
Fahima Gul
Department of Botany, University of Kashmir, Srinagar, 190006, India
Inayatullah Tahir
Department of Botany, University of Kashmir, Srinagar, 190006, India
Inam Ul Rasool
Department of Soils, Punjab Agriculture University, Ludhiana Punjab, 140001, India
International Journal of Botany
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 22-30







ABSTRACT
A study was conducted to determine the effects of pretreatment at varying concentrations of Cycloheximide (CHI) on senescence and postharvest performance in Nerine sarniensis. At a particular threshold concentration CHI delays senescence and above which it prevents flower opening and promotes senescence. The fact that cycloheximide delays tepal senescence demonstrates the synthesis of particular proteins probably enzymes, responsible for degradation of cellular constituents, executes the cell death programme in flower tepals. Pretreatment of scapes with CHI at 0.01 and 0.05 mM concentrations was found to delay visible signs of senescence, maintain a sustained rate of flower blooms and increase fresh and dry mass, besides lowering electrical conductivity of ion leachates and solution absorption. An increase in total sugar, phenolic and soluble protein content was observed with a concomitant decrease in α-amino acid content. Pretreatment of scapes with 0.05 or 0.01 CHI for 1 h can be used as an effective treatment to improve the post harvest longevity in this flower system.
PDF Abstract XML References Citation
Received: August 23, 2011;
Accepted: November 07, 2011;
Published: January 23, 2012
How to cite this article
Fahima Gul, Inayatullah Tahir and Inam Ul Rasool, 2012. Senescence and Postharvest Performance of Cut Nerine sarniensis Flowers: Effects of Cycloheximide. International Journal of Botany, 8: 22-30.
DOI: 10.3923/ijb.2012.22.30
URL: https://scialert.net/abstract/?doi=ijb.2012.22.30
DOI: 10.3923/ijb.2012.22.30
URL: https://scialert.net/abstract/?doi=ijb.2012.22.30
INTRODUCTION
Tepal senescence is a morphologically and metabolically an active process regulated strictly at both transcriptional and translational levels (Lawton et al., 1990; Nooden et al., 1997; Hoeberichts et al., 2005). Proteins or proteolytic cleavage is thought to play a significant role in the senescence of flowers as the expression of protease genes is one of the earliest senescence related gene change to be identified in various ornamental cut flowers (Eason et al., 2002; Wagstaff et al., 2002; Jones et al., 2005; Van Doorn and Woltering, 2008). The interactions between proteases and their inhibitors have been linked to modulation of cell death processes in plants and in certain cut flowers. Chemical inhibition of protease action modulates the onset of cellular senescence thereby developing a new strategy of delaying flower senescence and improving the postharvest performance in ornamental flowers (Eason et al., 2002; Sin and Chy, 2004; Pak and Van Doorn, 2005).
Cycloheximide (a protein synthesis inhibitor at the translational level) has been implicated to effectively delay senescence in flowers such as Consolida, Dianthus, Gladiolus, Hemerocallis, Ipomea, Iris, Narcissus (Wulster et al., 1982; Lukaszewski and Reid, 1989; Courtney et al., 1994; Jones et al., 1994; Celikel and Van Doorn, 1995; Gulzar et al., 2005; Shahri and Tahir, 2010). Cycloheximide has been found to improve the water balance of cut flowers such as Iris and Dianthus (Van Doorn et al., 1995; Drory et al., 1995). The role of cycloheximide in delaying or accelerating tepal senescence at various concentrations points to its regulatory effects on the possible senescence related proteins synthesized de novo which may also interfere with transient water stress in cut flowers (Drory et al., 1995).
Only a little information is available on the improvements of Nerine sarniensis as a cut flower crop (Downs and Reihana, 1987; Berghoef and Van Brenk, 1982; Lukaszewski, 1997; Vishnevetsky et al., 1997; Gul and Tahir, 2009). Nerines are native to South Africa and were first described as Narcissus japonicus rutilo flore in 1635 by J. Cornut. Later in 1735 Linnaeus included this species under the name Amaryllis sarniensis and in 1820 the genus Nerine was established by Herbert (Zonneveld and Duncan, 2006). Nerine flowers from early September to early October in Kashmir. The plants possess glorious cluster of red blooms on each sturdy scape and are obvious choice for cut flower production. The present study was undertaken to investigate the effects of pretreatment with different concentrations of Cycloheximide (CHI) before transfer to Distilled Water (DW) or Sucrose (0.15 M) on senescence with the ultimate aim to gain an insight into the mechanism of senescence and to improve its postharvest performance.
