
M. Harish Kumar
Department of Physiology II, Miyazaki Medical College, Miyazaki, Japan
K.S. Radha
Department of Physiology II, Miyazaki Medical College, Miyazaki, Japan
S.C. Gajaria
Laboratory of Applied Phycology, Department of Biosciences, Sardar Patel University, Vallabh Vidya Nagar 388 120, Gujarat, India
International Journal of Cancer Research
Year: 2005 | Volume: 1 | Issue: 1 | Page No.: 41-46







ABSTRACT
We investigated the effects of algae (Chlorella vulgaris and Enteromorpha sp.) incorporated diets in the prevention of diethyl nitrosamine (DEN: C4H10-N2O) induced hepatocarcinoma in rats. Toxicity was induced by i.p. administration of DEN in multiple doses @ 150 mg kg-1 BW. Rats were divided into 8 experimental Groups and fed either with control diet (synthetic diet without algae) or algae incorporated synthetic diets for a period of 8 weeks. DEN treated rats fed with synthetic diet showed a significant (p<0.05%) reduction in bodyweight, organ weight and relative organ/bodyweight, while Groups fed with algal diets showed gradual recovery from the morphological phenomenon. Likewise, algal diets helped in the process of improvement from liver damage as judged by studying the fluctuation of liver enzymes namely cytochrome P-450, SGPT, G-6-phosphatase and amino-n-demethylase. Serum enzymes, cathepsin B, cathepsin D, ß-D-galactosidases and glucosidases showed significantly (p<0.05%) lower activities in algal diet fed Groups than the synthetic diet fed Group. These results imply or suggest that a combination of Chlorella and Enteromorpha in diets is efficient in limiting the action of DEN during the initiation phase of hepatocarcinogenesis in rats and may help in prevention of liver damage.
PDF Abstract XML References
How to cite this article
M. Harish Kumar, K.S. Radha and S.C. Gajaria, 2005. Algal Diets Reverse Diethyl Nitrosamine (DEN) Induced Hepatocarcinoma in Rats. International Journal of Cancer Research, 1: 41-46.
DOI: 10.3923/ijcr.2005.41.46
URL: https://scialert.net/abstract/?doi=ijcr.2005.41.46
DOI: 10.3923/ijcr.2005.41.46
URL: https://scialert.net/abstract/?doi=ijcr.2005.41.46
INTRODUCTION
Diethyl nitrosamine (DEN; C4H10-N2O; MW 102.14), used as an additive in gasoline and lubricant oil and as a stabilizer compound in plastic industry is a known carcinogen. Nitrosocompound lead tumurogenesis, as described by Boremblunis theory, is a two stage process involving initiation and promotion stages like DNA damage, leakage and repair of nuclear DNA, finally leading to other types of carcinogenesis in vivo by other metabolic pathways[1]. The nutritional and therapeutic capacity of algae and vegetables as protective dietary factors against many types of cancers is well understood[2-4]. Algal metabolites like pigents, peptides and carbohydrates are principle agents to prevent nitrosocompound carcinogenesis[5] in animal studies. In Asian countries algae have been traditionally consumed along with daily food. Chlorella vulgaris and Enteromorpha sp. belong to family Chlorophyceae. The principal components of Chlorella like chlorophyll, ß-carotene, Chlorella Growth Factor (CGF), many vitamins, minerals as well as dietary fibers are reported to have health benefits. Recent investigations show Chlorella to be a potent detoxification agent[6], tumor suppressing agent[7], stress-induced apoptosis reducing agent in mice[8], gene expression agent in human growth hormone production[9], a beneficial antimetastatic immunopotention organism[10] and a good feed additive for marine herbivorous fish[11]. Chlorella vulgaris also helps in prevention of infection from Listeria monocytogens[12], hepatotoxicity and lipid peroxidation[13] and cures fibromyalgia syndrome[14]. Enteromorpha is a macroscopic fresh water alga, exhibiting suppressive effect on initiation and promotion phase of chemically induced mouse skin tumorogenesis[15]. Bioactivation and detoxification system plays an important role in carcinogenesis and are regulated by phase 1 and phase 2 enzymes like cyto-P450 and GST, respectively.
The present research was designed to study the effect of Chlorella vulgaris and Enteromorpha in combination and individually at different concentrations in treating DEN induced hepatotoxicity in rats. To our knowledge, the effect of combined algal diets on the actions of these enzymes to study the hepatotoxicities treatment is a less focused area. Furthermore, this study may help to understand the extent of interaction between algal diets in xenobiotic induced liver damage and its prevention in animal model.
MATERIALS AND METHODS
Chemicals: DEN and enzyme substrates of P-nitrophenol derivatives were obtained from Siga Fine chemical Co., St. Louis, MO-USA. All other chemicals and reagents used were of analytical grade and manufactured by SDH chemicals, Mumbai, India.
