
Wisam Nabeel Ibrahim
Department of Basic Medical Sciences for Nursing, Faculty of Nursing, International Islamic University Malaysia, Kuantan, Pahang, Malaysia
LiveDNA: 964.16203
Ridhwan Abdul Wahab
Department of Biomedical Sciences, Faculty of Allied Health Sciences, International Islamic University Malaysia, Kuantan, Pahang, Malaysia
Mohammad Syaiful Bahari Bin Abdull Rasad
Department of Biomedical Sciences, Faculty of Allied Health Sciences, International Islamic University Malaysia, Kuantan, Pahang, Malaysia
International Journal of Cancer Research
Year: 2017 | Volume: 13 | Issue: 1 | Page No.: 17-25







ABSTRACT
Background and Objective: Fibroblast stromal cells actively participates in tumor invasion by secreting matrix metalloproteinases (MMPs) within the tumor microenvironment. Expression of these enzymes is primarily regulated at a transcriptional level via interaction with transcription factors. Among these factors, YB-1 oncogenic factor binds with different nucleic acids to exert its diverse influences. Also it represents an important prognostic indicator in many types of tumors. The aim of this study was to assess the expression of collagenases MMPs (MMP1, MMP8, MMP13) and the cellular proliferation in one of the most invasive types of cancer cell types which is the A375 malignant melanoma cancer cell line using co-culture settings. Also, the study attempted to assess the expression of YB-1 factor and its in vivo interaction with the AP-1 gene promoter sequence. Materials and Methods: The experiment involved growing A375 cells with CCD1079SK fibroblasts cells in co-culture environment. The proliferation of cells was determined using serial trypan blue assays, while the expression of YB-1, MMP1, MMP8 and MMP13 was determined by the use of real-time PCR and western blotting analysis. The potential interaction between YB-1 protein and AP-1 promoter sequence was assessed through chromatin immunoprecipitation (ChIP) assay. SPSS with independent t- test was used to compare cell proliferation and real-time PCR Ct mean values between samples. Results: In co-culture setting, the proliferation of A375 cancer cells was significantly faster than the cells in monoculture setting (5.1×105, 3×105 respectively in day 3) (p<0.05). Also, there was a significant increase in the expression of MMP1 enzyme. YB-1 and MMP8 were significantly expressed more in the A375 cancer cells in comparison with normal fibroblasts cells (p<0.05). Conclusion: The study confirms the role of stromal fibroblasts by enhancing the proliferation of melanoma cancer cells in vitro and increasing the expression of the MMP1 enzyme. In addition, YB-1 factor remains as an important prognostic indicator in cancer that might regulate expression of MMPs without binding to the AP-1 promoter sequence.
PDF Abstract XML References Citation
Received: October 10, 2016;
Accepted: November 18, 2016;
Published: December 15, 2016
How to cite this article
Wisam Nabeel Ibrahim, Ridhwan Abdul Wahab and Mohammad Syaiful Bahari Bin Abdull Rasad, 2017. Expression of Collagenases Matrix Metalloproteinases and YB-1 Oncogenic Factor in Malignant Melanoma Cancer Cells and its Regulation by Stromal Fibroblasts. International Journal of Cancer Research, 13: 17-25.
DOI: 10.3923/ijcr.2017.17.25
URL: https://scialert.net/abstract/?doi=ijcr.2017.17.25
DOI: 10.3923/ijcr.2017.17.25
URL: https://scialert.net/abstract/?doi=ijcr.2017.17.25
INTRODUCTION
The tumor microenvironment is an evolving crosslink between tumor cells and the adjacent normal interstitial cells such as fibroblasts, endothelial and some immune cells1. Cancer cells recruit these stromal cells via various interactions and change their function within tumor tissue to promote their expansion and spread1. These interactions have been validated in many types of malignancies, most notably lung, breast, prostatic, pancreatic and skin cancer2,3. Among these cells, the normal spindle-shaped fibroblasts cells are important in maintaining the framework of extracellular matrix (ECM) by synthesizing collagens, proteoglycans and fibronectin in addition to the release of protease enzymes capable of degrading the ECM4. However, in the tumor stroma, these cells also known as Cancer-Associated Fibroblast (CAF) cells are perpetually activated and neither revert to a normal phenotype nor undergoes apoptosis or elimination5. The CAF cells communicate with cancer cells through a number of growth factors like vascular endothelial growth factor, platelet-derived growth factor, fibroblast growth factor, transforming growth factor and interleukin-66,7 in addition to their direct interaction with adjacent cancer cells to facilitative their proliferation, extravasations, invasion, metastasis and angiogenesis8. In these interactions, CAFs are believed to break down tissue basement membranes by releasing the matrix metalloproteinases (MMPs) which are a group of extracellular zinc-dependent endopeptidase enzymes, classified to several sub-groups including collagenases, gelatinases, stromelysins and matrilysins9,10. Collagenases include MMP1, MMP8 and MMP13 which are capable of initiating degradation of native fibrillar collagens (type I, II, III, V and IX) and play a vital role in embryonic tissue development and tissue remodeling11. These enzymes are also highly expressed in many malignancies12-14. For example, MMP1 is highly expressed in highly invasive malignant melanoma together with MMP1310,15. Similarly, MMP8 is highly expressed in breast cancer and number of ovarian cancer cell lines16, 17. The transcriptional regulation of these collagenases involves activated cancer pathways such as Mitogen-Activated Protein Kinase (MAPK) pathway18. Also, several transcription factors are found to indirectly regulate collagenases expression through interaction with specific gene promoter sequences such as AP-1 site19. The Activator Protein-1 (AP-1) DNA binding site 5'TGAGTCA’3 is an important binding site for many oncogenic factors for the transcriptional regulation of MMPs expression19. The AP-1 transcription factors also induces the expression of different factors related to enhancement of cancer cell proliferation and progression20.