MATERIALS AND METHODS
Plant material: Bulbs of N. sarniensis were planted in Kashmir University Botanical Garden in November 2009. The bulbs were allowed to grow under complete natural conditions. In October 2010 the emergent scapes of N. sarniensis cv. Red were used for the present study (Fig. 1). The scapes were harvested at 0800 h when the spathe had broken loose and buds were easily countable; stage III, loose bud stage (Fig. 2). The harvested scapes were immediately brought to the laboratory, cut to a uniform size of 30 cm and pulse treated for 1 h separately with varying concentrations of cycloheximide (0.01, 0.05, 0.1 and 0.5 mM). After pulse treatment the scape ends were washed thrice with distilled water. In each case two scapes were transferred to 250 mL Borosil conical flasks containing 200 mL of distilled water (Set A) or sucrose (Set B). A separate set of five flasks each containing unpulsed scapes represented controls (D.W) and sucrose (0.15 M). Overall there were ten treatments including control and sucrose. Treatment effects were evaluated by keeping the flowers in the laboratory at a temperature of 25±2°C under cool white fluorescent light with a mix of diffused natural light (10 W m-2) 12 h a day and RH of 60±10%. Treatment was given on the day of harvest which was designated as day zero.
Assessment of vase life, blooming and solution uptake: The average vase life of scapes was counted from the day zero and was arbitrarily assessed when the flowers showed reduced display value and had attained “S4” status which was designated as “less attractive”. Senescence was characterized by curling of tepals, turgor loss followed by wilting of tepals (Table 1). Number of blooms per scape was recorded till maximum numbers of buds bloomed in a particular treatment including the controls. The volume of holding solution absorbed by the scapes was calculated by measuring the volume of solution on a particular day and subtracting it from the initial quantity of the vase solution kept in the flasks. To account for the volume of solution evaporated blank flasks (containing particular vase solutions without scapes) were used in triplicates alongside the flasks with scapes. Volume of holding solution absorbed per scape was recorded on every second day of the experiment till the controls senesced.
 | |
| Fig. 1: | Plants of Nerine sarniensis cv. Red in full bloom |
 | |
| Fig. 2: | Stages of scape development in Nerine sarniensis cv. Red. Stage III “looses bud stage” was used for the present study |
| Table 1: | Assessment of vase life Nerine sarniensis cv. Red by assigning a particular status based on the display values |
 | |
Conductivity of leachates, fresh and dry mass: Conductivity of leachates, fresh and dry mass of the flowers was determined on 4th and 8th day of harvest. Dry mass was determined by drying the material in an oven for 48 h at 70°C. Membrane permeability was studied by measuring ion leakage from the tepal discs (5 mm in diameter) incubated in the dark in 15 mL glass double distilled water for 15 h at 20°C. The discs were punched from the flag region of tepals of 5 flowers. The discs were floated with their abaxial surface downwards and were removed after 15 h of incubation. Conductivity of leachates was measured by CM-180 ELICO Conductivity meter and was expressed in μS.
Tissue constituents: At each stage 0.5 g chopped material of tepal tissue was fixed in triplicate in hot 80% ethanol. The material was macerated and centrifuged three times. The supernatants were pooled and used for the estimation of sugars and total phenols. Reducing sugars were estimated by the method of Nelson (1944) using glucose as the standard. Total sugars were estimated after enzymatic conversion of non-reducing sugars into reducing sugars with invertase. Non-reducing sugars were calculated as the difference between total and reducing sugars. α-amino acids were estimated by the method of Rosen (1957) using Glycine as standard. Total phenols were estimated by the method of Swain and Hillis (1959) using Gallic acid as standard. Tissue constituents were estimated on day 4 and 8 of the treatment taking into consideration the already senescent status of flowers from controls i.e. DW or SUC (0.15 M) whose vase life was terminated by day 4 or 5 of the treatment, respectively.
