
Baby Joseph
Interdisciplinary Research Centre, Department of Biotechnology, Malankara Catholic College, Mariagiri-629 153, Kanyakumari District, Tamil Nadu State, India
P. Sankarganesh
Interdisciplinary Research Centre, Department of Biotechnology, Malankara Catholic College, Mariagiri-629 153, Kanyakumari District, Tamil Nadu State, India
Biby T. Edwin
Interdisciplinary Research Centre, Department of Biotechnology, Malankara Catholic College, Mariagiri-629 153, Kanyakumari District, Tamil Nadu State, India
S. Justin Raj
Interdisciplinary Research Centre, Department of Biotechnology, Malankara Catholic College, Mariagiri-629 153, Kanyakumari District, Tamil Nadu State, India
International Journal of Biological Chemistry
Year: 2012 | Volume: 6 | Issue: 2 | Page No.: 42-52







ABSTRACT
Endophytic Streptomycetes are the most exploited group in the medicinal and agro based industries. In this review, we correlated recently identified its bioactive compounds with plant chemical structures. Booming the global warm is the important problem to increase the alternative research initiatives for green chemicals. During long genetic co-evaluation with plants Streptomycetes have created novel and interesting chemistry. Their ability to produce plant related chemicals has increased its importance. It also reduces the need of harvesting plants for drug. Specific approaches from ethanobotanical valued plants with medicinal applications could be the key to isolating bioactive compound producing strains.
PDF Abstract XML References Citation
Received: January 03, 2012;
Accepted: March 06, 2012;
Published: May 29, 2012
How to cite this article
Baby Joseph, P. Sankarganesh, Biby T. Edwin and S. Justin Raj, 2012. Endophytic Streptomycetes from Plants with Novel Green Chemistry: Review. International Journal of Biological Chemistry, 6: 42-52.
URL: https://scialert.net/abstract/?doi=ijbc.2012.42.52
URL: https://scialert.net/abstract/?doi=ijbc.2012.42.52
INTRODUCTION
The rise of new and more drug resistant infectious diseases as a result of climatic change and deforestation remain one of the biggest killers in developing countries (Stella et al., 2010). Additionally, the resistance pattern of pathogens to synthetic drugs often creates a need for stronger and stronger drugs or an increased dosage that could result in severe deterioration of the patient’s health and an unexpected growth of drug resistance organisms like NDM-1 bacteria (Kumarasamy et al., 2010). So, countries spend billions of dollars to identify new broad spectrum antibiotics (Breman et al., 2007). Antibiotics have undoubtedly made major contributions to the improvement of health and welfare in both human and animals. In recent year there has been an alarming rise in the prevalence of resistance to some agents among certain groups of pathogens (Carl and Pascal, 2006). Concern is growing that therapeutic options will become increasingly limited if resistance rates continue to rise as it has in the past few decades. There is widespread agreement that immediate action is required to reverse or at least slow this process. Vector transmitting pathogens are a high level threat to humans and animal. The climatical changes will tremendously influence life cycles of arthropod vectors for the maximum productivity which resulting in changes in both vector and pathogen distribution and changes in the ability of arthropods to transmit pathogens (Tabachnick, 2010). So, there is an urgent need to study the most commonly used drugs for their potency in reducing parasite growths and symptoms of the disease (Talisuna et al., 2004).
Traditional approaches from natural sources: The World Health Organization estimates that over 65% of the world’s population relies on traditional medicine for their primary health needs (Wang et al., 2007; Fabricant and Farnsworth, 2001). The planet has various plant biodiversity. Natural products especially plants has used to treat various infectious diseases for thousands of years. Almost all plant varieties have at least some minimal source of medicinal properties. History records the use of natural products (Bhutani and Gohil, 2010) to cure many diseases i.e., TB, Leprosy, Gonorrhoea, various skin diseases and some internal disorders. In the battle field, wounded soldiers were treated with extracts of various plant leaves. Indian Ayurvedic systems, Susruta and Charaka dating back to 1000 BC and still now widely practiced, have used plant extracts as their main source of treatment (Kapoor, 1990). After invention of Penicillin by A. Flemming, the biomedical world has realized importance of microbial sourcing. The discovery of a plethora of microbes for applications has span broad spectrum active medicines (anti-TB, malaria, cancer, etc.).