The YB-1 oncogenic factor is a transcription factor that was reported to bind with AP-1 site21. This factor is a member of the cold-shock protein family containing a highly conserved nucleic-acid-binding motif responsible for DNA repair, RNA splicing and exon skipping21. Samuel et al.22 confirmed the role of this factor in promoting tumor cellular proliferation, invasion, metastasis and drug resistance. Also, YB-1 protein is found to interfere with the commonly activated cancer pathways such as the MAPK pathway, phosphoinositide 3-kinase (PI3K) pathway, mechanistic target of rapamycin (mTOR) pathway and p53 pathways23,24. Therefore, an elevated level of YB-1 protein is considered as a poor prognostic indicator in many malignancies such as breast, lung, bone and the skin cancers25,26. Currently, there are some conflicting evidences about the binding of YB-1 protein to the AP-1 sequence of MMPs gene promoter that show variable influences on MMPs transcription19,21. Therefore, this study aimed to investigate this ambiguity by examining YB-1 expression and YB-1-DNA interaction by chromatin immune precipitation (ChIP) analysis. The effect of stromal cells (fibroblasts) on the proliferation and the expression of MMP1, MMP8, MMP13 by the malignant melanoma cancer cells in the co-culture setting was also studied.
MATERIALS AND METHODS
The study was conducted in the Cell Culture and Molecular Laboratory in the Integrated Centre for Research Animal Care and Use (ICRACU) institute, International Islamic University Malaysia, Kuantan. All of the study laboratory procedures have taken place during the time between November 2016 and March 2017.
Cell culture conditions: The study involved the A375 malignant melanoma (ATCC® CRL-1619) and the CCD1079SK normal human skin fibroblast (ATCC® CRL-2097) cell lines. Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS), 1 mM sodium pyruvate, 10 mM HEPES buffer (pH 7.4) and 100 μg mL–1 penicillin-streptomycin was used to culture both cell lines in culture plate in a humidified incubator (5% CO2, 37°C). Cell lines were used within 15 passages from cryopreservation and all the reagents used were of cell culture grade. The growth rate of A375 cancer cells was optimized and the exponential and plateau growth phases harvesting time were determined at day 4 and day 7, respectively.
Co-culture setting: To determine the effect of stromal fibroblast cells on the proliferation of A375 melanoma cells in vitro, approximately 2×105 CCD1079SK fibroblast cells were seeded within 0.3 mL of DMEM complete media into polyethylene terephthalate (PET) membrane well inserts (Corning, USA), 0.4 μm pore size. The well inserts were kept inside the wells of 24 well plate (Corning, USA) containing 0.9 mL of complete media in each well. The plate was then incubated for 48 h supplemented with 5% CO2 at 37°C. On day 3, approximately 4×105 of A375 cells were isolated and seeded into the lower compartment in the well plate. By this method, the two cell lines were physically isolated from each other but the communication was maintained by the media exchanging through the PET membrane. Also, the same number of A375 cells was seeded into another set of wells with well insert containing only complete media with no cells as a negative control.
Another setting of co-culture was used following the protocol setting from Kim et al.8 with few modifications. In this procedure, we grew nearly 1×104 normal skin fibroblast cells in complete DMEM growth media for approximately five days to reach 60% confluence. Subsequently, approximately 1×104 A375 melanoma cells were seeded on the top of the attached fibroblasts in the same well and incubate both cells for 72 h. Lysates from these cells were used in subsequent real-time PCR and western blot processing and analysis. The preference of this co-culture setting was because the stromal cells were in direct contact with the A375 melanoma cancer cells in addition to the maintenance of the paracrine signaling effects established among the cells in the setting.