Animals and Housing: Wister strain rats were obtained from local scientific suppliers (Ahmedabad, India) and housed in polypropylene cages with sterile, inert husk materials as bedding. Commercial feed (Hindustan Lever feed Ltd, Mumbai) and water free of contaminants were provided ad libitum. The experimental animals were maintained in a controlled environment with 25oC and 60±5% relative humidity with 12 h light/dark cycle and were subjected to one week acclimatization period.
Organism (Algae): Enteromorpha sp. collected from fresh water river, was transported immediately to the laboratory and cleared of sand stone particles. Algae was washed in tap water, rinsed in deionised water, shade dried and powdered using electric ball mill. Chlorella vulgaris was grown in the laboratory in bioreactors using Arnon and Arnold medium. Fresh cells were harvested by centrifugation, rinsed in deionised water, dried and stored at room temperature in glass containers.
Statistical analysis: Analysis of data are presented as mean±SD for 10 rats in each Group. Significant differences between mean values were determined by one-way analysis of variance (ANOVA) and separated by using Duncan’s Multiple comparison test. Comparison of the observed values for statistically significant differences were set at p<0.05%. The analysis was carried out with the aid of the Prism Graphpad 3.03 version statistical package.
Experimental design: Prior to start of the experiment, all animals were caged and acclimatized for one week period and fed with synthetic diet. In total, 80 rats with body weights ranging from 185±2.1 to 202±1.6 g were used in the experiment and were divided into 8 Groups. While 7 Groups were administrated with DEN (in saline) intraperitoneally, daily, at 150 mg kg-1 body weight for 7 days, Group 8 alone received saline injection instead of DEN. Synthetic diet feeding was continued up to the second week i.e. till the completion of DEN course. Group 1 fed with synthetic diet alone and Group 8 animals that did not receive DEN served as control Groups. Experimental diets were prepared with different doses of powdered algae incorporated with the synthetic diet. Group 2 and 3 received diets containing 25 and 50% Chlorella dry powder mixed with synthetic diet proportionally. Group 4 received mixed diets containing 25% Chlorella, 25% Enteromorpha and 50% synthetic diet. Enteromorpha dry powder mixed at the rate of 25 and 50% along with synthetic diet proportionally was fed to Groups 5 and 6, respectively. Group 7 received diet of 40% Chlorella, 40% Enteromorpha and 20% synthetic diet. This feeding schedule continued up to 8 weeks. Feeding trail was monitored according to the standards of animal experimentation and feed water consumption was recorded on a daily basis. Daily observation of body weight and mortality rate was recorded as mean values on a weekly basis.
After the experimental period, blood from the animals was collected by cardiac puncture and centrifuged at 5000 rpm for 15 min. The serum was stored at -70oC for further analysis. All surviving rats were sacrificed by ex sanguification from the abdominal aorta under light anesthetic condition at the end of 8 weeks. Liver was dissected and 5 g was homogenized in phosphate buffer saline (PBS 0.05M, pH 7.2) using polytron homogenizer. Microsomes were prepared from liver by differential centrifugation method[15] and resuspended in Tris-HCl buffer (0.5 M, pH 7.2) containing 0.5 M sucrose and 10% glycerol.
Cyctochrome P450[16], Serum glutamate pyruvate transaminase (SGPT)[17], glucose-6-phosphatase[18], aminopyrine-n-demethylase[19] and total protein[20] were determined according to published methods. Activities of serum glycosidases namely ß-D-galactosidases, ß-D-glucosidases[21], cathepsin B[22] and cathepsin D[23] were estimated according to standard methods.
RESULTS
Quantitative data of body weight changes during the experimental period is given in Table 1. Treatment with DEN (Group 1) showed a significant reduction in body weight at the end of 8 weeks. Group 2 also showed a significant reduction in body weight (p<0.05%) corresponding to the initial week, while other Groups showed a marginal improvement/increase in body weight. It is interesting to note that Group 7 showed a significant (p<0.05%) increase in mean body weight from 190±1.7 g to 203±1.7 g at the termination of the experiment. Increase in the body weight was also noticed in the Group 8 animals but did not show any significant fluctuations in between weeks (Table 1). Basic data on mortality, body weight, liver weight and relative body vs liver weight ratio was recorded. As indicated in Table 2, mortality rate was highest in Group 1, showing the severity of toxicity in the animals. Rate of mortality reduced gradually as incorporation of algae in the diet increased.