Plants based phenols and polyphenols, flavonoids, isoflavones, terpenes and glucosinolates are responsible for bitterness (Drewnowski and Gomez-Carneros, 2000) and these “bitter,” chemicals generate signals which affect a cascade of regulatory events resulting in chemical differentiation in Streptomyces genome by activating “cryptic pathways” for secondary metabolite biosynthesis (Rigali et al., 2008) and offers new prospects in the fight against emerging diseases. The chemical and biological diversity of nature is immeasurable and provides an extraordinary resource for the discovery of new drugs (Luke et al., 2005).
Endophytes are microorganisms that colonize inside plant tissue and are relatively crucial for the medical and agricultural industries (Lodewyckx et al., 2002). They habitat in tissues below the plant’s epidermal cell layers due to the fact that some host plant’s tissues are transiently asymptomatic (Strobel, 2003; Tharek et al., 2011). Some plants were studied for endophytes; fortunately, the opportunity to find novel endophytic microorganisms from variety of plants in the ecosystems was great.
Scientific classification of streptomyces:
| Kingdom | : | Bacteria |
| Phylum | : | Actinobacteria |
| Class | : | Actinobacteria |
| Order | : | Actinomycetales |
| Family | : | Streptomycetaceae |
| Genus | : | Streptomycetes |
Almost all vascular plants harbor several endophytic actinomycetes, which would help to improve the host plant’s growth and promotional activity against diseases (caused by plant pathogens and/or various environmental stress factors) (Sachiko et al., 2006). Antibiotic productivity has been known to closely related to the cultural conditions that exist in a relevant environment (Takao, 2006). Research attempts to discover new bioactive compounds from prokaryotic organisms and in particular, Actinomycetes, seem to have declined over the recent years. Recently, various new genera of actinomycetes were found, resulting in the discovery of new bioactive substances. Bioactive compounds from endophytic Streptomyces have distinct chemical structures that may form the basis for synthesis of new drugs. Pharmaceutical companies and research and development units are trying to develop new bioactive compounds from the organisms by change their molecular characteristics to overcome the drug resistance problem (Bhattacharya and Sen, 2006). Plants have a myriad of chemical and anatomical defenses. The defense intensity level depends on biotic and environmental influences (Mika et al., 2008). Precursors often stimulate production of secondary metabolites by increasing the rate of a limiting precursors and unusual structures. Their formation regulated by nutrients, growth rate, feedback control, enzyme inactivation and enzyme induction (Demain, 1998). Individual organisms that collectively make up the biota have specific properties that make them of direct value in satisfying the consumption and production needs of society. The organisms occupying the endosphere are not accidentally there but most probably have selected for this niche by the plant; due to the beneficial effects they offer their host and their ability to resist the plant’s natural defense mechanisms. The energy lost by a plant in the production of endophytic biomass is in all likelihood adequately compensated for the improvements in plant health derived from the presence of endophytic microorganisms (Backman and Sikora, 2008). The innovative biotechnology field has at its disposal an infinite pool of novel chemistry from microbial diversity (Kekuda et al., 2010). Therefore the study of endophytic microbial diversity is more crucial than ever in understand and combating emerging diseases. Endophytic microbes seem to fit perfectly into this natural ‘warehouse’ of which only a small part, up to now, we have been able to exploit (Agata et al., 2008).
Collection and isolation of endophytic organisms: Accordingly ethanobotanical history, the plant leaves, stems and root portions can be taken for the study. The locations plotted by global positioning system. Small piece of sample can be collected in sterile polythene bag cover and can be stored at 4°C up to the experiment. During the experiment, the plant samples are thoroughly cleaned in tap water to remove the surface soil debris materials. Then the samples can thoroughly treated with 70% ethanol and dried under the aseptic condition. This will removes the epiphytes that present on the samples. Then, using sterile scalpel blade, the xylem and phloem tissues were excised, fragmented and inoculated on to water agar medium and selective isolation mediums. Then after prolong incubation the emerging of the hyphal growth can be screened and purified on the selective nutrient mediums. Their taxonomy can be confirmed by molecular biology techniques such as 16S rRNA gene sequencing. Finally, the pure cultures can be inoculated on the broth culture for the fermentation study. The culture growth parameters can be optimized for the production of the isolate and its bioactive metabolites. From the liquid culture the spores can be transferred into 15-20% glycerol and stored at -70°C as long storage. The bioactive metabolites can be assayed against variety of pathogens and it can be purified using commonly available procedures.