Incubation of A375 and CCD1079SK cells monocultures was in the same complete DMEM media and same incubation conditions until the cells reach 90% confluence. At the same time, some monocultures were left in incubation beyond the confluence level to reach the plateau phase of cell growth where the cell proliferation is greatly reduced and most of the metabolic activities in the cancer cells are significantly impaired. These overgrown cells were analyzed for the expression of same target proteins to demonstrate the effect of the less aggressive A375 cancer cells.
Cell quantification: Viable cells were detached using triple E-express (Gibco, USA), after incubation, the cell suspension was neutralized using media supplemented with FBS followed by centrifugation of the cell suspension in a sterile 15 mL tube for 5 min at 2200 rpm. Subsequently, 100 μL of cell suspension diluted with 400 μL of trypan blue solution incubated for 2 min at room temperature followed by cell counting using hemocytometer under the microscope magnification.
Real-time PCR: After detaching A375, CCD1079SK and the co-culture cells using triple E-express (Gibco, USA) followed by washing the cells with PBS buffer (pH 7.4), the mRNA was extracted using Aurum Total RNA Mini kit (BioRad, USA) followed by checking the RNA integrity on denaturing RNA electrophoresis confirming the 18S and 28S bands with no smearing. This was followed by RNA purity test using Nanodrop spectrophotometer (thermoScientific, USA) confirming A260/A280 and A260/A230 ratios being higher than 2.00. Then the RNA samples were normalized and subjected to cDNA formation using iScript™ Advanced cDNA Synthesis Kit (BioRad, USA). The amplification of cDNA samples was implemented using SsoAdvUniverSYBR Green master mix (BioRad, USA) with specific primers sets for each of the target proteins (BioRad, USA).
Gene expression data was analyzed using ΔΔ Ct method after normalizing the Ct mean values with Ct mean value of three housekeeping genes among the samples to determine the relative mRNA expression of each protein. The experiments involved triplicate sets for each of the experiments with multiple replicates.
Western blot analysis: The cell samples were mixed with Pierce RIPA lysis buffer (25 mM Tris-HCl (pH 7.6), 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) (Thermo Fisher, USA) on ice to extract membrane, nuclear and cytoplasmic proteins. The mixtures were subjected to 30 sec of water bath sonication for three times every 10 min while incubation on ice for 30 min followed by centrifugation at 12,000 rpm for 30 min at 4°C. The supernatant was carefully aspirated to a new tube and stored at -80°C. The protein lysates concentration was normalized using pierce bicinchoninic acid (BCA) assay kit (Thermo Fisher, USA) for the colorimetric detection and quantitation of total protein by running the standard curve of different known protein concentrations. After running SDS-polyacrylamide gel electrophoresis for 90 min, the proteins were transferred to polyvinylidene fluoride (PVDF) membrane using semi-dry blotting procedure. After exposing the membranes to blocking solution for 60 min, we incubate the membranes with primary specific monoclonal mouse antibodies (Santa-Cruz, Germany) at 4°C overnight with aggression. Then the membranes were washed five times with Tris-buffered saline with 0.1% Tween 20 (TBST) buffer (pH 7.6) followed by incubation in horseradish peroxidase (HRP)- conjugated secondary Rabbit anti-mouse antibodies solutions (Santa-Cruz, Germany) for 1 h at room temperature. After washing, HRP substrate was added to the membrane and the protein bands were detected and analyzed using ChemiDoc™ XRS (BioRad, USA).
Chromatin immune precipitation: The interaction between YB-1 proteins and the AP-1 gene promoter was determined using SimpleChIP® Plus Enzymatic Chromatin IP Kits (Cell Signaling, USA). In this method, formaldehyde was used to fix the protein-DNA complexes which were then isolated from the cell lysate using YB-1 ChIP-validated monoclonal antibodies (Santa-Cruz, Germany) by using magnetic beads separation. The cross-linking was then reversed to split the DNA from the proteins of interest followed by DNA purification and PCR amplification of AP-1 target sequence using specific primers sequence19. The PCR products were stained with ethidium bromide (Sigma-Aldrich, USA) and visualized on 1% agarose gel (Sigma-Aldrich, USA) by ChemiDoc™ XRS (BioRad, USA).
Statistical analysis: The study quantitative data were recorded as the Mean±Standard Error of the mean value and the statistical analysis was completed using SPSS 17.0 (SPSS Inc., Chicago, IL, USA). Comparison between two groups was done by independent t tests. p<0.05 was considered to indicate a statistically significant difference20.