Soluble proteins: Proteins were extracted from 0.5 g perianth tissue drawn separately from different flowers. The tissue was homogenized in 5 mL of 5% sodium sulphite (w/v) adding 0.1 g of polyvinylpyrrolidone and centrifuged. Proteins were precipitated from a suitable volume of cleared supernatant with equal volume of 20% trichloroacetic acid, centrifuged at 1000xg for 15 min and the pellet redissolved in 0.1 N NaOH. Proteins were estimated from a suitable aliquot by the method of Lowry et al. (1951) using BSA as the standard. Soluble proteins were estimated on day 4 and 8 of the treatment taking into consideration the already senescent status of flowers from controls i.e. DW or SUC (0.15M) whose vase life was terminated by day 4 or 5 of the treatment, respectively.
Statistical analysis: Each value represents the mean of five independent replicates. Differences between the treatments have been calculated by simple analysis of variance and Least Significant Difference (LSD) computed at P0.05 using Minitab version 11.
RESULTS
Vase life, blooming and solution uptake: The average life of an individual flower under field conditions after it had opened fully was about 3 days. This was followed by the loss of turgor and appearance of dull whitish patches on the curvy tepals. Finally, the tepals and stamens wilt and wither. The scape finally dislodges leaving a bunch of leaves behind. The average vase life of scapes harvested at loose bud stage was about 4-5 days in water or sucrose. The vase life of scapes pulse treated with 0.01 and 0.05 mM CHI before transfer to distilled water or sucrose was 10-11 days; pulse treatment with higher concentrations of CHI (0.1 and 0.5 mM) registered a decrease in vase life as compared to distilled water or sucrose (Fig. 3a).
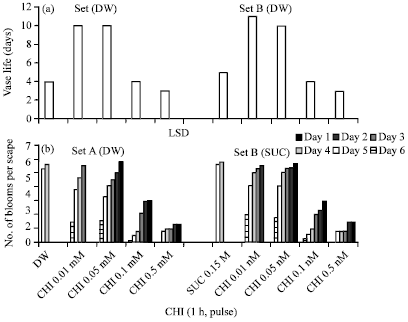 | |
| Fig. 3(a-b): | Histograms showing effect of pulse treatment with cycloheximide (1 h) (a) on vase life and (b) blooming in cut scapes of Nerine sarniensis cv. Red |
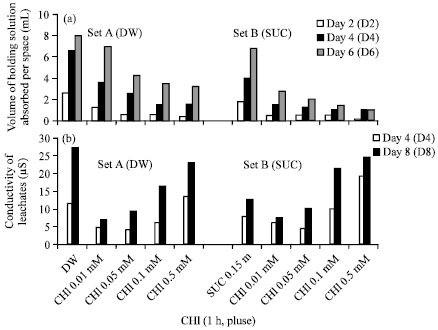 | |
| Fig. 4(a-b): | Histograms showing effect of pulse treatment with cycloheximide (1 h) on (a) solution uptake and (b) conductivity of leachates in cut scapes of Nerine sarniensis cv. Red |
Pulse treatment of scapes with CHI (0.01 and 0.05 mM) before transfer to distilled water or sucrose resulted in sustained rate of blooming as compared to untreated scapes held either in distilled water or sucrose which achieved 100% blooming on just second day after treatment; the opening of flowers was markedly prevented by 0.1 and 0.5 mM CHI treatment (Fig. 3b). Pulse treatment of scapes with various concentrations of CHI resulted in corresponding decrease in the volume of holding solution absorbed, the decrease was highly pronounced when the scapes were transferred to sucrose particularly after pulse treatment with 0.1 and 0.5 mM CHI. The increase in the volume of holding solution absorbed with the progression in time (days after harvest) was pronounced in the untreated scapes held in either distilled water or sucrose (Fig. 4a).
