
Houneida Attia
Department of Biology, Faculty of Science, Taif University, Taif, Kingdom of Saudi Arabia
LiveDNA: 216.30371
Khalid H Alamer
Department of Biology, Science and Arts College-Rabigh Campus, King Abdulaziz University, Jeddah, Kingdom of Saudi Arabia
Chayma Ouhibi
Physiology and Biochemistry of Plant Response to Abiotic Stress, Faculty of Science of Tunis, Tunis El Manar University, 2092 Tunis, Tunisia
Samia Oueslati
Biotechnology Center of Borj-Cedria, University of Tunis El Manar, Tunis, Tunisia
Mokhtar Lachaal
Physiology and Biochemistry of Plant Response to Abiotic Stress, Faculty of Science of Tunis, Tunis El Manar University, 2092 Tunis, Tunisia
International Journal of Botany
Year: 2020 | Volume: 16 | Issue: 1 | Page No.: 1-8
DOI: 10.3923/ijb.2020.1.8







ABSTRACT
Background and Objective: Salinity is a key factor limiting agricultural production around the world and often occurs at the same time as drought stress. Conventional selection in saline conditions has generally failed, in part due to the large variability of natural saline soils resulting from different salinity and drought. The purpose of the work was to assess the combined effect of drought and salinity in 2 pea (Pisum sativum L.) cultivars cv Lincoln and cv Douce de provence. Materials and Methods: Seedlings were cultured on liquid medium without or with NaCl, 75 mM in controlled conditions. Fourteen days old, salt treated plants were subjected to drought (PEG 6000). After 7 days of treatment, growth parameters, mineral content (Na+, Cl–, K+, Ca2+ and Mg2+) and chlorophyll content were measured. Peroxidases and catalase activities was assayed on gels. Results: The combined action of drought and salt restricted leaf biomass more pronounced in cv D. provence than in cv Lincoln. This growth decrease was accompanied by a restriction in leaf hydration and K+ uptake, concomitant with low Na+ accumulation in cv D. provence leaves. Considering the response of antioxidant enzymes to combined stress, leaves guaiacol peroxidase activity showed an increase as compared to control in much more pronounced in cv Lincoln than in cv D. provence. Conclusion: Both pea cultivars exhibited a stimulation of catalase activity with higher in cv D. Provence than in cv Lincoln. Thus, cv D. lincoln had a higher osmotic drought stress than cv D. provence.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Houneida Attia, Khalid H Alamer, Chayma Ouhibi, Samia Oueslati and Mokhtar Lachaal, 2020. Interaction Between Salt Stress and Drought Stress on Some Physiological Parameters in Two Pea Cultivars. International Journal of Botany, 16: 1-8.
DOI: 10.3923/ijb.2020.1.8
URL: https://scialert.net/abstract/?doi=ijb.2020.1.8
DOI: 10.3923/ijb.2020.1.8
URL: https://scialert.net/abstract/?doi=ijb.2020.1.8
INTRODUCTION
Field pea (Pisum sativum L.) is widely cultivated as an important pulse crop on a global basis for human nutrition and stock-feed consumption. This species is also used for forage production1, in rotations with cereals for provision of soil nitrogen2, and to provide disease breaks.
Salt and water stresses due to drought and soil salinity are the extremely important abiotic stresses in limiting the yield of food crops in the world3. Soil salinity is a grave danger to global crop that limits agricultural production globally4, and is present in 6% of the world’s soil area and 20% of irrigated soil5. In addition to a natural increase in saline soils especially in arid and semi-arid areas due to partial precipitation, high temperature and evapotranspiration, insufficient organization of freshwater and clearing of soil for dry land cultivation contributes to augmented salt stress in vegetation. Because salinity is mainly important for arid and semi-arid parts and because of the increasing frequency of droughts in many other parts of the world, salinity usually occurs at the same time as drought stress6. Both abiotic stresses decrease the potential water in the soil and the ability of plants to absorb water, reducing the rate of cell expansion in growing tissues, the stomatal conductance and thus photosynthetic rate3. While dehydration also reduces nutrient availability, nutrient uptake by roots and transport from roots to shoots, saline soils further diminish plant development most often through the vegetative phase due to definite ion toxicities and ionic imbalances7 which was projected as the 2 time representation of saline stress8.