RESULTS
Cell proliferation: The experiment involved growing equal numbers of A375 cells in monoculture and co-culture as shown in Fig. 1. The proliferation of the cancer cells was significantly increased (p<0.05) in the co-culture setting in comparison with cancer cell monoculture setting. The increased proliferation was also elicited under the microscope especially after 72 h of incubation in cancer cells monocultures and co-cultures as shown in Fig. 1.
Real-time PCR and Western blot analysis: In A375 monocultures, there was a significant decrease (p<0.05) in the relative expression of MMP1 gene in comparison with fibroblasts cells during the exponential and plateau growth phases. Remarkably, in co-culture setting, the relative expression of MMP1 gene was 178-fold significantly increased (p<0.05) in comparison with A375 monocultures (Fig. 2).
The relative expression of MMP8 gene had a (122 and 44)-folds significant up-regulation (p<0.05) in A375 cancer cells monocultures in comparison with fibroblasts cells during the exponential and plateau growth phases respectively. While in co-culture setting there was a significant reduction (p<0.05) in the relative expression of MMP8 in comparison with A375 monocultures. The MMP13 gene did not have any significant changes (p>0.05) in its relative expression between monocultures and co-cultures or between different growth phases.
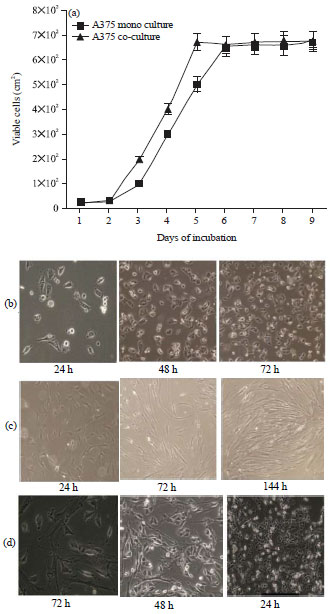 | |
| Fig. 1(a-d): | (a) Growth curve for A35 cells in monoculture and co-culture. (b-d) Microscope graphs (10X magnification) of cell growth morphology. (b) A375, (c) CCD1079SK and (d) co-culture of both cell lines |
The relative expression of YB-1 gene was significantly 6-folds increased (p<0.05) in A375 cancer cells monocultures in comparison with normal fibroblasts cells during the exponential growth phase. However, in the plateau growth phase of monocultures, the changes were reversed where YB-1 gene relative expression was significantly reduced (p<0.05) in A375 melanoma cells in comparison with normal fibroblasts cells. In the co-culture setting, YB-1 gene did not show any significant changes in relative expression compared with A375 cancer cells monoculture (p>0.05).
The protein expression of the target factors is shown in Fig. 3. YB-1 and MMP8 proteins were highly expressed in A375 cells monocultures, while MMP1 was highly expressed in the co-culture setting of A375 and fibroblasts cells.
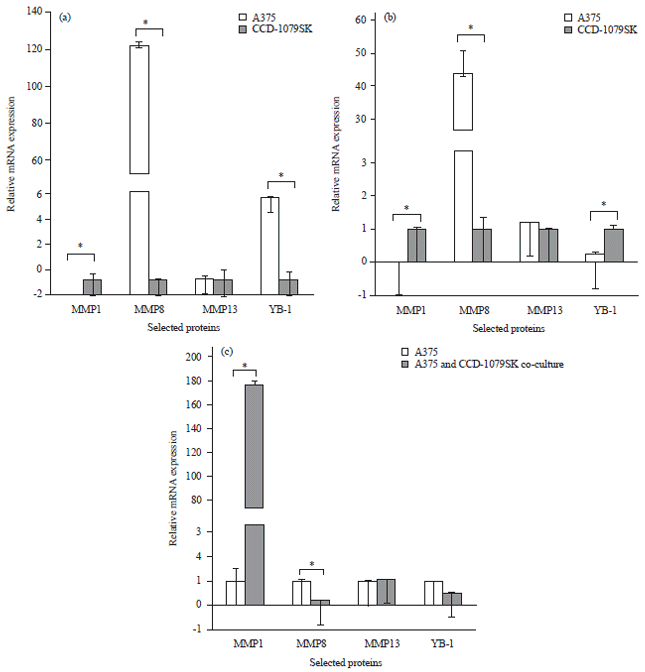 | |
| Fig. 2(a-c): | Relative mRNA expression analysis of target genes showing their mean values±SEM (a) Exponential cell growth of A375 and CCD-1079SK monocultures, (b) Plateau cell growth of A375 and CCD-1079SK monocultures and (c) Exponential cell growth of A375 monoculture and co-culture of A375 and CCD-1079SK *p<0.05 |
The Western blotting also elucidated a reduction in the expression of the YB-1 protein in co-culture setting and fibroblast cells in comparison with A375 melanoma cells monocultures.