Membrane permeability, fresh and dry mass: The ion leakage of the tepal discs increased with the progression in time up to 8th day after harvest in all the treatments, however the rise in the ion leakage was minimal in samples from scapes pretreated with 0.01 and 0.05 mM CHI indicating that at these concentrations membrane integrity was maintained in the tepals. Pretreatment of scapes with CHI (0.01 and 0.05 mM) before transfer to either distilled water or sucrose resulted in considerable decrease in the ion leachates as compared to the tepal discs of flowers from untreated scapes held either in distilled water or sucrose; however a sharp increase in the leachates was registered in samples from the scapes pretreated with 0.1 and 0.5 mM CHI before transfer to either distilled water or sucrose (Fig. 4b).
Generally pulse treatment of scapes with 0.01 mM CHI resulted in an increase in fresh and dry mass of flowers which was sustained up to 8 days after harvest as compared to untreated scapes held particularly in distilled water. Decrease in fresh and dry mass of flowers was generally recorded in samples from scapes pretreated with CHI at 0.05, 0.1 and 0.5 mM concentrations; however, fresh and dry mass of flowers was sustained with progression in time from 4 to 8 days after harvest (Fig. 5a, b).
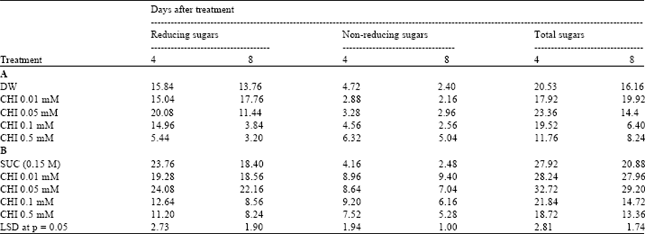
Tissue constituents (sugars, α-amino acids and phenols): The content of sugars decreased with the progression in age from 4 to 8 days after harvest. The reducing sugar content was maintained with reference to controls in the tepal tissue samples from scapes pulse treated with CHI at 0.01 and 0.05 mM concentrations followed by transfer to either distilled water or sucrose; however a sharp decrease in the content of reducing sugars was recorded in tepal tissue samples from scapes pulse treated with 0.1 and 0.5 mM CHI. The non-reducing sugar content generally registered an increase by treatment with various grades of CHI; however, the increase was sharp in pretreated scapes transferred to sucrose. The content of total sugars was, however, maintained in tepal tissue samples from scapes pulse treated with CHI from 0.01 to 0.05 mM concentrations (Table 2).
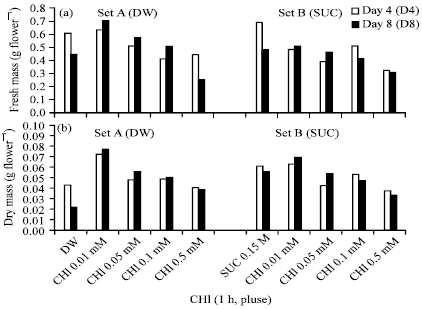 | |
| Fig. 5(a-b): | Histograms showing effect of pulse treatment with cycloheximide (1h) on (a) fresh mass g flower-1 and (b) dry mass g flower-1 in cut scapes of Nerine sarniensis cv. Red |
| Table 2: | Effect of pulse treatment with varying concentrations of cycloheximide (CHI, 1h pulse) before transfer to DW (A) or SUC (B) on sugar fractions in tepal tissues on day 4 and 8 after transfer, of cut scapes to holding solutions in Nerine Sarniensis cv. Red as expressed on fresh mass basis (mg g FM) |
 | |
| Each value is a mean of 5 independent replicates. Room temperature RT = (21±2°C) | |
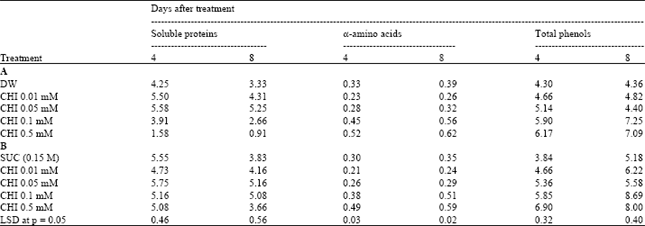
The total phenolic content registered an increase with progression in age from 4 to 8 days of harvest. Pretreatment with CHI generally resulted in an increase in the content of total phenols irrespective of the transfer of scapes to either distilled water or sucrose. The soluble protein content decreased with the progression in age from 4 to 8 days after harvest irrespective of treatment of scapes. However, the decrease in soluble protein content was minimal in tepal tissues from scapes pulse treated with 0.01 and 0.05 mM CHI. Indicating maintenance of proteins at these concentrations. The decrease in the soluble protein content was pronounced in tepal tissues from scapes pulse treated with 0.1 and 0.5 mM (Table 3).
