
Omesh Bajpai
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
LiveDNA: 91.4964
ORCID: 0000-0002-8440-6467
Anoop Kumar
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
Ashish K. Mishra
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
Nayan Sahu
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
Soumit K. Behera
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
Lal Babu Chaudhary
Plant Diversity, Systematics and Herbarium Division, CSIR-National Botanical Research Institute, Rana Pratap Marg, Lucknow, Uttar Pradesh-226 001, India
International Journal of Botany
Year: 2012 | Volume: 8 | Issue: 2 | Page No.: 66-72







ABSTRACT
The phenological studies of two tree species Shorea robusta Gaertn. f. and Ficus hispida L.f. occurring in Katerniaghat Wildlife Sanctuary (KWS), a tropical moist deciduous forest in Northern India along Indo-Nepal border has been carried out to gather information about the different phenophases of the plants. Both the candidate species are the co-dominant constituents of the sanctuary area. For each species ten trees were selected and a total of 160 twigs were tagged to collect phenological data. The initiation and completion of different phenophases such as leaf bud initiation, leaf emergence, young leaf formation, leaf fall, flower bud development, formation of flowers and fruits were recorded at monthly basis. In addition, the number of leaf buds, young leaves, matured leaves and total leaves has also been recorded. The individual leaf area was measured by using graph paper once they became stable in their growth and ultimately average leaf area was obtained. In both the species leaf bud busting and flowering/figing initiate in post winter (March to April) and maximum leaf fall was recorded in the post monsoon season (November to February). In Shorea robusta the young fruits start to appear in dry season (May) and get matured before the arrival of the monsoon (June) so that the seeds get proper moist condition to germinate.
PDF Abstract XML References Citation
Received: January 30, 2012;
Accepted: April 18, 2012;
Published: July 11, 2012
How to cite this article
Omesh Bajpai, Anoop Kumar, Ashish K. Mishra, Nayan Sahu, Soumit K. Behera and Lal Babu Chaudhary, 2012. Phenological Study of Two Dominant Tree Species in Tropical Moist Deciduous Forest from the Northern India. International Journal of Botany, 8: 66-72.
DOI: 10.3923/ijb.2012.66.72
URL: https://scialert.net/abstract/?doi=ijb.2012.66.72
DOI: 10.3923/ijb.2012.66.72
URL: https://scialert.net/abstract/?doi=ijb.2012.66.72
INTRODUCTION
Phenology of tree in any ecosystem and community strongly determines the flowering periods which is indirectly dependent on the environmental variations (Rivera et al., 2002; Hamann, 2004; Zhang et al., 2006). The dependence of plants on the climate for the succession of different life phases increases the significance of phenological studies in its connection with climate change and global climate monitoring (Schwartz, 1999; Mezel, 2002; Schwartz et al., 2006). The phenological studies are instrumental in assessing the response of plants and plant community against climatic disturbances. Although, the climate change is being recognised at global scale however, its assessment at the regional level may also be used to predict changes in the climate (Anonymous, 2001). The effect of climate change may be assessed by correlating seasonal climatic conditions and the different phenophases of the plants (Kushwaha and Singh, 2008). In tropical Asian forests, climate and phenology interactions are poorly understood (Corlett and Lafrankie, 1998; Borchert et al., 2002). Regional studies of plant phenology even carried out in small area are equally important and at the same time it require low budget also. Once the data is generated, these studies can throw light on regional peculiarities which can be utilised at national and international levels (Menzel and Estella, 2001).
India with a wide range of variations in climate, altitude and physiography exhibits enormous variations in the life cycle of plants of different regions (Koul and Bhatnagar, 2005). These variations are used to find out their effect on phenology worldwide. However, the phenological studies carried out in India are fragmentary and inconclusive (Koul and Bhatnagar, 2005). Precise phenological information with respect to flowering and fruiting evaluated against leafing and leafless periods is scarce in tropical deciduous forests in India which account for about 46% of the forested land in the country (Singh and Singh, 1988). The phenology of tree species in tropical dry deciduous forests has given conclusive knowledge about vegetation’s annual pattern of vegetative and floral biology (Prasad and Hegde, 1986; Singh and Singh, 1988, 1992a; Khan, 1999; Singh and Kushwaha, 2005a, b, 2006; Kushwaha and Singh, 2005; Yadav and Yadav, 2008). This information is useful to assess the influence of phenological events or faunal feeding, movement pattern and sociality (Leigh Jr and Windsor, 1982; Wada, 1983; Appanah, 1985; Coates-Estrada and Estrada, 1986) along with the climate change (Schwartz, 1999; Mezel, 2002).