Since salinity is mainly significant for arid and semi-arid areas, plants have developed tolerance strategies for both salt and water stress9. Salinity often induces osmotic adjustment, which is considered an important mechanism for maintaining water absorption and cell turgor under stress conditions10.
The improvement of the antioxidant defense system is considered effective in the development of resistance and adaptive characteristics in plants against drought stress. Many research findings confirm that enhanced activities of antioxidant enzyme components decrease oxidative damage, and develop and improve drought tolerance and plant resistance11,12.
In this study we investigated the effect of osmotic stress induced by a combination of salt and PEG 6000 on the growth and antioxidant enzymes of two cultivars of pea: Lincoln and D. provence.
MATERIALS AND METHODS
Plant materials, growth conditions and salt and drought treatments: The seeds of two pea (Pisum sativum L.) of commercial cultivars widely used in Tunisia come from the Seed Legume Laboratory of the Tunisian National Institute for Agriculture Research. The experiment was conducted from October, 2017 to October, 2018. Douce de Provence (D. Provence thereafter) has a shorter breeding cycle (45 days) than Lincoln (60 days). The seeds were surface sterilized with 96% (v/v) ethanol for 3 min and 2% (w/v) sodium hypochlorite for 5 min. After rinsing they were allowed to germinate on a filter paper in 9 cm diameter Petri dishes, moistened with distilled water or NaCl, 75 mM. The Petri dishes were stored for 4 days at room temperature (24±2°C) under dark conditions. Vigorous seedlings were selected and transferred to plastic pots and were grown in hydroponic culture containing 1/5 strength nutrient solution of Long Ashton13. The NaCl, 75 mM was added to the nutritive solution for half of plants. The conditions of the growth chamber were set at 25/18°C day/night, 80% relative humidity and 150 mmol m–2 sec–1 light intensity with a photoperiod of 14 h.
Fourteen days later, plants of salt medium were separated into two groups: The first was maintained on the same sline medium (S) and the second was supplemented with polyethylene glycol (PEG 6000) having a hydric potential -0.5 MPa. This solution is named (S+D) medium. Eight plants of each treatment were harvested 7 days later.
Biomass measurements and inorganic ions assays: At the harvest, individual plants were divided into leaf, stem and root fractions. Fresh weights (FW) were immediately determined. The samples were then dried in a forced draft oven at 70°C for 48 h and dry weights (DW) were determined. Leaf surface area was measured using a scanner and Optimas® software. Water content was calculated utilizing the following equation with FW, fresh weight and DW, dry weight:
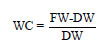
Ions were extracted with 0.5% HNO3. Sodium and potassium were assayed by flame photometry (Eppendorf) with butane-air flame, calcium by acetylene-air flame, magnesium by atomic absorption spectrophotometry (VARIAN 220 FS) and Cl– by coulometry (Butcher Cotlove chloridometer), according to manufacturers’ instructions.
Analysis of total chlorophyll content: Total chlorophylls were extracted from fresh leaves of 8 different individuals in 80% acetone in the dark and assayed photometrically according to Arnon14.
Enzyme extraction and assays: Leaf sample (200 mg) was frozen in liquid nitrogen and finely ground by pestle in a chilled motor, the frozen powder was added to 0.6 mL of phosphate buffer (pH 7.0). The homogenate was centrifuged at 13000×g for 30 min at 4°C and supernatant was used for enzymatic activity measurements. The protein concentration was determined according to Bradford15 using bovine serum albumin as a standard.
Enzyme activities were analyzed after native gel electrophoresis of the supernatant. Guaiacol peroxidase (GPX, EC 1.11.1.7), stacking gel was 5% acrylamide in pH 6.8, 0.5 M Tris-HCl buffer, and resolving gel was 5% acrylamide in pH 8.8, 1.5 M Tris-HCl buffer. Catalase (EC 1.11.1.6) was identified on 6% acrylamide gels. The same Tris-glycine analysis buffer 23 mM, pH 8.3 was used for the three enzymatic systems. The migration was carried at 4°C at 120 V, with a pH 4.0, 8 mM glycine buffer as analysis buffer, using the Mini-protean Bio-Rad system. To show the activity of peroxidase, the gels were incubated for 30 min in the dark in 50 mL of pH 4, 100 mM acetate buffer containing 1% (w/v) guaiacol. They were then transferred to 50 mL acetate buffer (100 mM, pH 5) containing 25 mg of 3-amino-9-ethylcarbazole, 25 mL of N,N-dimethyl formamide (99%, v/v), 1 mL 0.1 M CaCl2 and 0.5 mL H2O2 (30%, v/v). Gel staining for catalase activity was performed according to Chandlee and Scandalios16 after pretreatment in 0.01% (v/v) H2O2 for 10 min. The staining mixture contained 1% (w/v) FeCl3 and 1% (w/v) K3Fe(CN)6 in distilled water.
