other end products, the growth process can be represented stoichiometrically as:
| (1) |
where, ChmOl and ChpOnNq represent the elemental compositions of the organic substrate (phenol in this case) and biomass, respectively. The carbon and available electron balances on Eq. 1 yield Eq. 2 and 3, respectively (Solomon et al., 1994):
| (2) |
| (3) |
where, yc and d are fractions of substrate carbon incorporated into biomass and that which is evolved as carbon dioxide, respectively; η and ε are fractions of substrate energy utilized in biomass formation and heat evolution, respectively. Equation 2 and 3 may be used to check the consistency of data as earlier reported (Solomon et al., 1984, 1985, 1994; Layokun et al., 1985).
For chemostat operation, where D = μ:
| (4) |
| (5) |
| (6) |
| (7) |
where, MCO2 is molecular weight of CO2 (gg mol-1); MO2 is molecular weight of O2 (gg mol-1); OCO2 is rate of OCO2 production (mg L-1 h-1); QO2 is rate of O2 uptake (mg L-1 h-1). S is s ubstrate concentration, subscripts 0 and 1 stand for inlet and outlet respectively (mg L-1); X is biomass concentration (mg L-1); γ is reductance degree (equivalents of available electrons per gram atom carbon)(subscripts ‘b’ and ‘s’ stand for biomass and substrate); μ is specific growth rate (hσ1); σ, mass fraction carbon (subscripts ‘b’ and ‘s’ stand for biomass and substrate) and D is dilution rate (h-1).
Estimation of true yield (ηmax and Ymax) and maintenance coefficient (me): Pirt’s model (Pirt, 1965, 1982) for growth processes has been written in the following forms: based on specific rates of substrate consumption (rs), oxygen uptake (RO2) and carbon dioxide production (rCO2), respectively:
| (8a) |
| (9a) |
| (10a) |
where, ms is maintenance coefficient due to substrate; mO2, maintenance coefficient due to oxygen; mCO2, maintenance coefficient due to carbon dioxide and Ymax with subscripts x/s, x/o2 and x/co2 is true yield due to substrate, oxygen and carbon dioxide, respectively.
These equations have been reparameterized in energetic terms and shown to be correspondingly equivalent (Solomon et al., 1994) to:
| (8b) |
| (9b) |
| (10b) |
where, ηmax is true biomass energetic yield; me, maintenance coefficient due to available electrons; ek1 (k = 1, 2, 3) are correlated error terms with mean 0 and variance-covariance matrix φ and n is the number of observations.
The estimates of ηmax and me are the averages of individual estimates from Eq. 8b-10b. However, combined estimates of the true biomass energetic yield, ηmax and maintenance coefficient, me can be obtained using Eq. 11 as given below:
| (11) |
Nonetheless, the information contained in X1i, X2i and X3i may not be efficiently utilized (Yang et al., 1984; Layokun et al., 1985). Thus, by application of the covariate adjustment technique (Solomon et al., 1985, 1994, 1995) in which appropriate chosen set of covariates Z1i and Z2i which have expected values of zero and are linear function of X1i, X2i and X3i are included in the above Eq. 11, thereby, a better estimate may be obtained. Hence, the model (Yang et al., 1984; Layokun et al., 1985), Eq. 12 is preferred:
| (12) |
In this study, we assume the full set of linearly independent covariates that have zero means as:
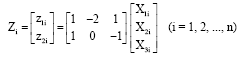 |
Although, due to measurement errors (which lead to data inconsistencies), the values of Z1i and Z2i are not usually zero. By using this full set of covariates, the least square estimates of ηmax and me based on model Eq. 12, then becomes the maximum likelihood estimates (that is estimate with minimum variance), based on the combined models, Eq. 8b, 9b and 10b.