The Terai region in the Himalayan foot hills from Uttarakhand to W. Bengal in India has lost majority of its natural Tropical Moist Deciduous Forests in last century due to anthropogenic pressure and unsustainable forest management. The native vegetation of this area is now shrunken and restricted to the protected areas, i.e. national parks and wildlife sanctuaries. The study site Katerniaghat Wildlife Sanctuary was selected due to its natural Sal and miscellaneous forest representing tropical moist deciduous vegetation and being a sanctuary it is a less disturbed area than any other adjoining forest in Terai region. Hence, in the present study two dominant tree species Shorea robusta Gaertn. f. and Ficus hispida L. f. occurring in Katerniaghat Wildlife Sanctuary (KWS) have been selected for their quantitative phenological studies.
The main aim of this quantitative phenological study was to assess the different phenological events in selected species of tropical moist deciduous forest to utilise them to understand the impact of climate changes on the plant species. Since, such type of studies has not been carried out properly from the tropical moist regions of the northern India, hence the data provided here may be utilise for further climate change assessment.
MATERIALS AND METHODS
Study area: The study was carried out at Katerniaghat Wildlife Sanctuary (KWS) (27°41’ -27° 56’ N and 81°48’ -81°56’ E; elevation 116-165 m) which is situated in the Himalayan Terai Region in the Bahraich district of Uttar Pradesh, India (Fig. 1). This tropical moist deciduous forest is spread over an area of 40 km long and 10 km wide range. Katerniaghat Wildlife Sanctuary consists of two major forest types (1) The Sal forest and (2) The miscellaneous forest (Champion and Seth, 1968) which basically falls under the Terai-bhabar subdivision of the Upper Gangetic Plain (Rodger and Panwar, 1988). The sanctuary area is made up of the alluvial soil of Kaudiyala and Saryu rivers flowing adjoining to it. On the geological point of view the sanctuary area has been divided into high and low land areas. The high land area has been further divided into Varanasi Old Alluvial Plains and High Lands of Ghaghara, Saryu and its tributaries.
The climate of the area is monsoonal. There are three seasonal variations in the area: winter (November to February), summer (April to June) and warm-rainy (July to September). March and October are considered as transition months between seasons. The mean maximum temperature varies from 22°C in January to 40°C in May and the mean minimum temperature varies from 8°C in January to 27°C in June. The annual rainfall ranges from 36 to 142 mm in winter, 34 to 662 mm in summer and 1294 to 1689 mm in warm-rainy seasons (Fig. 2).
The vegetation of the sanctuary area is tropical moist deciduous type and may be divided into upper stratum, lower stratum and ground vegetation. The upper stratum which receives the full solar radiation comprises trees like Shorea robusta, Tectona grandis, Terminalia elliptica, Madhuca indica, Ficus benghalensis, Ficus racemosa, Bombax ceiba, Sterculia villosa, Lannea coromandelica etc. The lower stratum is represented by Hymenodictyon excelsum, Syzygium cumini, Ehretia laevis, Lagerstroemia parviflora, Diospyros melanoxylon, Mallotus philippensis, Mallotus nudiflorus, Ficus hispida, Streblus asper etc. The ground vegetation where the minimal light is available chiefly consists of Murraya koenigii, Glycosmis pentaphylla, Clerodendrum viscosum, Grewia hirsuta, Lantana camara etc. The candidate tree species (Shorea robusta and Ficus hispida) are the dominant constituents of the forest associated with Tectona grandis, Mallotus philippensis, Syzygium cumini, Ficus racemosa, Terminalia elliptica, Ehretia laevis etc.