Endophytic microbial products: Endophytic microorganisms are a significant reservoir of genetic diversity and an important source in the discovery of novel bioactive secondary metabolites. These group of strains can produce single or multiple kinds of antibiotics including terpenoids, alkaloids, aromatic compounds and polypeptides (Gao et al., 2010) which are similar to host plant chemicals, thus triggering the expectation that endophytes can serve as an alternative source (Priti et al., 2009). So, plants with beneficial ethanobotanical history are also likely candidate for study. Since the medicinal uses to which the plant may have selected relates more to its population of endophytes than to the plant biochemistry itself. The microorganisms living inside the plant tissue for long duration will evolve due to the genetic variation with the host. This paves the best way to produce some phyto-chemicals by the guest organisms (Zhang et al., 2006). Endophytic organisms are found in all the types of plant tissues such as stems, roots, leaves, fruits, ovules, seeds, tubers, rachis and bark. Probably, hundreds of endophytic species from a single plant are also possible and among them, at least one generally shows host specificity (Tan and Zou, 2001). Thus, if endophytes can produce the same rare bioactive compounds as their host plants, this would not only reduce the need to harvest the slow-growing and possibly rare plants but also keep the world’s ever-diminishing biodiversity and cut human induced climate change.
These endophytes, displaying a broad spectrum of biological activities against plant pathogens continue to increase biocontrol significance. As a general rule, a single endophytic strain will produce multiple bioactive compounds (Strobel et al., 1993). In this respect, future success rests on the ability to find novel endophytic actinomycetes from plants. Previously, many fungal organisms were used for production of similar plant chemicals e.g., Colletotrichum gloeosporioides produces taxol (Justicia gendarussa) (Gangadevi and Muthusamy, 2008); Broad spectrum active Guanacastepene (Singh et al., 2000) antiviral active cytonic acid A and B (Guo et al., 2000), hinnuliquinone (Singh et al., 2004), isobenzofuranones, pestacin and isopestacin (Strobel et al., 2002), paclitaxel (Stierle et al., 1993; Li et al., 1998a, b; Benedikt et al., 2008). These like gifted bioactive endophytes will play the greatest role in bio-prospecting and will conserve the medicinal plants (Strobel et al., 2005) which were needed for novel drug discovery (Bhutani and Gohil, 2010).
Endophytic streptomycete and its novel compounds: Wang et al. (2010) has isolated four new p-aminoacetophenonic acids, Fig. 1-4 from endophytic Streptomyces sp. HK10552 of the mangrove plant Aegiceras corniculatum.
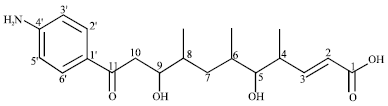 | |
| Fig. 1: | Chemical structure of (2E)-11-(4-aminophenyl)-5,9-dihydroxy-4,6,8-trimethyl-11-oxo-undec-2-enoic acid |
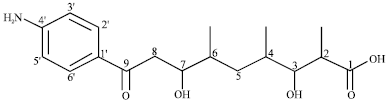 | |
| Fig. 2: | Chemical structure of 9-(4-aminophenyl)-3,7-dihydroxy-2,4,6-trimethyl-9-oxo-nonoic acid |
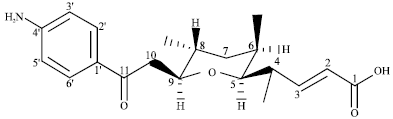 | |
| Fig. 3: | Chemical structure of (2E)-11-(4-aminophenyl)-5, 9-O-cyclo-4,6,8-trimethyl-11-oxo-undec-2-enoic acid |
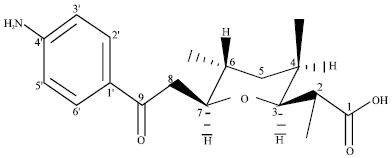 | |
| Fig. 4: | Chemical structure of 9-(4-aminophenyl)-3, 7-O-cyclo-2, 4, 6-trimethyl-9-oxo-nonoic acid |
p-aminoacetophenonic acids: The compound 1 and 2 show structural similarity to the polyene antibiotic, candicidin. Streptomyces griseus, which consists of the aromatic moiety, the macrolide ring and the amino sugar moiety. They have different polyketide chain lengths which may due to different biosynthetic polyketide assembly procedures (Gil and Campelo-Diez, 2003). This Polyene macrolide antibiotic can actively inhibit the fungal pathogens (Zotchev, 2003).