Chromatin immunoprecipitation assay: In the study, while investigating the interaction of the YB-1 protein with the AP-1 sequence of the MMP gene in vivo by fixing the cells in the culture environment using ChIP assay. The YB-1 protein did not bind with the AP-1 sequence as shown in Fig. 4. The sequence was validated in the DNA extracted from A375 cells.
DISCUSSION
Fibroblast cells enhance proliferation of melanoma tumor cells in vitro: To elucidate the effect of fibroblast cells on the proliferation of A375 cells, we established the cancer-stromal cell co-culture environment in separate compartments using PET membrane allowing the exchange of biochemical compounds between cells via this membrane.
 | |
| Fig. 3: | Western blot analysis |
1: A375, 2: CCD1079SK, 3: Co-culture (A375+CCD1079SK), The molecular weights of the target proteins were 52, 55 and 48 kDa for MMP1, MMP8 and MMP13, respectively, 55 and 50 kDa for YB-1 and Tubulin- alpha, respectively | |
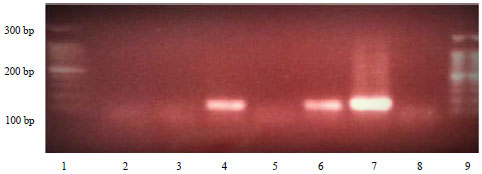 | |
| Fig. 4: | Chromatin immunoprecipitation assay |
Lane 1 and 9: 100 bp ladder, Lane 2: AP-1 site interaction with YB-1, Lane 3: AP-1 site interaction with CTCF, Lane 4: 144 bp product for AP-1 site interaction with +ve control histone antibody, Lane 5: AP-1 site interaction with -ve control anti Rabbit antibody, Lane 6: 2% output that confirms the AP-1 sequence in the DNA isolated, Lane 7: Another 160 bp PCR product showing the interaction of PCR positive control with YB-1, Lane 8: PCR negative control (no DNA sample) | |
As demonstrated in the growth curve in Fig. 1 and section C, there was a marked increase (p<0.05) in the proliferation of A375 melanoma cells by the co-cultured CCD1079SK fibroblast cells. As stated in the review by Shiga et al.5, CAFs interact with cancer cells by their contact and via secretion of different growth factors to enhance tumor cells proliferation and metabolism3,27-29. Due to this, cancer cells are no longer the main contributor in tumor progression. The recruitment of these activated CAF cells in the tumor stroma are associated with higher risk of metastasis and poor life expectancy in many types of cancer such as breast cancer, lung cancer and pancreatic cancer29-31. Several studies conducted on co-culture cells have shown that CAFs stimulated the cancer cell growth in vitro32,33. Also, in vivo studies targeting CAFs in cancer models have demonstrated suppression of tumor growth34. Therefore, targeting these cells in patients is expected to slow down the growth of cancer cells and improve patient’s life expectancy35.
Co-culture of fibroblast and melanoma cells affects the pattern of collagenases expression: As demonstrated in Fig. 2, the study showed a (177 fold) significant increase (p<0.05) of the expression of MMP1 in co-culture setting which was significantly reduced (p<0.05) in A375 cancer cells monoculture. The relative expression of MMP8 gene was significantly reduced (p<0.05) in A375 cancer cells co-culture setting, while it was significantly increased (p<0.05) in A375 melanoma cells monocultures.
The MMPs are reportedly known as important contributors in the progression of the tumor13 either by directly breaking down the components of the extra cellular matrix easing the escape of cancer cells or by activating several signaling pathways through the release of different cytokines, growth factors and adhesion molecules within the extracellular environment. These reactions participate in enhancing cancer cell migration, angiogenesis, invasion and metastasis10. However, these changes are not applicable to all MMPs members. Flach et al.1, have shown an overexpression of MMP1 and MMP13 in the vertical growth phase of highly invasive malignant melanoma cells in comparison with the local less aggressive type of melanoma. The source of these MMPs was first shown to be by the cancer cells however Flach et al.1 elucidated that these enzymes are more significantly produced by the stromal fibroblasts in the tumor stroma. The MMP1 released by these stromal cells was shown to promote the metastatic behavior of cancer cells and induce profound inflammatory response causing more aggressive features by the cancer cells2. The MMP8 had shown some controversial influences on the progression of cancer cells despite the consensus about the active role played by MMPs in invasion and metastasis. Vihinen et al.36 elucidated the tumor promoting effect by MMP8 in patients with malignant melanoma tumor. However, the later evidence did elucidate that this MMP exhibits some anti-tumor properties by limiting the metastatic behavior of breast cancer cells and improving the prognosis of patients with squamous cell carsinoma10,37. Also, it was noticed that any interference with the enzymatic activity of MMP8 might worsen the prognosis of cancer, whereby correlation was elucidated between mutations of MMP8 gene with reduction of enzymatic activity and an increased proliferation and invasion of melanoma cancer cells38. These evidences are in line with the study results that showed an increase in the expression of MMP1 and a reduction in MMP8 in A375 cancer cells co-culture setting as mentioned earlier.