Statistical analysis: Statistical analysis were performed with StatisticaTM Software, using ANOVA and the Newman-Keuls test for post hoc mean comparison.
RESULTS
Biomass production: After 21 d of culture in salt, the leave dry weight exhibited marked reductions only in D. provence (70% of control) (Fig. 1). cv Lincoln appeared more tolerant to salinity in the nutrient solution than cv D. provence. Under the combined action of drought and salt, D. provence leaves were more affected than those from Lincoln and the decrease in dry weight reached 48 and 18% of control, respectively (Fig. 1).
The water content on a dry matter basis of both pea cultivars decreased with salinity and when both stresses were combined, with much more marked impact for the second constraint and for cv D. provence (48% of control) (Fig. 1). Moreover, we noticed early leaf wilting of the latter cultivar on PEG, announcing osmotic effects due to this organic osmoticum.
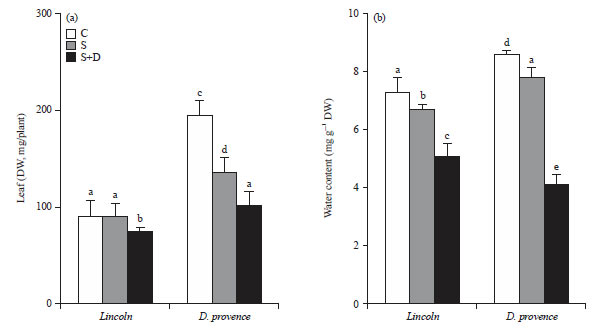 | |
| Fig. 1(a-b): | Combined effect of NaCl and PEG 6000 on (a) Leaf dry weight (DW, mg/plant) and (b) Water content (mL g–1 DW) in 2 pea cultivars, Lincoln and D. provence |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000 (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000, mean of 8 plants and confidence intervals for p = 0.05, mean sharing a same letter are not significantly different at p = 0.01 (ANOVA and mean comparison with Newman-Keuls test) | |
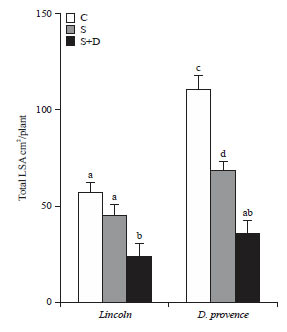 | |
| Fig. 2: | Combined effect of NaCl and PEG 6000 on total leaf surface area (cm2/plant) in 2 pea cultivars, Lincoln and D. provence |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000. (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000, mean of eight plants and confidence intervals for p = 0.05, mean sharing a same letter are not significantly different at p = 0.01 (ANOVA and mean comparison with Newman-Keuls test) | |
Total area of leaves/plant was limited in the presence of NaCl to about 81 and 61% of control, in Lincoln and D. provence, respectively (Fig. 2). However, this effect was not statistically significant (p = 0.05) in Lincoln. Salt presence in the culture medium of plants subjected to drought strongly restricted leaf expansion, more active in cv D. provence (32% of control) than in cv Lincoln (42% of control) (Fig. 2).
Salt accumulation: Great contents of Na+ and Cl– were accumulated in treated plants, more than 1.5 mmol g–1 DW in leaves of both cultivars (Fig. 3). Combined effect of drought and salinity (S+D treatment) greatly reduced sodium and chloride accumulation in cv Lincoln leaves, as compared to cv D. provence leaves.
Mineral nutrition: In the leaves of cv Lincoln and cv D. provence, accumulation of K+, Ca2+ and Mg2+ was reduced by salt treatment compared to control in both pea cultivars, Lincoln and D. provence (Fig. 4). Combined salinity+drought stress partially mitigated the limitations caused by NaCl to potassium content only in Lincoln. In leaves of D. provence, it gave rise to an accumulation of K+ almost equal to that of the salt treatment.