α-amino acids followed a reverse trend as that of soluble proteins. The amino acids content registered an increase with the progression in age from day 4 to day 8 of treatment of scapes. However, the amino acids content was generally maintained over a period of time in tepal tissue samples from scapes pulse treated with 0.01 and 0.05 mM CHI. The amino acid content registered a sharp increase in tepal tissue samples from scapes pretreated with higher concentrations of CHI (0.1 and 0.5 mM) as compared to untreated controls both in DW or SUC indicating enhanced protein degradation (Table 3).
| Table 3: | Effect of pulse treatment with varying concentrations of cycloheximide (CHI, 1h pulse) before transfer to DW (A) or SUC (B) on soluble proteins α-amino acids and total phenols in tepal tissues on day 4 and 8 after transfer of cut scapes to holding solutions in Nerine sarniensis cv. Red as expressed on fresh mass basis (mg g-1 FM) |
 | |
| Each value is a mean of 5 independent replicates. Room temperature RT = (21±2°C) | |
DISCUSSION
The results of our experiments suggest that pulse treatment of scapes at a particular threshold level of CHI (0.05 mM) followed by transfer to distilled water or sucrose (0.15 M) enhanced vase life by an increment of about 6 days besides resulted in sustained rate of blooming. However, at higher concentrations of CHI (0.1 and 0.5 mM) the flowers wilted quickly and senesced before controls besides; the opening of flowers was also prevented. With increasing concentrations of cycloheximide senescence was accelerated and flower blooming was retarded (Fig. 6). Cycloheximide has been found to delay senescence and inhibit flower opening in Dianthus, Hemerocallis, Gladiolus and Iris (Wulster et al., 1982; Lukaszewski and Reid, 1989; Jones et al., 1994; Van Doorn et al., 1995; Celikel and Van Doorn, 1995; Gulzar et al., 2005; Zhuo et al., 2005). Present results suggest that at levels below 0.05 mM cycloheximide regulated senescence and maintained a sustained rate of flower bloom.
The effect of cycloheximide in delaying the senescence does not seem to be due to improvement of water balance of cut Nerine scapes as pulse treatment of scapes with various concentrations of CHI resulted in corresponding decrease in the volume of holding solution absorbed. Cycloheximide has been shown to rapidly reduce the rate of transpiration and water uptake in Iris tepals which apparently was not suggested to be the reason for the delay in tepal wilting (Van Doorn et al., 1995).
The present study suggests that pulse treatment of scapes with CHI (0.01 and 0.05 mM) resulted in considerable decrease in the ion leachates of tepal tissue samples. The loss of membrane integrity has been shown to cause an increase in the permeability and leakage during senescence in various flowers such as Arum, Ipomoea, Dianthus, Iris, Hemerocallis, Rosa and Gerbera (Van Meeteren, 1979; Halevy and Mayak, 1979; Lay-Yee et al., 1992; Celikel and Van Doorn, 1995; Gulzar et al., 2005). The delay in leakiness of tepal cells due to the application of cycloheximide indicates that one or more proteins synthesized de novo are responsible for the increase in leakiness; the protein that is rate limiting in the senescence programme could either be a factor that regulates gene expression or an enzyme (Celikel and Van Doorn, 1995). It is interesting to note that understanding the cause of membrane breakdown in senescing tissues has implications for signal transduction chains, as the components of these chains are often associated with the membranes. Use of potent protein synthesis inhibitors like CHI in maintaining the membrane integrity could modulate such cascades. Such studies offer the scope to identify the components of the signal transduction chain resulting in PCD. Maintenance of higher fresh and dry mass of flowers particularly at lower concentration of CHI (0.01 mM) could be due to lower respiratory losses as CHI has been found to suppress respiration in certain plant tissue; besides in Hemerocallis it has been shown to abolish the peak in respiration at the start of senescence (Ellis and Macdonald, 1970; Bieleski and Reid, 1992).