However, maximum likelihood estimates may not the best estimates as in cases where covariates which are uncorrelated with ![]() are excluded. In such cases, the residual variance of model (Eq. 12) is not decreased but that of the degree of freedom for fitting the model, because the covariates included contain no information about X1 (Layokun et al., 1985). Therefore, a measure of goodness of a set of covariates that should be included in Eq. 12 is J = σ2'(n-r-c-1) where σ2 is the mean square error for fitting the model, r is the number of parameters of interest, c is the number of covariates included in the model and n is the number of observations. For this study, r = 2 and 0≤c≤2 and selection of the best estimate is based on the value of σ2/(n-3-c) which is a measure of the goodness of the set of covariates that are included in Eq. 12. The lowest value of J was considered the best for fitting the model for the range when no covariate is included to when both covariates are included. Further details of this statistical method are found in Solomon et al. (1984) and Yang et al. (1984). Nonetheless, the above equations are based on Monod kinetics, which is:
are excluded. In such cases, the residual variance of model (Eq. 12) is not decreased but that of the degree of freedom for fitting the model, because the covariates included contain no information about X1 (Layokun et al., 1985). Therefore, a measure of goodness of a set of covariates that should be included in Eq. 12 is J = σ2'(n-r-c-1) where σ2 is the mean square error for fitting the model, r is the number of parameters of interest, c is the number of covariates included in the model and n is the number of observations. For this study, r = 2 and 0≤c≤2 and selection of the best estimate is based on the value of σ2/(n-3-c) which is a measure of the goodness of the set of covariates that are included in Eq. 12. The lowest value of J was considered the best for fitting the model for the range when no covariate is included to when both covariates are included. Further details of this statistical method are found in Solomon et al. (1984) and Yang et al. (1984). Nonetheless, the above equations are based on Monod kinetics, which is:
| (13) |
where, μmax is the maximum specific growth rate (h-1) and Ks, the half-saturation constant (mg L-1). Equation 13 requires a well defined substrate consumption rate. However, in many cases, growth of microbes is a consequence of substrate consumption and not vice versa (Sonnleitner and Kappeli, 1986; Solomon et al., 1994). It has been shown that in this approach as S tends to 0, μ = 0 and yet a finite quantity of substrate consumption, me is required (that is due to maintenance) (Solomon et al., 1994). Hence, there is a substrate consumption even for S = 0 which is physiologically impossible. Also the substrate consumption is the limiting step and the microorganism’s growth actually follows substrate availability; therefore, instead of Eq. 13, a model of the form:
| (14) |
where, rsmax is the maximum specific substrate consumption rate (mg/mg/h). Equation 14 makes more biological as well as mathematical sense. Therefore, in place of Eq. 8a to 10a, the following would become valid:
| (15a) |
| (16a) |
| (17a) |
The Eq. 15a to 17a have been reparameterized in energetic terms and are shown to be correspondingly equivalent (Solomon et al., 1994) to:
| (15b) |
| (16b) |
| (17b) |
where, m' = -me ηmax. The values of m' have the same dimension as μ mathematically and hence cannot be referred to as the maintenance. They may be described as specific death rates and physiologically as energy not available for growth (Solomon et al., 1994). Equations 15b to 17b were also used to estimate ηmax and m'e.
RESULTS AND DISCUSSION
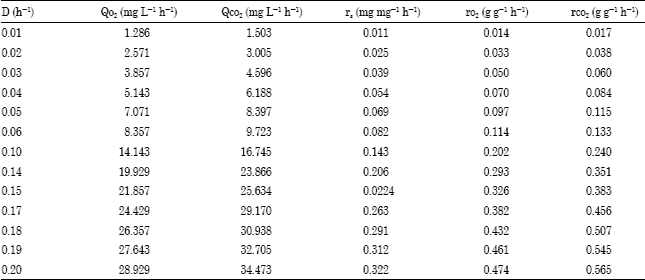
Data consistency test: The calculated values of phenol consumption rates (Qs), oxygen uptake rate (QO2) and carbon dioxide production rate (QCO2) (Table 1) were used for the estimation of the biomass energetic yield (η) and carbon yield (ye) for the binary mixed culture of P. aeruginosa and P. fluorescence using the carbon and available electron balances as given in Eq. 4-7. For the estimation, the average values of σb = 0.490 and γb = 4.793 which have been calculated from the measured composition of Pseudomonas species obtained by Layokun (1982) were used. The instantaneous available electron and carbon balances results obtained are presented in Table 2. From Table 2, it could be seen that the biomass energetic yield (η) and carbon yield (yc) are low (i.e., less than 1) which thus agree with the available electron and carbon balance equation.