Antimycin A18: Streptomyces albidoflavus isolated from mangrove plants able to produce Antimycin A18, showed broadly spectrum of against pathogenic microorganisms (Yan et al., 2010). Previously Antimycin 1-7 was reported from Streptomyces sp. K01-0031 (Shiomi et al., 2005). A microbial alkaloid, 4-methyl-2-quinazolinamines produced by Streptomyces sp. GS DV232 which is isolated from Traditional Chinese Medicinal plants (TCM). This new alkaloid has shown anti-proliferative bioactivity (Vollmar et al., 2009).
Arylcoumarins: S. aureofaciens CMUAc130 isolated from Zingiber officinale Rose, reported for production of antifungal and antitumor compound 4-Arylcoumarins (5, 7-d i m e t h o x y-4-p- methoxylphenylcoumarin (1), 5, 7-dimethoxy-4-phenylcoumarin (2) (Taechowisan et al., 2005).
These compounds showed possible inhibitory effect on growth of s.c. transplanted Lewis Lung Carcinoma (LLC) BDF-1 mice by intraperitoneal administration. The T/C value of 5, 7 - d i m e t h o x y-4-p-methoxylphenylcoumarin is 80.8 and 50.0% at the doses of 1 and 10 mg kg-1, where, 5, 7-dimethoxy-4-phenylcoumarin is 81.5 and 44.9% at the doses of 1 and 10 mg kg-1. These two antitumor compounds show low toxicity in normal human cell lines and potentially active in malignant cell lines (Taechowisan et al., 2007).
Kakadumycin A: Streptomyces sp. A35-1 (NRRL 30566) isolated from Grevillea pteridifolia, produced broad spectrum antibiotic kakadumycins. This quinoxaline compound related antibiotic is more effective against plant pathogenic fungi including Botrytis sp., Alternaria sp., Helminthosporium sp. and Pythium ultimum. They are potentially active against various drug resistant pathogenic bacteria. It shows activity against malaria parasite Plasmodium falciparum with an IC50 of 4.5 ng mL-1 (Castillo et al., 2003).
Coronomycin: Streptomyces sp. MSU-2110 isolated from Monstera sp., reported to produce coronomycins, a novel peptide antibiotic. It shows activity against human fungal pathogen Cryptococcus neoformans. It acts against the malaria parasite, Plasmodium falciparum, with an IC50 of 9.0 ng mL-1 (Ezra et al., 2004).
Munumbicins A-D: Streptomyces NRRL 30562 isolated from Kennedia nigricans, produces broad spectrum active munumbicins A, B, C and D. These all are highly active against Bacillus anthracis, multidrug resistant Mycobacterium tuberculosis, Methicillin Resistant Staphylococcus aureus (MRSA), Glycopeptides Intermediate and Staphylococcus Aureus (GISA) and plant pathogenic fungal organisms. Munumbicin A shows activity against Vancomycin Resistant Enterococcus faecalis (VRE). Munumbicin B shows activity against Mycobacterium tuberculosis with IC50 of 10 and 46 μg mL-1. Munumbicin C shows activity against malaria parasite Plasmodium falciparum with an IC50 of 6.5 ng mL-1, compared to an IC50 of Chloroquine of 7 ng mL-1. Munumbicin D showed activity against the malaria parasite Plasmodium falciparum, having an IC50 of 4.5±0.07 ng mL-1. Munumbicin B and C showed greatest anticancer activity in ME-180 cancer cell lines (cervical epithelial epidermoid carcinoma). The LC50 value is 0.1-0.14 μg mL-1 (Castillo et al., 2002).