During the current investigation it has been shown that the content of sugars generally decreased with progression in age from 4 to 8 days after harvest, however the content of total sugars was maintained in samples from scapes pulse treated with CHI (0.01 and 0.05 mM) concentrations. Flower maturation and senescence has been shown to be accompanied by a decline in total carbohydrate content in flowers such as Carnations and it has been reported that there is a small but consistent respiratory climacteric coinciding with the early senescence in Hemerocallis flowers (Nichols, 1973; Paulin and Jamain, 1982; Lukaszewski and Reid, 1989; Lay-Yee et al., 1992; Gulzar et al., 2005).
 | |
| Fig. 6(a-d): | Effect of pretreatment with varying grades of cycloheximide (CHI 1h pulse) and subsequent transfer to distilled water (D.W) or sucrose 0.15 M (SUC) on scapes of Nerine sarniensis cv. Red scapes. From left to right are arranged scapes held in D.W (control), SUC, CHI (0.01 mM→DW), CHI (0.01 mM→SUC), CHI (0.05 mM→DW), CHI (0.05 mM→SUC), CHI (0.1 mM→DW), CHI (0.1 mM→SUC), CHI (0.5 m→M DW) and CHI (0.5 mM→SUC). Various figures represent scapes (a) 3 days after harvest, (b) 6 days after harvest, (c) 9 days after harvest and (d) 12 days after harvest |
The maintenance of total sugars in CHI treated samples during the current investigation suggests that delay in senescence is caused by curtailment of respiratory climacteric otherwise in normal senescence process sugars are rapidly respired and utilized (Mwangi et al., 2003; Gulzar et al., 2005).
The soluble protein content decreased whereas, α-amino acid and total phenolic content increased with progression in age from 4 to 8 days of harvest. The higher content of total phenols has been shown to be associated with longer vase life in cut rose petals and Hemerocallis (Van Doorn and Stead, 1994; Mwangi et al., 2003; Gulzar et al., 2005). In Day lily tepals, a sharp decrease in protein levels preceded the visible symptoms of senescence and cycloheximide delayed the decrease in protein levels and increased the time to visible senescence (Lay-Yee et al., 1992; Courtney et al., 1994). This suggests that protein degradation or increased ubiqutination may lead to senescence. Cycloheximide treated flowers of Day lily apart from exhibiting a delay in senescence showed a curtailment of the decline in the protein content and protein population (Lay-Yee et al., 1992). The current investigation suggests that at threshold level of 0.05 mM CHI maintained the protein population of the tepals and delayed senescence by 6 days. It may be either due to blocking of ubiqutin proteins or over all reduction in protein synthesis or by inhibiting the synthesis of specific proteases responsible for protein degradation. Conversely at higher concentrations CHI promoted senescence by increasing tepal wilting; however in order to establish a relationship between increase in cell leakiness and protein degradation, protein levels of tonoplast or plasma membrane need to be assessed.
Regulating the senescence associated activity of proteases may be achieved with different molecular strategies e.g., the accumulation of cysteine protease mRNA’s in senescing carnation flowers is associated with a decrease in protease inhibitor mRNA (Sugawara et al., 2002), indicating that inhibitor proteins may play a role in senescence associated protease activity in flowers. However proteases have been found to be localized to plant vacuoles and both post translational modification and sub cellular localization provide the cell with the means to regulate protease activity. The modification of proteolysis in the down regulation of senescence associated cysteine proteases offers a strategy to produce transgenic plants as has been achieved in broccoli (Eason et al., 2005). Even the active compound in the mucilage of daffodil flowers (Narciclasine) which delays tepal senescence in cut Iris flowers is known to inhibit protein synthesis (Van Doorn et al., 2004). So far present studies on a range of protein synthesis inhibitors such as Phenyl Methyl Sulfonyl Fluoride (PMSF), Amikacin Sulphate (AMS) and Cycloheximide (CHI) etc., have shown that cycloheximide is a versatile candidate for studies on inhibition of protein synthesis.