| Table 1: | Calculated oxygen and carbon dioxide transfer rates for the continuous degradation of phenol by binary mixed culture of P. aeruginosa and P. fluorescence |
 | |
| Table 2: | Examination of data consistency using instantaneous available electron and carbon balances for the growth of binary mixed culture of P. aeruginosa and P.fluorescence in phenol-limited continuous culture (chemo stat operation) |
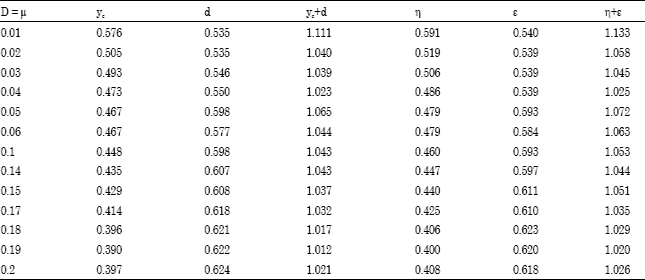 | |
It could also be seen from Table 2 that both the biomass energetic yield (η) and carbon yield (yc) decreased as the dilution rate increased. This observation may be due to decrease in steady state biomass concentration and decrease in phenol removal at high dilution rate (Agarry et al., 2008). Consistency tests (checks) were made using Eq. 2-3. It has been established (Solomon et al., 1981) that in consistency analysis allowance has to be made for deviation from the ideal. The parameters by which consistency is defined should satisfy 0.94≤ (yc+d) ≤1.06 and 0.93≤(η+ε)≤ 1.07. The results of the data consistency tests are as shown in Table 2. Thus, it could be seen from Table 2 that the consistency equations are generally satisfied. Also, it could be seen from Table 2 that the (yc-d) and (η+ε) values generally decreased as the dilution rate increased. This is in agreement with the report of Solomon et al. (1995). Generally, therefore, the consistency tests suggest that in phenol-limited chemostat culture, the binary mixed culture of P. aeruginosa and P. fluorescence were able to oxidatively metabolized phenol to carbon dioxide and water with concomitant biomass production.
| Table 3: | Estimates of true biomass growth yields and maintenance coefficient for the growth of mono and mixed culture of Pseudomonas species in phenol-limited continuous culture using Pirt’s model (Eq. 8a-10a) |
 | |
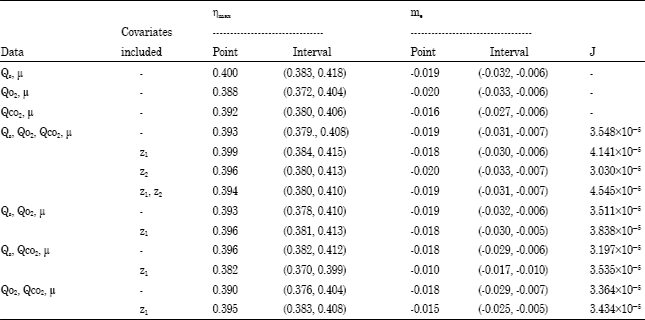
| Table 4: | Estimates of true biomass energetic yields and maintenance coefficient for the growth of binary mixed culture of P. aeruginosa and P. fluorescence in phenol-limited continuous culture using Pirt’s model (Eq. 8b-10b) |
 | |
Estimation of true yield and maintenance coefficient: Pirt”s model for growth as given in Eq. 8a to 10a was used to estimate the true yields and maintenance coefficients in terms of substrate, oxygen and carbon dioxide. The calculated specific rates of phenol consumption (rs), oxygen uptake (rO2) and carbon dioxide production (rCO2) obtained for the binary mixed culture were plotted as a function of dilution rate (D) to obtain the true growth yield and maintenance coefficients, respectively. The estimated values are given in Table 3.
The Pirt’s model was reparameterized to produce multiresponse models with common parameters as given in Eq. 8b to 10b and application of covariate adjustment technique (Solomon et al., 1994) to these equations resulted in a unit variate linear model with covariates. These allow combined point and interval estimates of biomass energetic yield and maintenance coefficient to be obtained using standard multiple regression programs. Therefore, using Eq. 8b to 10b, various estimates of the true biomass energetic yield and maintenance coefficients based on the data in Table 1 were obtained for the binary mixed culture and are presented in Table 4. The first three estimates in Table 4 are the individual least square estimates using substrate and biomass data and Eq. 8b and oxygen and biomass data and Eq. 9b and carbon dioxide and biomass data and Eq. 10b, respectively. These estimates are quite comparable but differ because of measurement errors. At low dilution rates, measurement errors are usually significant because of low gas consumption and production rates which cannot be measured accurately (Solomon et al., 1995).