Lansais: An endophyte Streptomyces aureofaciens CMUAc130 produces lansais (5, 7-dimethoxy-4-p-methoxylphenylcoumarin and 5, 7-dimethoxy-4-phenylcoumarin) (Taechowisan et al., 2009). The compound C and D have anti-inflammatory compounds which inhibits the inducible nitric oxide synthase and nitric oxide which induced by pro-inflammatory cytokines during inflammation in LPS-induced RAW 264.7 cells. They mainly inactivate the nuclear factor κB and signal transducer and transcription-1 activator, which are a significant transcription factors for iNOS (Taechowisan et al., 2010).
Cyclopentenone: Streptomyces sp. (GT-20026114) isolated from the mangrove plant Aegiceras corniculatum collected in South China. This organism produces four new cyclopentenone derivatives on seawater-based medium. These rare compounds already reported from Trichoderma sp. which has the potential to convert ciprofloxacin and norfloxacin to those conjugates (Parshikov et al., 2002). They are namely (5R) 3-amino-5-hydroxy-5-vinyl-2- cyclopenten-1-one (1), (5R) 5-hydroxy-3-[(methoxycarbonyl) amino]-5-vinyl-2-cyclopenten-1-one (2), (5R) 5- hydroxy-3-[[2-(4-hydroxyphenyl)ethyl]amino]-5-vinyl-2- cyclopenten-1-one (3) and 3-isobutylpropanamide-2- cyclopenten-1-one (4) (Lin et al., 2005).
Isocutellarin and cichoriin: An root endophyte Streptomyces sp. Tc052 isolated from Alpinia galanga, produces antimicrobial active kaempferol, isocutellarin, umbelliferone and cichoriin which is active against gram positive, gram negative and yeast cells. Its overall MIC value is 64-128 μg mL-1 (Taechowisan et al., 2008).
p-aminoacetophenonic acids: An endophyte Streptomyces griseus isolated from Kandelia candel, produces three new sesquiterpenes (p-aminoacetophenonic acids) which are the antimicrobial compounds (Guan et al., 2005).
Fistupyrone: Spring onion isolated Streptomyces sp. TP-A0569 producing fistupyrone, has been reported for potent antifungal activity (Aremu et al., 2003) against the Alternaria brassicicola which causes black leaf spot in Brassica plants (Igarashi et al., 2000).
Pterocidin: A new cytotoxic compound isolated from the endophytic Streptomyces hygroscopicus TP-A0451. It showed cytotoxicity against some human cancer cell lines with IC50 values of 2.9~7.1 μM (Igarashi et al., 2006). Endophytic Streptomyces laceyi MS53 producing 6-alkylsalicylic acids; salaceyins A and B has more cytotoxicity against a human breast cancer cells (Kim et al., 2006).
CONCLUSION
Natural products are powerful biochemical tools. They serve as ‘pathfinders’ for molecular biology and chemistry and are the investigators of cellular functions (David et al., 2009). The WHO estimates that approximately 80% of the world’s inhabitants rely on traditional medicine for their primary health care (Fabricant and Farnsworth, 2001). It is noteworthy that, of the nearly 300,000 plant species that exist, each individual plant is host to one or more endophytes. Only a few of these plants have been completely studied for their endophytic biology. Accordingly, an opportunity to find new and beneficial endophytic microorganisms among the diversity of plants in different ecosystems is considerable (Ryan et al., 2008). Currently, endophytes looked upon as a prominent source of bioactive natural products. It appears that these biotypical factors can be important in plant selection, since they may govern the novelty and biological activity of the products associated with endophytic microbes (Strobel, 2003). Sometimes extremely unusual and valuable organic substances produced by these endophytes which may assist in solving health problems in humans (Khucharoenphaisan et al., 2012), plants and animals. A specific rational approach for endophytic isolation needed to discover products (Gunatilaka, 2006). The plant choice can be adopted by the geographical nature of the plants and ethanobotanical value of the plants (Gordon et al., 2009). This is one of the most attractive and significant research themes for plant-microbe interactions (Sachiko et al., 2006). Finding new varieties of antibiotics can be one of the greatest events in the history of medicine. This discovery will have a profound effect on human life and our environment (Bhattacharya and Sen, 2006). A search for specific endophytes that may produce antibiotics can have its origin in ethnobotany, which uses the medicinal lore of native peoples.