The present investigation suggests that at a particular threshold level CHI delays senescence and beyond it CHI prevents flower opening and promotes senescence. The present results suggest that the effects of cycloheximide indicate a programme at the cellular level. The fact that cycloheximide delays tepal senescence demonstrates that the synthesis of some particular proteins orchestrates the cell death programme, however, it is necessary to show that these proteins and their products actually play a causal role. Isolation of these proteins could provide an important tool for studying senescence and in future manipulating it. Though cycloheximide may have some side effects but a pulse treatment below 0.05 mM marks it as an important means f increasing the commercial value of ornamental plants like Nerine. Scapes of Nerine sarniensis pulse treated with CHI (0.01 and 0.05 mM) resulted in marked improvement in vase life, blooming and maintenance of membrane integrity, besides optimising its postharvest performance.
REFERENCES
- Berghoef, J. and G. van Brenk, 1982. Effect of temperature on flowering of Nerine bowdenii W. Watts. Acta Hort., 134: 41-48.
Direct Link - Bieleski, R.L. and M.S. Reid, 1992. Physiological changes accompanying senescence in the ephemeral daylily flower. Plant Physiol., 98: 1042-1049.
Direct Link - Celikel, F.G. and W.G. van Doorn, 1995. Solute leakage, lipid peroxidation and protein degradation during the senescence of Iris tepals. Plant Physiol., 94: 515-521.
Direct Link - Courtney, S.E., C.C. Rider and A.D. Stead, 1994. Changes in protein ubiquitination and the expression of ubiquitin-encoding transcripts in daylily petals during floral development and senescence. Physiol. Plant., 91: 196-204.
CrossRefDirect Link - Downs, C. and M. Reihana, 1987. Extending vase life and improving quality of Nerine cut flowers with preservatives. Hort. Sci., 22: 670-671.
Direct Link - Drory, A., S. Beja-Tal, A. Borochov, E. Gindin and S. Mayak, 1995. Transient water stress in cut carnation flowers: Effects of cycloheximide. Sci. Hort., 64: 167-175.
Direct Link - Eason, J.R., D.J. Ryan, T.T. Pinkney and E.M. O'Donoghue, 2002. Programmed cell death during flower senescence: Isolation and characterization of cysteine proteinases from Sandersonia aurantiaca. Funct. Plant Biol., 29: 1055-1064.
Direct Link - Eason, J.R., D.J. Ryan, L.M. Watson, D. Hedderley, M.C. Christey, R.H. Braun and S.A. Coupe, 2005. Suppression of the cysteine protease, aleurain, delays floret senescence in Brassica oleraceae. Plant Mol. Biol., 57: 645-657.
CrossRef - Ellis, R.J. and I.R. MacDonald, 1970. Specificity of cycloheximide in higher plant systems. Plant. Physiol., 46: 227-232.
PubMedDirect Link - Gul, F. and I. Tahir, 2009. Effect of cool and wet storage on postharvest performance of Nerine sarniensis cv. red scapes. Acta Hort., 847: 345-352.
Direct Link - Gulzar, S., I. Tahir, I. Amin, S. Farooq and S.M. Sultan, 2005. Effect of cytokinins on the senescence and longevity of isolated flowers of daylily (Hemerocallis fulva) cv. Royal crown sprayed with cycloheximide. Acta. Horticult., 669: 395-403.
Direct Link - Halevy, A.H. and S. Mayak, 1979. Senescence and postharvest physiology of cut flowers, Part 1. Hortic. Rev., 1: 204-236.
Direct Link - Jones, R.B., M. Serek, C.L. Kuo and M.S. Reid, 1994. The effect of protein synthesis inhibition on petal senescence in cut bulb flowers. J. Am. Soc. Hort. Sci., 119: 1243-1247.
Direct Link - Jones, M.L., G.S. Chaffin, J.R. Eason and D.G. Clark, 2005. Ethylene sensitivity regulates proteolytic activity and cysteine protease gene expression in petunia corollas. J. Exp. Bot., 56: 2733-2744.