Field sampling and data collection: A specific number is given to all 10 selected sample trees for both the species. These trees were selected on the basis of their diameter (70±15 cm for Shorea robusta and 30±8 cm for Ficus hispida). In each tree four major branches were selected in all four directions and tagged. On each tagged branch four healthy twigs were selected and tagged and on each twigs three to five leaves were tagged when they appeared. The observations were recorded on initiation and completion of phenological events of leaf flush, flowering, fruiting etc. In the experiment, time of leaf emergence, young leaf formation, leaf fall initiation, number of leaf buds, number of young leaves, number of mature leaves, number of total leaves and average area of leaves have been recorded. The leaf area was measured by using graph paper once they became stable in growth. When no increment was observed in the leaf area, it was considered as the mature leaf.
Data analysis: A total of 160 twigs from 10 trees under each species were marked for the observation. The average leaf area of a twig has been calculated by multiplying the mean leaf area of 3-5 leaves with total number of leaves. The phenological data were collected at the interval of one month in the mid of the every month.
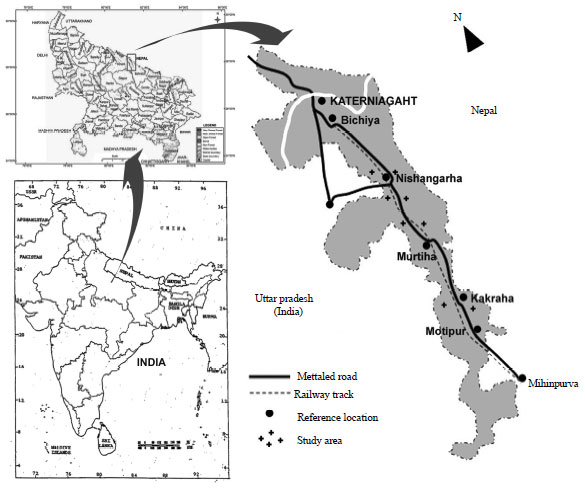 | |
| Fig. 1: | Location map of study site |
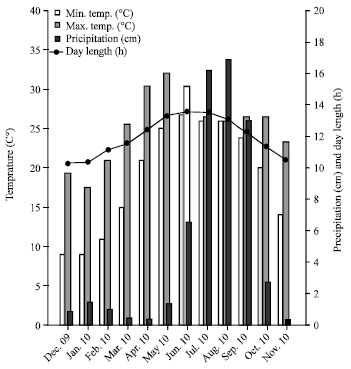 | |
| Fig. 2: | Variation in climatic condition at KWS |
The phenophases observed in a month was assumed to appear first time before fifteen days than the observation (Kushwaha and Singh, 2005). The synchrony index between selected tree specimens and whole population was calculated for both candidate tree species (Devineau, 1999).
RESULTS
In Shorea robusta, the leaf bud busting initiated in the end of March with the increment of day light duration and average minimum and maximum temperature. In the next month (i.e. in the month of April) the young leaves become jubilant on the twigs. These newly emerged leaves take about one to two months to get matured. The average age of the leaves in the species has been observed between 9-11 months. In the sanctuary area the Shorea robusta shows minimum number of total matured leaves in the month of April while the maximum in the month of June. The average area of a leaf (108.79±16.05 cm2) was measured which was utilised to calculate the average leaf area of all leaves present on a twig by multiplying the average leaf area of single leaf with total number of leaves. The average leaf area has been observed minimum in April while maximum in the month of June. The synchrony between selected tree specimens and whole population was found 0.88 for leaf-flush and 0.86 for leaf-fall. Almost every month a few leaves fall down from the tagged twigs. It has been observed that Shorea robusta takes three to four months to complete its reproductive cycle which comprises development of flower buds to the formation of fruits. The formation of flower buds starts in the month of March which converted into matured flowers in April. The young fruits were observed in the month of May. The fruits were upheld by tree for 30-45 days between May-June. The study reflects that in this species most of the growth phases have been occurred between March to June (Fig. 3a, b).
According to Table 1 in Ficus hispida the leaf bud busting initiated in the end of April after winter and all the young leaves become jubilant on the twigs in the month of July.