The over exploitation of wild medicinal flora and fauna for use in modern medicinal systems has led to a significant threat to the endemic medicinal flora and fauna (Arora et al., 2010). There is an enormous possibility for isolation of novel secondary metabolites (Joseph and Priya, 2011) from this poorly investigated group of microbial floras of ethanobotanical valued plants which, is an ample and reliable source of bioactive and novel chemical compound. Majority of naturally occurring compounds in these biological systems are endlessly in a metabolic flux, due to continuous interactions and fresh synthesis. Thus, need to that endophytic Streptomycetes from unexplored or underexploited niches be pursued as source of novel bioactive secondary metabolites (Lam, 2006; Zin et al., 2011; Raja et al., 2011). Although the use of this vast resource has just begun, it is already clear that there is a tremendous opportunity for discovering of organisms, products and the use stimulates continuous hope. It hoped that more efficient and effective application of natural products will improve the drug discovery process. Insights from such research may give alternative methods of producing natural drugs which could be reliable, economical and environmentally safe and will conserve natural biodiversity.
REFERENCES
- Lodewyckx, C., J. Vangronsveld, F. Porteous, E.R.B. Moore, S. Taghavi, M. Mezgeay and D. van der Lelie, 2002. Endophytic bacteria and their potential applications. Crit. Rev. Plant Sci., 21: 583-606.
CrossRefDirect Link - Strobel, G.A., 2003. Endophytes as sources of bioactive products. Microbes Infect., 5: 535-544.
CrossRefPubMedDirect Link - Hasegawa, S., A. Meguro, M. Shimizu, T. Nishimura and H. Kunoh, 2006. Endophytic actinomycetes and their interactions with host plants. Actinomycetology, 20: 72-81.
CrossRefDirect Link - Demain, A.L., 1998. Induction of microbial secondary metabolism. Int. Microbiol., 1: 259-264.
PubMedDirect Link - Backman, P.A. and R.A. Sikora, 2008. Endophytes: An emerging tool for biological control. Biol. Control, 46: 1-3.
CrossRefDirect Link - Kekuda, T.R.P., K.S. Shobha and R. Onkarappa, 2010. Fascinating diversity and potent biological activities of Actinomycete metabolites. J. Pharm. Res., 3: 250-256.
Direct Link - Staniek, A., H.J. Woerdenbag and O. Kayser, 2008. Oliver endophytes exploiting biodiversity for the improvement of natual product based drug discovery. J. Plant Interact., 3: 75-98.
CrossRef - Kumarasamy, K.K., M.A. Toleman, T.R. Walsh, J. Bagaria and F. Butt et al., 2010. Emergence of a new antibiotic resistance mechanism in India, Pakistan and the UK: A molecular, biological and epidemiological study. Lancet Infect. Dis., 10: 597-602.
CrossRefDirect Link - Tabachnick, W.J., 2010. Challenges in predicting climate and environmental effects on vector-borne disease episystems in a changing world. J. Exp. Biol., 213: 946-954.
CrossRef - Talisuna, A.O., P. Bloland and U.D'Alessandro, 2004. History, dynamics and public health importance of malaria parasite resistance. Clin. Microbiol. Rev., 17: 235-254.
CrossRef - Bhutani, K.K. and V.M. Gohil, 2010. Natural products drug discovery research in India: Sataus and appraisal. Indian J. Exp. Biol., 48: 199-207.
Direct Link - Drewnowski, A. and C. Gomez-Carneros, 2000. Bitter taste, phytonutrients and the consumer: A review. Am. J. Clin. Nutr., 72: 1424-1435.
Direct Link - Rigali, S., F. Titgemeyer, S. Barends, S. Mulder, A.W. Thomae, D.A. Hopwood and P. van Gilles, 2008. Feast or famine, the global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Reports, 9: 670-675.