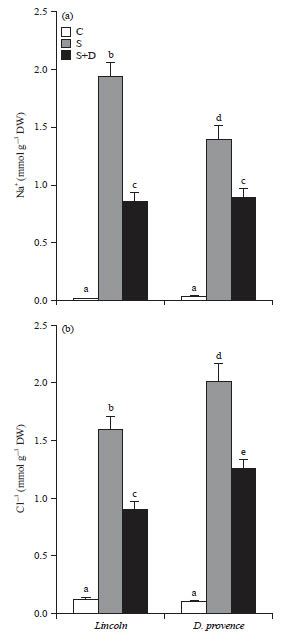 | |
| Fig. 3(a-b): | Combined effect of NaCl and PEG 6000 on salt accumulation in two pea cultivars, Lincoln and D. provence |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000 (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000, mean of eight plants and confidence intervals for p = 0.05, mean sharing a same letter are not significantly different at p = 0.01 (ANOVA and mean comparison with Newman-Keuls test) | |
In combined salt and drought treatment, Ca2+ and Mg2+ accumulation was similar to that of salt-treated plants for both cultivars with a small decrease for Ca2+ accumulation in D. provence leaves (Fig. 4).
Total chlorophylls: Treatment with NaCl did not change total chlorophylls in Lincoln leaves, but stimulated an increase in cv D. provence (Fig. 5). The effects of the two constraints when combined increased the chlorophyll content in both cultivars.
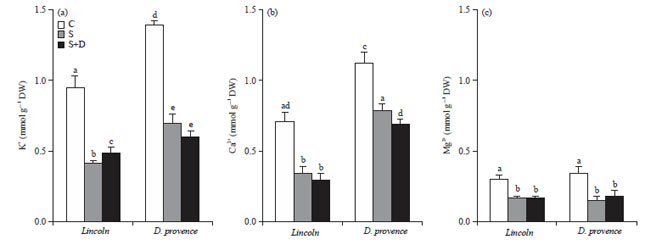 | |
| Fig. 4(a-c): | Combined effect of NaCl and PEG 6000 on mineral nutrition (a) K+, (b) Ca+2 and (c) Mg2+ in two pea cultivars, Lincoln and D. provence |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000 (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000, mean of 8 plants and confidence intervals for p = 0.05, mean sharing a same letter are not significantly different at p = 0.01 (ANOVA and mean comparison with Newman-Keuls test) | |
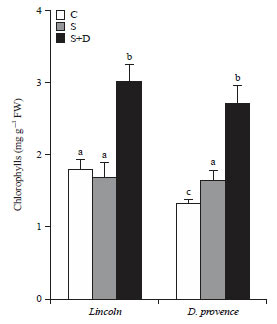 | |
| Fig. 5: | Combined effect of NaCl and PEG 6000 on total chlorophylls in 2 pea cultivars, Lincoln and D. provence |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000 (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000, mean of eight plants and confidence intervals for p = 0.05. Mean sharing a same letter are not significantly different at p = 0.01 (ANOVA and mean comparison with Newman-Keuls test) | |
Leaf antioxidant enzyme activities: The electrophoretic examination illustrated that independently of the treatment, GPX activity was higher in cv Lincoln than in cv D. provence.
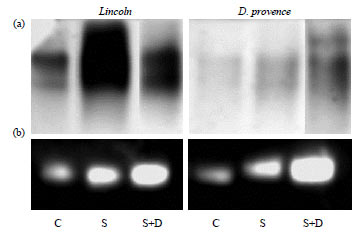 | |
| Fig. 6(a-b): | Combined effect of NaCl and PEG 6000 on leaf antioxidant enzyme activities in 2 pea cultivars, Lincoln and D. provence. Electropherograms of antioxidant enzymes in extracts from pea leaves, (a) Guaiacol peroxidase and (b) Catalase |
Fourteen days old salt-treated plants were grown for 7 days in the absence (S) or in the presence (S+G) of PEG 6000 (C) plants grown in the absence of both NaCl (75 mM) and PEG 6000 | |
In addition, this activity considerably stimulated in cv Lincoln under NaCl; however, in cv D. provence, it appears to remain unaffected or slightly stimulated (Fig. 6a). Under both water deficit and salinity stress, GPX activity decreased in cv Lincoln compared to salt treatment, but increased in cv D. provence (Fig. 6a).