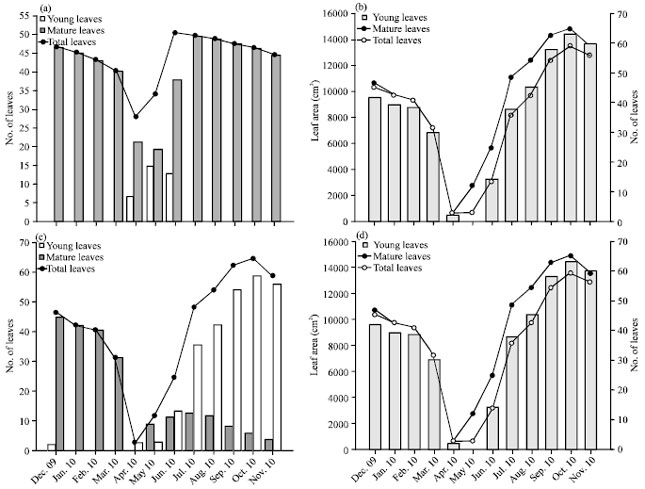 | |
| Fig. 3(a-d): | Shorea robusta Gaertn. f., (a) Variation of leaves number, (b) Variation of leaf area and Ficus hispida L. f., (c) Variation of leaves number and (d) Variation of leaf area |
| Table 1: | Different phenophases in Shorea robusta Gaertn. f. and Ficus hispida L. f. |
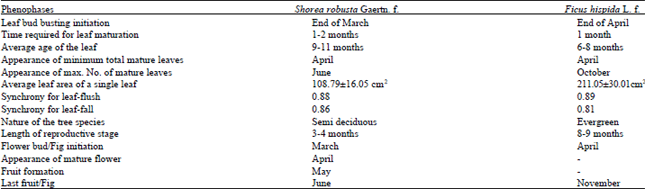 | |
The newly emerged leaves take about one month to get matured. The average age of the leaves in the species varies between 6-8 months. In the sanctuary area the Ficus hispida shows minimum number of total leaves in the month of April before the emergence of new leaf buds while the maximum in the month of October. As the average air temperature starts falling in the month of November, the leaf fall starts. The average area of single leaf was measured as 211.05±30.01 cm2. The average leaf area has been observed minimum in April while maximum in the month of October. The synchrony between selected tree specimens and whole population is 0.89 for leaf-flush and 0.81 for leaf-fall. Almost in every month except winter a few young leaves appear on the tagged twigs which show the evergreen nature of the species. After winter season, the formation of young figs starts in the month of April along with leaf bud initiation which continues till November. The young figs take up to two months for ripening. It shows that in this species the growth phase has dominates throughout the year except winter (Fig. 3c, d).
DISCUSSION
The leaf emergence and maturation in Shorea robusta were reported in pre monsoon period and it continues 3-4 months as also observed by Frankie et al. (1974), Whitmore (1984) and Singh and Singh (1992a). In Shorea robusta leaf flush initiates just after the winter and continue up to 3-4 months because it is a deep rooted tree which exploits the resources at slower rate and conserves them till the end of winter (Kushwaha and Singh, 2005). Ficus hispida also shows the leaf bud busting and leaf flushing in the same season but in the end of April as also observed by Frankie et al. (1974) and Whitmore (1984). The reason behind this emergence and maturation of the leaves in the dry season could be the increased day length, rise of temperature and change in photoperiod which favour to maximize the photosynthesis and vegetative growth (Sharma, 1970; Salisbury and Ross, 1974; Bhat, 1992; Bhat and Murali, 2001; Rivera et al., 2002; Hamann, 2004; Kushwaha and Singh, 2005; Dar and Malik, 2009). According to Borchert (1994), the increased photoperiod with rising temperature may cause conversion of starch into sugar in the roots and stem and osmotic adjustment in bud tissues which may induced bud busting by increasing water absorption and availability of sugars in summer flushing trees. In the present study leaf fall was observed maximum in the post monsoon season (Fig. 3a, c). Similar observation has also been reported by Sukwong et al. (1975) in dry Dipterocarp forest and Mishra et al. (2006) in moist deciduous forest. In the case of Shorea robusta, the tagged twigs drop a few leaves almost every month which reflect the semi-evergreen nature of the plants. Bor (1953) and Singh and Kushwaha (2005a) have also observed the semi-evergreen nature of the sal tree. Singh and Singh (1992b) reported that the initiation of leaf fall in post monsoon season may be a mechanism to maintain turgidity of shoot in cool and dry period. The high synchrony of leaf flush (0.88 for Shorea robusta and 0.89 for Ficus hispida) and leaf fall (0.86 for Shorea robusta and 0.81 for Ficus hispida) in both the species take place almost in the same period.