CrossRef - Luke, S.T., A. Eric, M. Kerry, F. Patricia and H.G. William, 2005. Marine natural products as anticancer drugs. Mol. Cancer Ther., 4: 333-342.
Direct Link - Wang, M.W., X. Hao and K. Chen, 2007. Biological screening of natural products and drug innovation in China. Philos. Trans. R. Soc. B: Biol. Sci., 362: 1093-1105.
CrossRefDirect Link - Fabricant, D.S. and N.R. Farnsworth, 2001. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect., 109: 69-75.
CrossRefPubMedDirect Link - Gao, F.K., C.C. Dai and X.Z. Liu, 2010. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res., 4: 1346-1351.
Direct Link - Zhang, H.W., Y.C. Song and R.X. Tan, 2006. Biology and chemistry of endophytes. Nat. Prod. Rep., 23: 753-771.
CrossRefPubMedDirect Link - Tan, R.X. and W.X. Zou, 2001. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep., 18: 448-459.
PubMedDirect Link - Gangadevi, V. and J. Muthusamy, 2008. Isolation of colletotrichum gloeosporioides, a novel endophytic taxol-producing fungus from the leaves of a medicinal plant Justicia gendarussa. Mycol. Balcanica, 5: 1-4.
Direct Link - Singh, M.P., J.E. Janso, S.W. Luckman, S.F. Brady, J. Clardy, M. Greenstein and W.M. Maiese, 2000. Biological activity of guanacastepene, a novel diterpenoid antibiotic produced by an unidentified fungus CR115. J. Antibiot., 53: 256-261.
Direct Link - Guo, B., J. Dai, S. Ng, Y. Huang, C. Leong, W. Ong and B.K. Carte, 2000. Cytonic acids A and B: Novel tridepside inhibitors of hCMV protease from the endophytic fungus Cytonaema species. J. Nat. Prod., 63: 602-604.
CrossRefDirect Link - Stierle, A., G.A. Strobel and D. Stierle, 1993. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science, 260: 214-216.
CrossRefPubMedDirect Link - Li, J.Y., R. Sidhu, A. Bollon and G.A. Strobel, 1998. The stimulation of taxol production in liquid cultures of Pestalotiopsis microspora Ne32. Mycol. Res., 102: 461-464.
Direct Link - Li, J.Y., R.S. Sidhu, E. Ford, D.M. Long, W.M. Hess and G.A. Strobel, 1998. The induction of taxol production in the endophytic fungus-Periconia sp. from Torreya grandifolia. J. Ind. Microbiol., 20: 259-264.
CrossRefDirect Link - Strobel, G., B. Daisy and U. Castillo, 2005. The biological promise of microbial endophytes and their natural products. Plant Pathol. J., 42: 161-176.
CrossRefDirect Link - Wang, F., M. Xu, Q. Li, I. Sattler and W. Lin, 2010. P-aminoacetophenonic acids produced by a mangrove endophyte Streptomyces sp. (strain HK10552). Molecules, 15: 2782-2790.
CrossRefDirect Link - Gil, J. and A. Campelo-Diez, 2003. Candicidin biosynthesis in Streptomyces griseus. Applied Microbiol. Biotechnol., 60: 633-642.
CrossRefDirect Link - Yan, L.L., N.N. Han, Y.Q. Zhang, L.Y. Yu and J. Chen et al., 2010. Antimycin A18 produced by an endophytic Streptomyces albidoflavus isolated from a mangrove plant. J. Antibiotic, 63: 259-261.
PubMed - Vollmar, D., A. Thorn, I. Schuberth and S. Grond, 2009. A comprehensive view on 4-methyl-2- quinazolinamine, a new microbial alkaloid from Streptomyces of TCM plant origin. J. Antibiot., 62: 439-444.
PubMed - Ezra, D., U.F. Castillo, G.A. Strobel, W.M. Hess and H. Porter et al., 2004. Coronamycins, peptide antibiotics produced by a verticillate Streptomyces sp. (MSU-2110) endophytic on Monstera sp. Microbiology, 150: 785-793.
CrossRefDirect Link - Castillo, U.F., G.A. Strobel, E.J. Ford, W.M. Hess and H. Porter et al., 2002. Munumbicins, wide spectrum antibiotics produced by Streptomyces (NRRL 30562) endophyticon Kennedia nigriscans. Microbiology, 148: 2675-2685.