Catalase (CAT) is implicated in scavenging hydrogen peroxide created by dismutation of superoxide anions catalyzed by superoxide dismutase enzyme. The electrophoretic examination discovered higher CAT activity in response to salt stress and much more elevated under the interaction stress of salinity and drought both in cultivars (Fig. 6b).
DISCUSSION
Salinity and drought are 2 environmental stresses that often occur concurrently in arid areas. The capacity to overcome various and concurrent stresses is of big importance for the plant development and endurance in environmental constraints17. Despite the presence of different methods for application of water stress, polyethylene glycol 6000 was used in this study for the reason that a cytorrhytic instead of plasmolytic reduce water potential effect can be imposed with solutions including a high molecular weight solute such as polyethylene glycol with molecular mass 6000 or above18. Polyethylene glycol 6000 or above cannot penetrate the pores of plant cells19. In addition, PEG is a good choice to impose a lower water potential than mannitol, an often used solute, as it has been shown that mannitol is absorbed by the plant cells and may have adverse effects on development20.
Destructive conventional samplings do not allow dissecting the three mechanisms of tolerance to salinity: Excluding Na+, tissue tolerance against Na+ and osmotic tolerance21. In a previous work22, seedlings of 2 pea cultivars Lincoln and D. provence grow for 14 days on ordinary medium were disputed for 21 days with NaCl using a split-root system. Processing complete salt (S/S) resulted growth inhibition more marked in cv Lincoln than in cv D. provence22. The opposite was observed in this work, explaining the effect of seeds irrigation with saline solution. Combined effect of drought and salt stresses causes a decrease in dry weight, especially in cv D. provence which shows a particular sensitivity to this organic osmoticum. In general, it is accepted that the key destructive factors at the cellular stage in water-deficit and salt are the toxic and osmotic effects of salt23.
The interaction between salinity and drought stresses diminished water potential in barley24. These results showed that salinity considerably diminished water content in leaves, with a remarkable impact when plants subjected to NaCl and PEG 6000 in the culture medium, more pronounced in cv D. provence than in cv Lincoln (Fig. 1). The main differentiation among low water potential situation due to salinity against water deficit is the whole quantity of water accessible. Plants have a ability to regulate their osmotic potential, which avert defeat of turgor and produce a minor water potential, which allows the plants to access water in soil solution for development25.
As for ion effects, concentrations of Na+ and Cl– accumulation in plants subjected to combined stress were greatly lower than those treated only with NaCl. This may propose that salinity cannot lead to effect of ions due to salinity under combined stress, signifying that the exclusion of Na and Cl– cannot always be a tolerance of plants to salt stress, which is maintained by Genc et al.26.
The increased salt treatments cause an increase in Na+ and Cl– and a decrease in Ca2+, K+ and Mg2+ leaves in several plants27. Plant physiology studies have shown that throughout salt stress, early symptoms on plant development are due to water stress, while effects of salt-specific only occur much later (from a few days to a few days)28,29.
Chlorophyll content in leaves is a key parameter for characterization of physiological yield of plants30. Decrease in photosynthesis is often related to the decrease of pigment content caused by inhibition of their synthesis or stimulated damage as well as chloroplasts destruction31. In the present study, salt stress did not change total chlorophylls in leaves of cv Lincoln, but stimulated an increase in cv D. provence ones (Fig. 5). The concentrations of total chlorophylls in plant tissue when the two constraints where combined were higher in the two pea cultivars which can be attributed to a capacity and carbohydrate formation under water deficit and salinity stresses. Augmentation of chlorophyll content as a result of salt constraint was observed in a previous study32.