| Table 1: | Body weight changes (g) in 8 weeks |
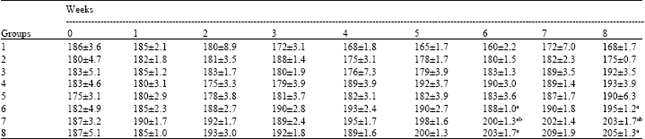 | |
| Values represent mean ±SD of three independent experiments, each consisting of data pooled from 10 rats. Superscript a denotes a significance (p<0.05%) than Group 1 and b denotes significant than Group 1 and Group 6 (Duncan’s Multiple Range Test) | |
| Table 2: | Mortality rate, body weight, liver weight and relative liver weight after eight weeks of experimental periods | ||||||||||||
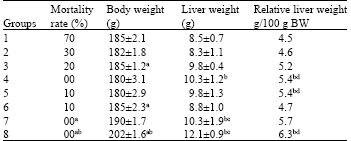 | |||||||||||||
| Values represent mean±SD of three independent experiments, each involving 10 rats | |||||||||||||
| |||||||||||||
| Table 3: | Activities of serum cathepsins and glycosidases | |||||||||
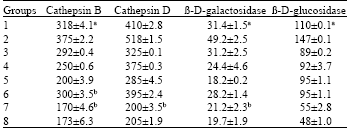 | ||||||||||
| Serum Cathepsin and Glycosidases activity in rats treated with different algal diets and control diets (without algae) in different Groups. Enzyme activity was measured according to standard methods. | ||||||||||
| Values represent mean±SD of three independent experiments, each consisting of tissues pooled from 10 rats. Cathepsin B unit: n mol P-nitroanilide liberated/h/mL serum. Cathepsin D unit: n mol of tyrosine formed/h/mL serum. ß-D-galactosidase unit: n mol p-nitrophenol liberated/h/mL serum. ß-D-glucosidases unit: n mol p-nitrophenol liberated/h/mL serum. | ||||||||||
| ||||||||||
Pattern of liver weight changes reveals the lowest relative liver weight of 4.5/100 g BW (p<0.05%) in Group 1. On the other hand, all the other Groups, which were given algal powder, did show improvement in relative liver index. Chlorella and Enteromorpha at 40% w/w basis (Group 7) showed the highest value of 5.7/100 g BW (Table 2).
| Table 4: | Activities of liver enzymes after 8 weeks after treating with DEN and algal diets | |||||||||
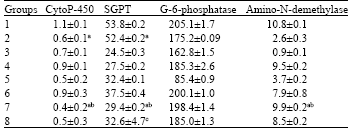 | ||||||||||
| Values represent mean±SD of three independent experiments, each consisting of tissues pooled from 10 rats. Cytochrome-p450 unit: n mol mg-1 protein SGPT unit: Units/mL/mg protein; G6 phosphate unit: n mol P liberated/min/mg protein Amino-N-demethylase unit: n mol/min/mg protein | ||||||||||
| ||||||||||
This result suggests that algae at highest concentration do have beneficiary effect on gross health condition of rats.
Serum enzymes: Table 3 summarizes the activities of the marker enzymes cathepsin B, cathepsin D, ß-D-galactosidase and ß-D-glucosidase in serum. A significant (p<0.05%) enhancement of all four enzymes were observed in DEN treated animals (Group 1). Notably, Group 7 fed with the highest concentration of algae in combination showed highly decreased activity that is statistically significant (p<0.05%) and almost similar to Group 8. Interestingly, ß-D-galactosidase alone was significantly higher (p<0.05%) in 25% Enteromorpha fed Group than the control values. ß-D-glucosidases in the rats given DEN (Group1) was significantly higher as compared to animals treated with algal diets and non treated animals (Group 8).
Liver enzymes: In order to explore the index of DEN toxicity and cancer prevention by algal diets, activities of different liver enzymes namely Cyto-P450, SGPT, G-6-phosphatase and amino-N-demethylase were measured (Table 4). SGPT activity in algal diet fed Groups tended to
be lower than DEN treated animals (Group1). The elevated activity of Cyto-P450 in Group1 is also notable observation in the present study. Algal diets significantly decreased the G-6-phosphatase activities than the synthetic diet. The elevated activity of Amino-N-demethylase activity induced by DEN was reduced significantly (p<0.05%) in Groups 5 and 6, indicating that elevated enzyme activity induced by DEN can be suppressed to normal values by consumption of algal diets.
DISCUSSION
It is known for decades that consumption of algae and seaweed reduces the risk of many types of cancer in human beings. Glycosidases are lysosomal enzymes responsible for degradation of carbohydrate moieties of glycoprotein, lipid, glycosaminoglycans and are known marker enzymes in cancer study[24]. Likewise an elevated activity of cathepsin-a protease is also an index of malignancy[25,26].