YB-1 protein is highly expressed in melanoma cells without binding to MMP AP-1 gene sequence: In this study, there was an increased expression of the YB-1 protein in A375 melanoma cells during the exponential growth phase as shown in Fig. 2. Although the co-culture cells had a lower expression of the Yb-1 protein, the gene expression was not significantly different. This finding is in agreement with studies that confirmed the association between the level of YB-1 protein and the aggressiveness of many types of tumors such as in breast cancer, synovial sarcoma39, non-small-cell lung carcinoma40 and prostate cancer41. Although some studies claimed the binding YB-1 with AP-1 sequence of MMPs gene promoter is necessary to enhance their expression as stated by Sarkar et al.42. However, in the studies conducted by Norusis20 and Samuel et al.22, YB-1 protein demonstrated an MMP gene insulating effect by binding to the AP-1 sequences. In this study, though YB-1 was highly expressed by the A375 melanoma cells, however, later ChIP analysis revealed no in vivo interaction with AP-1 sequence of MMP gene as shown in Fig. 4. The AP-1 sequence was validated in the extracted DNA from A375 cells. Also the positive control histone antibody did bind with the DNA segments containing the AP-1 sequence. This can be explained by the evidence of YB-1 protein involvement with the acceleration of well-known cancer activated pathways such as MAPK pathway, PI3K/Akt/mTOR pathway and p53 pathways that might be the underlying mechanism to enhance the expression of these enzymes24-26. Additionally, the Yb-1 protein may also activate the tumor stromal cells by releasing different growth factors43. In the same context, several in vivo and in-vitro studies were established with transient and permanent knock down of YB-1 protein expression were associated with anti-invasive, anti-metastatic, anti-proliferative and other anticancer influences within the cancer hallmarks of progression44,45. Therefore, YB-1 remains as an essential biomarker of cancer progression and as a promising target for treatment.
CONCLUSION
The study elucidated that stromal cells (fibroblasts), MMP1 enzyme and YB-1 oncogenic factor are promising targets in the development of cancer treatment that might improve the life expectancy of cancer patients. However, in this study, the mechanism of YB-1 regulation of MMPs expression was not revealed. Therefore intervention studies by knocking down the expression of YB-1 protein with further investigation on different signaling pathways, growth factors and chemokines are necessary to uncover its relation with the expression of these MMPs.
SIGNIFICANCE STATEMENTS
This study elucidates the effect of stromal cells in the tumor microenvironment that can be considered as an important target in the treatment of cancer patients. Also, it confirms that the effect of YB-1 protein on the expression of collagenases MMPs does not involve its binding to the AP-1 gene promoter sequence.
ACKNOWLEDGMENT
This study was supported by the research initiative grant scheme from International Islamic University Malaysia (RIGS 15-089-0089).
REFERENCES
- Flach, E.H., V.W. Rebecca, M. Herlyn, K.S.M. Smalley and A.R.A. Anderson, 2011. Fibroblasts contribute to melanoma tumor growth and drug resistance. Mol. Pharm., 8: 2039-2049.
CrossRefDirect Link - Cirri, P. and P. Chiarugi, 2012. Cancer-associated-fibroblasts and tumour cells: A diabolic liaison driving cancer progression. Cancer Metastasis Rev., 31: 195-208.
CrossRefDirect Link - McAnulty, R.J., 2007. Fibroblasts and myofibroblasts: Their source, function and role in disease. Int. J. Biochem. Cell Biol., 39: 666-671.
CrossRefDirect Link - Shiga, K., M. Hara, T. Nagasaki, T. Sato, H. Takahashi and H. Takeyama, 2015. Cancer-associated fibroblasts: Their characteristics and their roles in tumor growth. Cancers, 7: 2443-2458.
CrossRefDirect Link - Bremnes, R.M., T. Donnem, S. Al-Saad, K. Al-Shibli and S. Andersen et al., 2011. The role of tumor stroma in cancer progression and prognosis: Emphasis on carcinoma-associated fibroblasts and non-small cell lung cancer. J. Thoracic Oncol., 6: 209-217.