Environmental constraints restrictive photosynthesis may augment oxygen-induced cellular damage as a result of augmented ROS generation33,34. The level of damage by ROS is related to the equilibrium between ROS product and its elimination by these antioxidant systems35,36. It has been found that a relationship between the antioxidant capacity and NaCl tolerance was established by assessment of tolerant cultivars with sensitive cultivars in pea plants22. In the present study, the peroxidases and catalase activity was increased in leaves under salt stress and combination of 2 stresses stress, but pea cultivars response pattern was different. The most induction of enzyme activity was observed in salt-treated cv Lincoln and in cv D. provence under drought stress in combination with salinity. The results suggest that different regulatory mechanisms may exist in the regulation of antioxidant enzyme activity under different situations. Several studies have related oxidative defense systems with salt tolerance. On the other hand, the regulation of a number of oxidative defenses can be a secondary response to salt stress37. It has been shown that the production of organic osmolytes through plant stresses such as NaCl or water deficit develops osmoregulation and protects the enzymatic function in the cytosol38, as validated in recent transgenic studies39 including tobacco40 and potato41. Little study has been conducted to date to classify QTLs controlling creation of different osmoprotectants, except proline accumulation studies42. The efficacy of various osmolytes in improving salinity tolerance may also vary from one species to another43 which may provide another way to identify genes that are useful for pea tolerance to salt stress.
CONCLUSION
In conclusion, an important finding of our study is that a higher sensitivity to osmotic drought stress, induced by a combination of NaCl and PEG 6000, of cv D. provence compared to cv Lincoln was evident from the higher reduction in growth, water content, mineral nutrition mainly K+ and Ca2+ accumulation, which was concomitant with low Na+ accumulation in cv D. provence leaves.
SIGNIFICANCE STATEMENT
This study discovers the Interaction between salt stress and drought stress in 2 pea cultivars Lincoln and D. provence. These 2 cultivars showed, in a previous study, different responses of plants subjected to salt stress. This study will help the researcher to uncover the most tolerant cultivar for salt responses as well as for water stress and the combined effect on a species growing as a significant crop on a universal basis for human nutrition. Thus, a cultivar shows a degree of tolerance to combined stress, may be retained.
ACKNOWLEDGMENTS
Authors are indebted to Dr. Hasna Ellouzi for her help. Attia H. is grateful to chemistry lab of Biotechnology Center of Borj-Cedria in Tunisia. She specially thanks Prof. Chedly Abdelly for hosting her in his research groups.
REFERENCES
- Kocer, A. and S. Albayrak, 2012. Determination of forage yield and quality of pea (Pisum sativum L.) mixtures with oat and barley. Turk. J. Field Crops, 17: 96-99.
Direct Link - Omokanye, A.T., F.M. Kelleher and A. McInnes, 2011. Low-Input cropping systems and nitrogen fertilizer effects on crop production: Soil nitrogen dynamics and efficiency of nitrogen use in maize crop. Am.-Eurasian J. Agric. Environ. Sci., 11: 282-295.
Direct Link - Munns, R., 2011. Plant adaptations to salt and water stress: Differences and commonalities. Adv. Bot. Res., 557: 1-32.
CrossRefDirect Link - Tavakkoli, E., P. Rengasamy and G.K. McDonald, 2010. The response of barley to salinity stress differs between hydroponic and soil systems. Funct. Plant Biol., 37: 621-633.
CrossRefDirect Link - Munns, R., 2005. Genes and salt tolerance: Bringing them together. New Phytol., 167: 645-663.
CrossRefPubMedDirect Link - Hu, Y., Z. Burucs and U. Schmidhalter, 2006. Short-term effect of drought and salinity on growth and mineral elements in wheat seedlings. J. Plant Nutr., 29: 2227-2243.
CrossRefDirect Link - Hu, Y. and U. Schmidhalter, 2005. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci., 168: 541-549.
CrossRefDirect Link - Munns, R., 1993. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ., 16: 15-24.
CrossRefDirect Link - Munns, R. and M. Tester, 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59: 651-681.
CrossRefDirect Link - Chaves, M.M., J. Flexas and C. Pinheiro, 2009. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot., 103: 551-560.
CrossRefDirect Link - De Carvalho, M.H.C., 2008. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal Behav., 3: 156-165.
CrossRefPubMedDirect Link - Yazdanpanah, S., A. Baghizadeh and F. Abbassi, 2011. The interaction between drought stress and salicylic and ascorbic acids on some biochemical characteristics of Satureja hortensis. Afr. J. Agric. Res., 6: 798-807.
Direct Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chandlee, J.M. and J.G. Scandalios, 1983. Gene expression during early kernel development in Zea mays. Dev. Genet., 4: 99-115.
CrossRefDirect Link - Lichtenthaler, H.K., 1996. Vegetation stress: An introduction to the stress concept in plants. J. Plant Physiol., 148: 4-14.