When all the measured data were used (i.e. Qs, Qo2, Qco2, μ) the best estimate was the Maximum Likelihood Estimate (MLE) which corresponded to when one covariate (Z2) was included. This was based on the lowest value of J = 3.030x10-5. The respective combined point estimates for ηmax and me were 0.396 and -0.020 h-1 with the corresponding 95% confidence intervals (0.380, 0.413) and (-0.033, -0.007) h-1. When the carbon dioxide data were excluded (i.e.,Qs, Qo2, μ were used), then the respective best point and interval estimates for ηmax were 0.393 and (0.378, 0.410) and the me are -0.019 h-1 and (-0.032, 0.006) h-1. With the oxygen data excluded (i.e., Qs, Qco2 were used), ηmax = 0.396 with interval (0.382, 0.412) and me = -0.018 h-1 with interval (-0.029,-0.006) h-1. Lastly, when substrate measurements were excluded (i.e. Qo2, Qco2 were used), ηmax = 0.392 with interval (0.376, 0.404) and me = -0.018 h-1 with interval (-0.029, -0.007) h-1.
For the mixed culture of organisms studied, even though the respective values of these combined point estimates were different from one another, all the 95% confidence intervals were overlapping and included all the point estimates. Generally, based on the least measure of goodness of fit value, the best estimate was obtained when J = 3.030x10-5 which was for the case when all the measured data were used and corresponded to the Maximum Likelihood Estimate (MLE) value of ηmax = 0.396 with 95% confidence intervals (0.380, 0.413) and me = -0.020 h-1 with interval (-0.033, 0.007) h-1. In earlier applications of this procedure (Solomon et al., 1981,1983) the best combined estimates was always assumed to be obtained when all the measured data were used. The results obtained for binary mixed culture (P. aeruginosa and P. fluorescence) have shown that a combined estimate from all the measured data might in fact lead to a better estimate. This is in agreement with the observations of Layokun et al. (1985) and Solomon et al. (1994, 1995) when all the measured data was used.
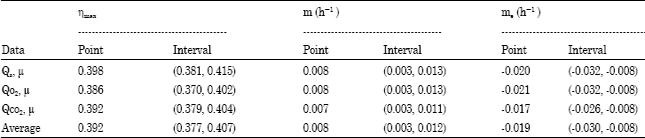
The estimates of ηmax and me using the modified Pirt model (Eq. 15b-17b) and the data in Table 1 are presented in Table 5. For these cases, only the individual estimates have been made because the covariate adjustment technique was not suitable. However, there was good agreement between the corresponding individual estimates for the two cases (Pirt’s model and the modified Pirt’s model) as shown in Table 4 and 5, respectively. The most reliable estimate in Table 5 was the average which gave ηmax = 0.392 and me =-0.019 h-1 with the respective 95% confidence interval (0.377, 0.407) and (-0.030, -0.008) h-1. The individual estimates (ηmax and me) obtained for the binary mixed culture of P. aeruginosa and P. fluorescence using modified model equation (Eq. 15b-17b) were found to be lower than the individual estimates obtained using the reparameterized Pirt’s model. A similar observation has been reported by Solomon et al. (1994, 1995). The estimates of me in Tables 4 and 5 are statistically significantly lower than zero and therefore negligible. Hill and Robinson (1975) reported that the maintenance coefficient for phenol degradation is negligible.
The true yields and maintenance coefficients in terms of substrate, oxygen and carbon dioxide were obtained using the modified model. The true yield (Ymax) and maintenance coefficient in terms of substrate, oxygen and carbon dioxide obtained using the modified model were found to be slightly lower than the individual estimates obtained when Pirt’s model was used (Table 6).
| Table 5: | Estimates of true biomass energetic yields and maintenance coefficient for the growth of binary mixed culture of P. aeruginosa and P. fluorescence in phenol-limited continuous culture using modified Pirt’s model (Equations 15b-17b) |
 | |
| Table 6: | Summary of true biomass growth yields and maintenance coefficient for the growth of binary mixed culture of P. aeruginosa and P. fluorescence in phenol-limited continuous culture |
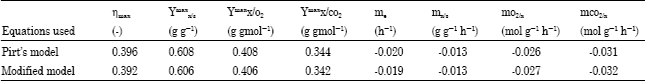 | |
The true biomass energetic and growth yield obtained for the binary mixed culture of P. aeruginosa and P. fluorescence was found to be higher than that obtained for the monoculture of P. fluorescence (ηmax = 0.262 and Ymaxx/s = 0.463) and P. aeruginosa (ηmax= 0.359 and Ymaxx/s = 0.540) (Agarry, 2009). However, it is lower than that obtained for monoculture of P. cepacia G4 (ηmax = 0.432 and Ymaxx/s = 0.785) by Solomon et al. (1994).