Wattenburg[27] showed that the synergistic actions of blocking agents occurring in diets are important in cancer prevention. Morita et al.[6] found that dietary fiber in algae is a good agent for reduction of PCB and dioxin toxicity in rats. Dietary fibers are digested slowly in colon to form short chain fatty acids like acetate, propionate and butyrate by fermentation. It has been proposed that butyrate inhibits cell proliferation, promotes cell differentiation and leads to apoptosis, contributing to the healthy condition of colon mucosa[28]. This study, combination of chlorella with Enteromorpha at 40% concentration level showed efficiency in preventing DEN induced carcinogenesis. Chlorophyllin-a chlorophyll derivative forms a complex with heterocyclic amines[29,30]. Chlorella and Enteromorpha have rich amount of chlorophyll and fiber respectively, which form complex with DEN thereby reducing the toxicity. Another line of studies by Lipman et al.[31] showed that retinoid in vegetables and fruits decrease carcinogenicity by inhibiting the cytochrome P-450 directed metabolic product. In the present study, high cytochrome P-450 (cyto-P450) in Group 1 reduced gradually in algae incorporated diets and showed lowest activity of 0.5±0.08 in Group 7 (Table 4). Nutritional and non-nutritional factors have direct relationship on the expression of the highly inducible cyto-P450 activity in animals and humans. Cytochrome P-450 is known to catalyze the N-oxidation of 4-aminophenyle, 2-napthalamine and several aromatic amines, which have proved to increase the risks of cancer. It is a key enzyme in the pathway of heterocyclic amine metabolic activation leading to final DNA binding forms[1]. Levels of cytochrome P-450 in rat liver is modulated selectively by retinoids that scavenge the reactive oxygen [RO] in animal body. In agreeing with results of Lipmann et al.[31] this study shows that Chlorella and Enteromorpha with good amounts of total retinoids and chlorophyll are better for hepatotoxic prevention since they have higher amount of active compounds. The exact mechanism could not be drawn in this experiment, but, indirect interaction of algal metabolites like chlorophyll and retinoids and marker enzymes in animal body is likely the mechanism of actions in prevention of further metabolism of DEN in animal body. The present study implies that algal ingredients mainly ß-carotene and chlorophyllin block the carcinogen metabolic activation and enhance the carcinogen detoxification in animals. Bhat et al.[32] studied that phycocyanin prevents hepatotoxicity, by causing a significant decrease of G-6-phosphatase and amino-n-demethylase activities in carbon tetrachloride induced hepatotoxicity in liver. Similarly, we observed the elevated activity of these two enzymes in DEN treated animals (Table 4), be significantly reduced with the aid of the algal diets.
Infiltration and migration of tumor cells to surrounding tissues is facilitated and enhanced by destruction and modification of structural proteins by release of proteolytic enzymes that solubilise the extra cellular matrix[33]. Serum proteases and glycosidases have been proposed as potential indicator molecules of inflammation condition in human beings. It may be possible that differential fluctuation of protease glycosidase activity in serum could modify the integrity of cell surface as stated by Bosmann[34]. Elevated levels of cathepsins B and D is a significant observation in human breast cancer cells[13] and gastric cancer[28]. The mechanism of increase in lysosomal enzymes activity during the course of carcinogenesis is not completely clear but latent amount of activity can be liberated during injury of cells in body inflammation conditions. In our studies, wherein we examined the influence of algal diets on the activities of cathepsins B and D, we found a significant increase (p<0.05%) in both the cathepsins in DEN treated Group than the untreated or algal diets treated Groups (Table 2). Toda and Yokogoshi<’s[35] suggestion that soya protein isolate caused an improvement in muscle condition through reduction of proteosome activities may indicate that in the current study, protein moiety present in the algae interacted with the cathepsins B and D to help in the detoxification of DEN to ameliorate the muscle atrophy, finally leading to improvement of body weight. It is probable that administration of Chlorella and enteromorpha in diets may prevent the migration of T-cell and ß cells as observed by Cheng et al.[36] either by decreasing the protease activity or checking the onslaught of free radical attack and decreasing the augmented levels of cathepsins B and D and glycosidases in carcinogen induced animals. However specific cell line in vitro experiments are required to establish this hypothesis. Although results are promising, it is difficult to judge the pattern of action of algal active ingredients in vivo at present. Therefore, it is necessary to investigate the mechanisms of actions of other enzyme pathways and at the molecular level.
In conclusion, the present study reveals that active compounds present in Chlorella and Enteromorpha suppress cancer promotion is an interesting and promising subject for future study. Daily intake of algal diets such as Chlorella and Enteromorpha may be useful against hepatotoxicity.
ACKNOWLEDGMENT
The authors thank the Head of the Department for providing with facilities to carry out this study.
REFERENCES
- Okuzumi, J., T. Takahashi, T. Yamane, Y. Kitao and M. Inagake et al., 1993. Inhibitory effects of fucoxanthin, a natural carotenoid, on N-ethyl-N-nitro-N-nitrosoguanidine-induced mouse duodenal carcinogenesis. Cancer Lett., 68: 159-168.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link