Both the species initiates flowering/figing in post winter season i.e. in the month of March (Shorea robusta) and April (Ficus hispida). The similar observations have also been reported by Mishra et al. (2006) and Bhat (1992) from the tree species of tropical moist deciduous forests of India. The flowering and figing are triggered by many abiotic factors (i.e. day length, irradiance, temperature, photoperiod etc.) associated with biotic factors (i.e. pollinators etc.) in the tropical trees (Alvim and Alvim, 1978; Putz, 1979; Hilty, 1980; Boulter et al., 2006). It was observed that in Shorea robusta the fruit setting starts in the month of May and get matured till June, just before the rains. The similar observation has been reported by Bhat (1992) and Mishra et al. (2006). The initiation of fruiting in dry season and maturation in late dry season, just before the rains provides opportunity for optimum seed germination and seedling establishment on moist soil (Frankie et al., 1974; Primack, 1987; Singh and Singh, 1992b; Singh and Kushwaha, 2005b, 2006; Kushwaha et al., 2011).
The study gives an idea about the time span of different life phases in the species. It clearly indicates that most of the phenological events in both the species are induced by photoperiod and temperature. This information may be used as base line for further evaluation of phenological variations in response of the climate change.
ACKNOWLEDGMENTS
The authors are grateful to Dr. C.S. Nautiyal, Director, CSIR - National Botanical Research Institute, Lucknow for providing facilities to carry out the study. The financial support was received from CSIR, New Delhi under NWP-020. The authors are also thankful to the forest staff of Katerniaghat Wildlife Sanctuary, Uttar Pradesh for granting permission and providing logistic support to conduct research work in the sanctuary area.
REFERENCES
- Appanah, S., 1985. General flowering in the climax rain forests of South-east Asia. J. Trop. Ecol., 1: 225-240.
CrossRef - Bhat, D.M., 1992. Phenology of tree species of tropical moist forest of Uttara Kannada district, Karnataka, India. J. Biosci., 17: 325-352.
CrossRefDirect Link - Bhat, D.M. and K.S. Murali, 2001. Phenology of understorey species of tropical moist forest of Western Ghats region of Uttara Kannada district in South India. Curr. Sci., 81: 799-805.
Direct Link - Borchert, R., 1994. Induction of rehydration and bud break by irrigation or rain in decidous trees of a tropical dry forest in Costa Rica. Trees, 8: 198-204.
CrossRef - Borchert, R., G. Rivera and W. Haynauer, 2002. Modification of vegetative phenology in a tropical semi-deciduous forest by abnormal drought and rain. Biotropica, 34: 27-39.
Direct Link - Boulter, S.L., R.L. Kitching and B.G. Howlett, 2006. Family, visitors and the weather: Patterns of flowering in tropical rain forests of northern Australia. J. Ecol., 94: 369-382.
CrossRef - Champion, H.G. and S.K. Seth, 1968. A Revised Survey of the Forest Types of India. 1st Edn., Govt. of India Press, Nueva Delhi, India, Pages: 404.
Direct Link - Coates-Estrada, R. and A. Estrada, 1986. Fruiting and frugivores at a strangler fig in the tropical rain forest of Los Tuxtlas, Maxico. J. Trop. Ecol., 2: 349-357.
CrossRef - Corlett, R.T. and J.V. Lafrankie, 1998. Potential impacts of climate change on tropical Asian forests through an influence on phenology. Clim. Change, 39: 439-453.
CrossRef - Devineau, J.L., 1999. Seasonal rhythms and phenological plasticity of savanna woody species in a fallow farming system (south-west Burkina Faso). J. Trop. Ecol., 15: 497-513.