CONCLUSIONS
The advantage of combined estimates using covariate adjustment technique has been demonstrated. This analysis showed that with a combined use of material and energy balances and statistical procedure, discrimination may be made between various variables to identify those with more errors. The results demonstrated that the Pirt’s model approach (based on Monod kinetics) which require well-defined substrate consumption as well as the modified approach which assumed that substrate consumption was rate limiting were similar and led to similar estimates However, the latter approach did not allowed the application of a multivariate statistical method for parameter estimation.
From this analysis, about 38-41% of the energy in phenol may be incorporated into binary mixed culture of P. aeruginosa and P. fluorescence biomass, while the balance, 58.7-62% is mostly evolved as heat with little energy use for maintenance of the cells. When compared with the degradation of phenol by P. cepacia G4 (Solomon et al., 1994), the energetics were similar. However, less substrate energy was incorporated into biomass and hence more evolved as heat in the case of binary mixed culture of P. aeruginosa and P. fluorescence. This might probably be due to differences in ATP production and utilization in the processes (Solomon et al., 1995). The combined estimates, which seems to be an improvements on the estimates made from individual measurements are the values most likely to be used when true biomass energetic yield and maintenance coefficients are applied to the design of fermentors.
REFERENCES
- Afzal, M., S. Iqbal, S. Rauf and Z.M. Khalid, 2007. Characteristics of phenol biodegradation in saline solutions by monocultures of Pseudomonas aeruginosa and Pseudomonas pseudomallei. J. Hazard Mater, 149: 60-66.
PubMed - Agarry, S.E. and B.O. Solomon, 2008. Inhibition kinetics of phenol degradation by Pseudomonas aeruginosa from continuous culture and washout data. Bioremed. J., 12: 12-20.
CrossRef - Agarry, S.E., B.O. Solomon and S.K. Layokun, 2008. Substrate inhibition kinetics of phenol degradation by binary mixed culture of Pseudomonas aeruginosa and Pseudomonas fluorescence from steady state and washout data. Afr. J. Biotechnol., 7: 3927-3933.
Direct Link - Agarry, S.E., B.O. Solomon and T.O.K. Audu, 2010. Optimization of process variables for the batch degradation of phenol by Pseudomonas fluorescence using response surface methodology. Int. J. Chem. Technol., 2: 33-45.
CrossRefDirect Link - Beniwal, V. and V. Chhokar, 2010. Statistical optimization of cultural conditions for tannase production by Aspergillus awamori MTCC9299 under submerged fermentation. Asian J. Biotechnol., 2: 46-52.
Direct Link - Bettmann, H. and H.J. Rehm, 1984. Degradation of phenol by polymer entrapped microorganisms. Applied Microbial. Biotechnol., 20: 285-290.
CrossRef - Ferrer, A. and L.E. Erickson, 1979. Evaluation of data consistency and estimation of yield parameters in hydrocarbon fermentations. Biotechnol. Bioeng., 21: 2203-2233.
CrossRefDirect Link - Folsom, B.R., P.J. Champan and P.H. Pritchard, 1990. Phenol and Trichloroethylene degradation by Pseudomonas cepacia G4: Kinetics and interactions between substrates. Applied Environ. Microbial., 56: 1279-1285.
Direct Link - Hill, G.A. and C.W. Robinson, 1975. Substrate inhibition kinetics: Phenol Degradation by Pseudomonas putida. Biotechnol. Bioeng., 17: 599-615.
CrossRef - Layokun, S.K., B.O. Solomon and I.A. Fatile, 1985. Data consistency, yield and maintenance coefficient for the growth of Candida lipolytica and Pseudomonas aeruginosa on n-hexadecane. Applied Microbiol. Biotechnol., 21: 368-373.
CrossRefDirect Link - Li, Y., J. Li, C. Wang and P. Wang, 2010. Growth kinetics and phenol biodegradation of psychrotrophic Pseudomonas putida LY1. Bioresour. Technol., 101: 6740-6744.
PubMed - Nikakhtari, H. and G.A. Hill, 2006. Continuous bioremediation of phenol-polluted air in an external loop airlift bioreactor with a packed bed. J. Chem. Technol. Biotechnol., 81: 1029-1038.
CrossRef - Oboirien, B.O., B. Amigun, T.V. Ojumu, O.A. Ogunkunle, O.A. Adetunji, E. Betiku and B.O. Solomon, 2005. Substrate inhibition kinetics of phenol degradation by Pseudomonas aeruginosa and Pseudomonas fluorescence. Biotechnology, 4: 56-61.