CrossRefDirect Link - Fabris, V.T., A. Sahores, S.I. Vanzulli, L. Colombo, A.A. Molinolo, C. Lanari and C.A. Lamb, 2010. Inoculated mammary carcinoma-associated fibroblasts: Contribution to hormone independent tumor growth. BMC Cancer,Vol. 10.
CrossRefDirect Link - Kim, S.H., C. Choe, Y.S. Shin, M.J. Jeon and S.J. Choi et al., 2013. Human lung cancer-associated fibroblasts enhance motility of non-small cell lung cancer cells in co-culture. Anticancer Res., 33: 2001-2009.
PubMedDirect Link - Hassona, Y., N. Cirillo, K. Heesom, E.K. Parkinson and S.S. Prime, 2014. Senescent cancer-associated fibroblasts secrete active MMP-2 that promotes keratinocyte dis-cohesion and invasion. Br. J. Cancer, 111: 1230-1237.
CrossRefDirect Link - Gialeli, C., A.D. Theocharis and N.K. Karamanos, 2011. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS. J., 278: 16-27.
CrossRefPubMedDirect Link - Nissinen, L. and V.M. Kahari, 2014. Matrix metalloproteinases in inflammation. Biochim. Biophys. Acta Gen. Subj., 1840: 2571-2580.
CrossRefDirect Link - Moro, N., C. Mauch and P. Zigrino, 2014. Metalloproteinases in melanoma. Eur. J. Cell Biol., 93: 23-29.
CrossRefDirect Link - Hua, H., M. Li, T. Luo, Y. Yin and Y. Jiang, 2011. Matrix metalloproteinases in tumorigenesis: An evolving paradigm. Cell. Mol. Life Sci., 68: 3853-3868.
CrossRefDirect Link - Chaudhary, A.K., S. Chaudhary, K. Ghosh and A. Nadkarni, 2016. Pleiotropic roles of metalloproteinases in hematological malignancies: An update. Asian Pac. J. Cancer Prev., 17: 3043-3051.
PubMedDirect Link - Schropfer, A., U. Kammerer, M. Kapp, J. Dietl, S. Feix and J. Anacker, 2010. Expression pattern of matrix metalloproteinases in human gynecological cancer cell lines. BMC Cancer, Vol. 10.
CrossRefDirect Link - Thirkettle, S., J. Decock, H. Arnold, C.J. Pennington, D.M. Jaworski and D.R. Edwards, 2013. Matrix metalloproteinase 8 (collagenase 2) induces the expression of interleukins 6 and 8 in breast cancer cells. J. Biol. Chem., 288: 16282-16294.
CrossRefDirect Link - Westermarck, J. and V.M. Kahari, 1999. Regulation of matrix metalloproteinase expression in tumor invasion. FASEB J., 13: 781-792.
PubMedDirect Link - Samuel, S., K.K. Beifuss and L.R. Bernstein, 2007. YB-1 binds to the MMP-13 promoter sequence and represses MMP-13 transactivation via the AP-1 site. Biochim. Biophys. Acta-Gene Struct. Expression, 1769: 525-531.
CrossRefDirect Link - Karin, M., Z.G. Liu and E. Zandi, 1997. AP-1 function and regulation. Curr. Opin. Cell Biol., 9: 240-246.
CrossRefPubMedDirect Link - Samuel, S., J.C. Twizere and L.R. Bernstein, 2005. YB-1 represses AP1-dependent gene transactivation and interacts with an AP-1 DNA sequence. Biochem. J., 388: 921-928.
CrossRefDirect Link - Eliseeva, I.A., E.R. Kim, S.G. Guryanov, L.P. Ovchinnikov and D.N. Lyabin, 2011. Y-box-binding protein 1 (YB-1) and its functions. Biochemistry (Moscow), 76: 1402-1433.
CrossRefDirect Link - Kaufmann, T., A. Strasser and P.J. Jost, 2012. Fas death receptor signalling: Roles of Bid and XIAP. Cell Death Differ., 19: 42-50.
CrossRefDirect Link - Sinnberg, T., B. Sauer, P. Holm, B. Spangler, S. Kuphal, A. Bosserhoff and B. Schittek, 2012. MAPK and PI3K/AKT mediated YB‐1 activation promotes melanoma cell proliferation which is counteracted by an autoregulatory loop. Exp. Dermatol., 21: 265-270.
CrossRefDirect Link - Lasham, A., C.G. Print, A.G. Woolley, S.E. Dunn and A.W. Braithwaite, 2013. YB-1: Oncoprotein, prognostic marker and therapeutic target? Biochem. J., 449: 11-23.
CrossRefDirect Link - Yamamura, Y., N. Asai, A. Enomoto, T. Kato and S. Mii et al., 2015. Akt-Girdin signaling in cancer-associated fibroblasts contributes to tumor progression. Cancer Res., 75: 813-823.