CrossRef - Hamann, A., 2004. Flowering and fruiting phenology of a Philippine submontane rain forest: Climatic factors as proximate and ultimate causes. J. Ecol., 92: 24-31.
CrossRefDirect Link - Hilty, S.L., 1980. Flowering and fruiting periodicity in a premontane rain forest in pacific Columbia. Biotropica, 13: 292-306.
Direct Link - Kushwaha, C.P. and K.P. Singh, 2005. Diversity of leaf phenology in a tropical deciduous forest in India. J. Trop. Ecol., 21: 47-56.
CrossRef - Kushwaha, C.P., S.K. Tripathi and K.P. Singh, 2011. Tree specific traits affect flowering time in Indian dry tropical forest. Plant Ecol., 212: 985-998.
CrossRefDirect Link - Leigh Jr., E.G. and D.M. Windsor, 1982. Forest Production and Regulation of Primary Consumers on Barro Colorado Island. In: The Ecology of a Tropical Forest, Seasonal Rhythms and Long-Term Changes. Leigh Jr., E.G., A.S. Rand and D.M. Windsor (Eds.). Smithsonian Institution Press, Washington, DC USA., pp: 111-122.
- Mezel, A., 2002. Phenology: Its importance to the global change community. An editorial comment. Clim. Change, 54: 379-385.
CrossRef - Mishra, R.K., V.P. Upadhyay, S. Bal, P.K. Mohapatra and R.C. Mohanty, 2006. Phenology of species of moist deciduous forest sites of Similipal biosphere reserve. Lyonia, 11: 5-17.
Direct Link - Koul, M. and A.K. Bhatnagar, 2005. Phenology and climate change. Curr. Sci., 89: 243-244.
Direct Link - Prasad, S.N. and M. Hegde, 1986. Phenology and seasonality in the tropical deciduous forest of Bandipur, South India. Proc. Plant Sci., 96: 121-133.
Direct Link - Primack, R.B., 1987. Relationship among flowers, fruits and seeds. Ann Rev. Ecol. Sys., 18: 409-430.
CrossRef - Rivera, G., S. Elliott, L.S. Caldas, G. Nicolossi, V.T.R. Coradin, R. Borchert, 2002. Increasing day-length induces spring flushing of tropical dry forest trees in the absence of rain. Trees Struct. Funct., 16: 445-456.
Direct Link - Schwartz, M.D., 1999. Advancing to full bloom: Planning phenological research for the 21st century. Int. J. Biometeorol., 42: 113-118.
CrossRef - Schwartz, M.D., R. Ahas and A. Aasa, 2006. Onset of spring starting earlier across the northern hemisphere. Global Change Biol., 12: 343-351.
CrossRef - Singh, J.S. and V.K. Singh, 1992. Phenology of seasonally dry tropical forest. Curr. Sci., 63: 684-689.
Direct Link - Singh, K.P. and C.P. Kushwaha, 2005. Paradox of leaf phenology: Shorea robusta is a semi-evergreen species in tropical dry deciduous forests in India. Curr. Sci., 88: 1820-1824.
Direct Link - Singh, K.P. and C.P. Kushwaha, 2005. Emerging paradigms of tree phenology in dry tropics. Curr. Sci., 89: 964-975.
Direct Link - Singh, K.P. and C.P. Kushwaha, 2006. Diversity of flowering and fruiting phenology of trees in a tropical deciduous forest in India. Ann. Bot., 97: 265-276.
CrossRef - Yadav, R.K. and A.S. Yadav, 2008. Phenology of selected woody species in a tropical dry deciduous forest in Rajasthan, India. Trop. Ecol., 49: 25-34.
Direct Link - Zhang, G.M., Q.S. Song and D.R. Yang, 2006. Phenology of ficus racemosa in Xishuangbanna, Southwest China. Biotropica, 38: 334-341.
CrossRef - Dar, M.E.U.I. and Z.H. Malik, 2009. A floristic list and phenology of plant species of Lawat Area District Neelum, Azad Jammu and Kashmir, Pakistan. Int. J. Bot., 5: 194-199.
CrossRefDirect Link