CrossRefDirect Link - Oner, M.D., L.E. Erickson and S.S. Yang, 1986. Estimation of the true growth yield and maintenance coefficient for yoghurt cultures. Biotechnol. Bioeng., 28: 919-926.
PubMedDirect Link - Paller, G., R.K. Hommel and H.P. Kleber, 1995. Phenol degradation by Acinetobacter calcoaceticus NCIB 8250. J. Basic Microbiol., 35: 325-335.
PubMedDirect Link - Pawlowsky, U., J.A. Howell and C.T. Chi, 1973. Mixed culture biooxidation of phenol. III: Existence of multiple steady states in continuous culture with wall growth. Biotechnol. Bioeng., 15: 905-916.
CrossRefDirect Link - Pirt, S.J., 1965. The maintenance energy of bacteria in growing cultures. Proc. R. Soc. London Ser. B, 163: 224-231.
Direct Link - Pirt, S.J., 1982. Maintenance energy: A general model for energy-limited and energy-sufficient growth. Arch. Microbiol., 133: 300-302.
CrossRef - Amer, R.A., M.M. Nasier and E.R. El-Helow, 2008. Biodegradation of monocyclic aromatic hydrocarbons by a newly isolated Pseudomonas strain. Biotechnology, 7: 630-640.
CrossRefDirect Link - Ruiz-Ordaz, N., J.C. Ruiz-Lagunez, J.H. Castanon-Gonzalez, E. Hernandez-Manzano, E. Cristiani-Urbina and J. Galindez-Mayer, 2001. Phenol biodegradation using a repeated batch culture of Candida tropicalis in a multistage bubble column. Revista Latinoamericana de Microbiologia, 43: 19-25.
Direct Link - Schroeder, M., C. Muller, C. Posten, W.D. Deckwer and V. Hecht, 1997. Inhibition kinetics of phenol degradation from unstable steady state data. Biotechnol. Bioeng., 54: 567-576.
Direct Link - Solomon, B.O., L.E. Erickson and J.L. Hess, 1981. Application of data consistency tests and new parameter estimation methods to microbial growth on corn dust in batch culture. Biotechnol. Bioeng., 23: 2333-2360.
CrossRefDirect Link - Solomon, B.O., L.E. Erickson and S.S. Yang, 1983. Utilisation of statistics and experimental design in data collections and analysis. Biotechnol. Bioeng., 25: 2683-2705.
PubMedDirect Link - Solomon, B.O., M.D. Oner, L.E. Erickson and S.S. Yang, 1984. Estimation of parameters where dependent observations are related by equality constraints. AIChE J., 30: 747-757.
CrossRefDirect Link - Solomon, B.O., S.K. Layokun, I.A. Fatile and G.N. Agho, 1985. Analysis of the growth of Trichosporon cutaneum on glucose: Yield and maintenance requirements. J. Chem. Tech. Biotechnol., 35: 266-272.
CrossRefDirect Link - Solomon, B.O., C. Posten, M.P.F. Harder, V. Hecht and W.D. Deckwer, 1994. Energetics of Pseudomonas cepacia growth in a chemostat with phenol Limitation. J. Chem. Technol. Biotechnol., 60: 275-282.
CrossRef - Solomon, B.O., V. Hecht, C. Posten and W.D. Deckwer, 1995. Estimation of the energetic parameters associated with the aerobic degradation of quinoline by Comamonas acidovorans in continuous culture. J. Chem. Tech. Biotechnol., 62: 94-97.
Direct Link - Sonnleitner, B. and O. Kappeli, 1986. Growth of Saccharomyces cerevisiae is controlled by its limited capacity formulation and verification of a hypothesis. Biotechnol. Bioeng., 28: 927-937.
PubMedDirect Link - Udayasoorian, C., P.C. Prabu and G. Balasubramanian, 2007. Degradation of pentachlorophenol by white rot fungus (Phanerochaete chrysosporium-TL 1) grown in ammonium lignosulphonate media. Biotechnology, 6: 76-80.
CrossRefDirect Link - Yang, R.D. and A.E. Humphrey, 1975. Dynamic and steady state studies of phenol biodegradation in pure and mixed cultures. Biotechnol. Bioeng., 17: 1211-1235.
CrossRefPubMedDirect Link - Yang, S.S., B.O. Solomon, M.D. Oner and L.E. Erickson, 1984. A method of estimation and testing common parameters for some multiresponse models associated with growth and bioenergetics. Technometrics, 26: 355-361.
Direct Link