CrossRefPubMedDirect Link - Lazennec, G. and A. Richmond, 2010. Chemokines and chemokine receptors: New insights into cancer-related inflammation. Trends Mol. Med., 16: 133-144.
CrossRefPubMedDirect Link - Orimo, A., P.B. Gupta, D.C. Sgroi, F. Arenzana-Seisdedos and T. Delaunay et al., 2005. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell, 121: 335-348.
CrossRefDirect Link - Rasanen, K. and A. Vaheri, 2010. Activation of fibroblasts in cancer stroma. Exp. Cell Res., 316: 2713-2722.
CrossRefDirect Link - Hwang, R.F., T. Moore, T. Arumugam, V. Ramachandran and K.D. Amos et al., 2008. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res., 68: 918-926.
CrossRefDirect Link - Cornil, I., D. Theodorescu, S. Man, M. Herlyn, J. Jambrosic and R. Kerbel, 1991. Fibroblast cell interactions with human melanoma cells affect tumor cell growth as a function of tumor progression. Proc. Natl. Acad. Sci. USA., 88: 6028-6032.
Direct Link - Shao, H., L. Cai, J.M. Grichnik, A.S. Livingstone, O.C. Velazquez and Z.L. Liu, 2011. Activation of Notch1 signaling in stromal fibroblasts inhibits melanoma growth by upregulating WISP-1. Oncogene, 30: 4316-4326.
CrossRefDirect Link - Zhou, L., K. Yang, T. Andl, R.R. Wickett and Y. Zhang, 2015. Perspective of targeting cancer-associated fibroblasts in melanoma. J. Cancer, 6: 717-726.
CrossRefDirect Link - Vihinen, P., I. Koskivuo, K. Syrjänen, T. Tervahartiala, T. Sorsa and S. Pyrhonen, 2008. Serum matrix metalloproteinase-8 is associated with ulceration and vascular invasion of malignant melanoma. Melanoma Res., 18: 268-273.
CrossRefDirect Link - Decock, J., W. Hendrickx, U. Vanleeuw, V. van Belle and S. van Huffel et al., 2008. Plasma MMP1 and MMP8 expression in breast cancer: Protective role of MMP8 against lymph node metastasis. BMC Cancer, Vol. 8, No. 1.
CrossRefDirect Link - Debniak, T., A. Jakubowska, P. Serrano-Fernandez, G. Kurzawski and C. Cybulski et al., 2011. Association of MMP8 gene variation with an increased risk of malignant melanoma. Melanoma Res., 21: 464-468.
CrossRefDirect Link - Oda, Y., Y. Ohishi, T. Saito, E. Hinoshita and T. Uchiumi et al., 2003. Nuclear expression of Y‐box‐binding protein‐1 correlates with P‐glycoprotein and topoisomerase II alpha expression and with poor prognosis in synovial sarcoma. J. Pathol., 199: 251-258.
CrossRefDirect Link - Gessner, C., C. Woischwill, A. Schumacher, U. Liebers and H. Kuhn et al., 2004. Nuclear YB‐1 expression as a negative prognostic marker in nonsmall cell lung cancer. Eur. Respir. J., 23: 14-19.
CrossRefDirect Link - Gimenez‐Bonafe, P., M.N. Fedoruk, T.G. Whitmore, M. Akbari and J.L. Ralph et al., 2004. YB-1 is upregulated during prostate cancer tumor progression and increases P-glycoprotein activity. Prostate, 59: 337-349.
CrossRefDirect Link - Sarkar, R., J. Lee, R. Mahimkar and D. Lovett, 2004. P53, YB-1 and the AP-1 family of transcription factors mediate MMP-2 transcription induced by skeletal muscle ischemia. J. Am. Coll. Surgeons, 99: 103-103.
CrossRefDirect Link - Gopal, S.K., D.W. Greening, R.A. Mathias, H. Ji and A. Rai et al., 2015. YBX1/YB-1 induces partial EMT and tumourigenicity through secretion of angiogenic factors into the extracellular microenvironment. Oncotarget, 6: 13718-13730.
Direct Link - Wang, H., R. Sun, M. Gu, S. Li and B. Zhang et al., 2015. shRNA-mediated silencing of Y-box binding protein-1 (YB-1) suppresses growth of neuroblastoma cell SH-SY5Y in vitro and in vivo. PloS One, Vol. 10, No. 5.
CrossRefDirect Link - Dolfini, D. and R. Mantovani, 2013. Targeting the Y/CCAAT box in cancer: YB-1 (YBX1) or NF-Y? Cell Death Differ., 20: 676-685.
CrossRefPubMedDirect Link