CrossRefDirect Link - Lawton, K.A., K.G. Raghothama, P.B. Goldsbrough and W.R. Woodson, 1990. Regulation of senescence-related gene expression in carnation flower petals by ethylene 1. Plant Physiol., 93: 1370-1375.
Direct Link - Lay-Yee, M., A.D. Stead and M.S. Reid, 1992. Flower senescence in daylily (Hemerocallis). Physiol. Plant., 86: 308-314.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Lukaszewski, A.J., 1997. Improving keeping qualities of Nerine cut flowers with preservatives. Acta Hort, 430: 439-446.
Direct Link - Lukaszewski, T.A. and M.S. Reid, 1989. Bulb type flower senescence. Acta. Horticult., 261: 59-62.
Direct Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
Direct Link - Nooden, L.D., J.J. Guiamet and I. John, 1997. Senescence mechanisms. Physiol. Plant, 101: 746-753.
CrossRefDirect Link - Pak, C. and WG. van Doorn, 2005. Delay of Iris flower senescence by protease inhibitors. New Phytol., 165: 473-480.
PubMed - Paulin, A. and C. Jamain, 1982. Development of flowers and changes in various sugars during opening of cut carnations (Dianthus caryophyllus). J. Am. Soc. Horticult. Sci., 107: 258-261.
Direct Link - Rosen, H., 1957. A modified ninhydrin colorimetric analysis for amino acids. Arch. Biochem. Biophys., 67: 10-15.
CrossRefPubMedDirect Link - Shahri, W. and I. Tahir, 2010. Effect of cycloheximide on senescence and postharvest performance of Ranunculus asiaticus flowers. Pak. J. Bot., 42: 3577-3585.
Direct Link - Sin, S.F. and M.L. Chy, 2004. Expression of proteinase inhibitor II proteins during floral development in Solanum americanum. Planta, 219: 1010-1022.
CrossRefDirect Link - Sugawara, H., K. Shibuya, T. Yoshioka, T. Hashiba and S. Satoh, 2002. Is a cysteine proteinase inhibitor involved in the regulation of petal wilting in senescing carnation (Dianthus caryophyllus L.) flowers? J. Exp. Bot., 53: 407-413.
Direct Link - Swain, T. and W.E. Hillis, 1959. The phenolic constituents of Prunus domestica. I. The quantitative analysis of phenolic constituents. J. Sci. Food Agric., 10: 63-68.
CrossRefDirect Link - Van Doorn, W.G., A. Sinz and M.M. Thompson, 2004. Daffodil flowers delay senescence in cut Iris flowers. Phytochemistry, 65: 571-577.
CrossRef - Van Doorn, W.G. and E.J. Woltering, 2008. Physiology and molecular biology of petal senescence. J. Exp. Bot., 59: 453-480.
CrossRefDirect Link - Van Meeteren, U., 1979. Water relations and keeping quality of cut gerbera flowers. III. Water content, permeability and dry weight of aging petals. Sci. Horticult., 10: 261-269.
CrossRef - Vishnevetsky, J., N. Azizbekova, M. Ziv and H. Lilien-Kipnis, 1997. Development of the bulb and inflorescence in outdoor grown Nerine sarniensis. Acta Hort., 430: 147-157.
Direct Link - Wagstaff, C., M.K. Leverentz, G. Griffiths, B. Thomas, U. Chanasut, A.D. Stead and H.J. Rogers, 2002. Cysteine protease gene expression and proteolytic activity during senescence of Alstroemeria petals. J. Exp. Bot., 53: 233-240.
Direct Link - Zonneveld, B.J.M. and G.D. Duncan, 2006. Genome size for the species of Nerine herb. (Amaryllidaceae) and its evident correlation with growth cycle, leaf width and other morphological characters. Plant Syst. Evol., 257: 251-260.
CrossRefDirect Link - Hoeberichts, F.A., A.J. de Jong and E.J. Woltering, 2005. Apoptotic-like cell death marks the early stages of gypsophila (Gypsophila paniculata) petal senescence. Postharvest Biol. Technol., 35: 229-236.
CrossRef