Direct Link - Taechowisan, T., A. Wanbanjob, P. Tuntiwachwuttikul and J. Liu, 2009. Anti-inflammatory activity of lansais from endophytic Streptomyces sp. SUC1 in LPS-induced RAW 264.7 cells. Food Agric. Immunol., 20: 67-77.
CrossRefDirect Link - Taechowisan, T., A. Wanbanjob, P. Tuntiwachwuttikul and J. Liu, 2010. Anti-inflammatory effects of lansai C and D cause inhibition of STAT-1 and NF-κB activations in LPS-induced RAW 264.7 Cells. Food Agric. Immunol., 21: 57-64.
CrossRef - Lin, W., L. Li, H. Fu, I. Sattler, X. Huang and S. Grabley, 2005. New cyclopentenone derivatives from an endophytic streptomyces sp. Isolated from the mangrove plant aegiceras comiculatum. J. Antibiot., 58: 594-598.
PubMed - Guan, S., S. Grabley, I. Groth, W. Lin, A. Christner, D. Guo and I. Sattler, 2005. Structure determination of germacrane-type sesquiterpene alcohols from an endophyte Streptomyces griseus subsp. Magn. Reson. Chem., 43: 1028-1031.
CrossRefDirect Link - Igarashi, Y., M. Ogawa, Y. Sato, N. Saito and R. Yoshida et al., 2000. Fistupyrone, a novel inhibitor of the infection of Chinese cabbage by Alternaria brassicicola, from Streptomyces sp. TP-A0569. J. Antibiot., 53: 1117-1122.
Direct Link - Igarashi, Y., S.S. Miura, T. Fujita and T. Furumai, 2006. Pterocidin, a cytotoxic compound from the endophytic streptomyces hygroscopicus. J. Antibiot., 59: 193-195.
CrossRef - Kim, N., J.C. Shin, W. Kim, B.Y. Hwang, B.S. Kim, Y.S. Hong and D. Lee, 2006. Cytotoxic 6 alkylsalicylic acids from the endophytic Streptomyces laceyi. J. Antibiot., 59: 797-800.
CrossRef - Ryan, P.R., K. Germaine, A. Franks, D.J. Ryan and D.N. Dowling, 2008. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett., 278: 1-9.
CrossRefDirect Link - Gunatilaka, A.A.L., 2006. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity and implications of their occurrence. J. Nat. Prod., 69: 509-526.
CrossRefDirect Link - Cragg, G.M., P.G. Grothaus and D.J. Newman, 2009. Impact of natural products on developing new anti-cancer agents-A review. Chem. Rev., 109: 3012-3043.
CrossRefPubMedDirect Link - Joseph, B. and R.M. Priya, 2011. Bioactive compounds from endophytes and their potential in pharmaceutical effect: A review. Am. J. Biochem. Mol. Biol., 1: 291-309.
CrossRefDirect Link - Lam, K.S., 2006. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol., 9: 245-251.
CrossRefPubMedDirect Link - Tharek, M., K. Dzulaikha, S. Salwani, H.G. Amir and N. Najimudin, 2011. Ascending endophytic migration of locally isolated diazotroph, Enterobacter sp. Strain USML2 in rice. Biotechnology, 10: 521-527.
CrossRef - Khucharoenphaisan, K., N. Sripairoj and K. Sinma, 2012. Isolation and identification of actinomycetes from termite's gut against human pathogen. Asian J. Anim. Vet. Adv., 7: 68-73.
CrossRefDirect Link - Raja, M.M.M., A. Raja, M.M. Imran and E.V. Ugin, 2011. Anti mycobacterial activity of actinomycetes producing mycothiol. Singapore J. Sci. Res., 1: 144-148.
Direct Link - Castillo, U., J.K. Harper, G.A. Strobel, J. Sears and K. Alesi et al., 2003. Kakadumycins, novel antibiotics from Streptomyces sp. NRRL 30566, an endophyte of Grevillea pteridifolia. FEMS Microbiol. Lett., 224: 183-190.
CrossRefDirect Link