CrossRefDirect Link - Verslues, P.E., M. Agarwal, S. Katiyar-Agarwal, J. Zhu and J.K. Zhu, 2006. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J., 45: 523-539.
CrossRefPubMedDirect Link - Oertli, J.J., 1985. The response of plant cells to different forms of moisture stress. J. Plant Physiol., 121: 295-300.
CrossRefDirect Link - Hohl, M. and P. Schopfer, 1991. Water relations of growing maize coleoptiles: Comparison between mannitol and polyethylene glycol 6000 as external osmotica for adjusting turgor pressure. Plant Physiol., 95: 716-722.
CrossRefDirect Link - Rajendran, K., M. Tester and S.J. Roy, 2009. Quantifying the three main components of salinity tolerance in cereals. Plant Cell Environ., 32: 237-249.
CrossRefDirect Link - Attia, H., S. Nouaili, A. Soltani and M. Lachaâl, 2009. Comparison of the responses to NaCl stress of two pea cultivars using split-root system. Sci. Hortic., 123: 164-169.
CrossRefDirect Link - Zhu, J.K., 2002. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol., 53: 247-273.
CrossRefPubMedDirect Link - Ahmed, I.M., H.X. Dai, W. Zheng, F.B. Cao, G.P. Zhang, D.F. Sun and F.B. Wu, 2013. Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiol. Biochem., 63: 49-60.
CrossRefDirect Link - Genc, Y., G.K. McDonald and M. Tester, 2007. Reassessment of tissue Na+ concentration as a criterion for salinity tolerance in bread wheat. Plant Cell Environ., 30: 1486-1498.
CrossRef - Khan, M.A., I.A. Ungar and A.M. Showalter, 2000. Effects of sodium chloride treatments on growth and ion accumulation of the halophyte Haloxylon recurvum. Commun. Soil Sci. Plant Anal., 31: 2763-2774.
CrossRefDirect Link - Denby, K. and C. Gehring, 2005. Engineering drought and salinity tolerance in plants: lessons from genome-wide expression profiling in Arabidopsis. Trends Biotechnol., 23: 547-552.
CrossRef - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Lu, S., X. Lu, W. Zhao, Y. Liu, Z. Wang and K. Omasa, 2015. Comparing vegetation indices for remote chlorophyll measurement of white poplar and Chinese elm leaves with different adaxial and abaxial surfaces. J. Exp. Bot., 66: 5625-5637.
CrossRef - Wang, Y. and N. Nii, 2000. Changes in chlorophyll, ribulose bisphosphate carboxylase-oxygenase, glycine betaine content, photosynthesis and transpiration in Amaranthus tricolor leaves during salt stress. J. Hortic. Sci. Biotechnol., 75: 623-627.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Neill, S., R. Desikan and J. Hancock, 2002. Hydrogen peroxide signalling. Curr. Opin. Plant Biol., 5: 388-395.
CrossRefDirect Link - Demiral, T. and I. Turkan, 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot., 53: 247-257.
CrossRefDirect Link - Khan, M.H. and S.K. Panda, 2008. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Planta., 30: 81-89.
CrossRefDirect Link - Ashraf, M., 2009. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv., 27: 84-93.
CrossRefPubMedDirect Link - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Hussain, S.S., H. Raza, I. Afzal and M.A. Kayani, 2011. Transgenic plants for abiotic stress tolerance: Current status. Arch. Agron. Soil Sci., 58: 693-721.
CrossRef - Ziaf, K., R. Loukehaich, P.J. Gong, H. Liu and J.Q.Q. Han et al., 2011. A multiple stress-responsive gene ERD15 from Solanum pennellii confers stress tolerance in tobacco. Plant Cell Physiol., 52: 1055-1067.
CrossRefDirect Link - Rahnama, H., H. Vakilian, H. Fahimi and B. Ghareyazie, 2011. Enhanced salt stress tolerance in transgenic potato plants (Solanum tuberosum L.) expressing a bacterial mtlD gene. Acta Physiol Plant., 33: 1521-1532.
CrossRef - Asharaf, M. and M.R. Foolad, 2012. Crop breeding for salt tolerance in the era of molecular markers and marker‐assisted selection. Plant Breed., 132: 10-20.
CrossRefDirect Link