
Saikat Gantait
Department of Biotechnology, Instrumentation and Environmental Science, Bidhan Chandra Krishi Viswavidyalaya, Mohanpur, W.B. 741252, India
LiveDNA: 91.241
ORCID: 0000-0001-5059-2428
Nirmal Mandal
Department of Biotechnology, Instrumentation and Environmental Science, Bidhan Chandra Krishi Viswavidyalaya, Mohanpur, W.B. 741252, India
International Journal of Botany
Year: 2010 | Volume: 6 | Issue: 3 | Page No.: 207-219







ABSTRACT
Anthurium, with over 108 described genera and 3750 species, is one of the most popular tropicals, highly appreciated for its brightly coloured and long lasting flowers. The exotic Anthurium industry plays a significant role in the global floriculture trade. To overcome the demerits of conventional propagation of Anthurium, plant tissue culture proved to be an influential tool that can complement conventional breeding and accelerate Anthurium development. This review presents a consolidated explanation of in vitro propagation and focuses upon contemporary information on biotechnological advances made in Anthurium.
PDF Abstract XML References Citation
Received: May 27, 2010;
Accepted: June 09, 2010;
Published: September 02, 2010
How to cite this article
Saikat Gantait and Nirmal Mandal, 2010. Tissue Culture of Anthurium andreanum: A Significant Review and Future Prospective. International Journal of Botany, 6: 207-219.
DOI: 10.3923/ijb.2010.207.219
URL: https://scialert.net/abstract/?doi=ijb.2010.207.219
DOI: 10.3923/ijb.2010.207.219
URL: https://scialert.net/abstract/?doi=ijb.2010.207.219
INTRODUCTION
Floriculture puts its best foot forward to shift a considerable amount of capital in productive chain every year. Anthurium, for its exquisiteness, durability and by the terms of long vase-life stands out among most of the tropical cultivated flowers. As a member of the Araceae family, Anthurium, the native to the tropics of Central and South America (Gantait et al., 2008), consists of 108 genera and approximately 3750 monocotyledonous species (both herbaceous and creepers) (Viegas et al., 2007). Principally, the arums are a tropical family, occurring naturally on all continents, with an exception of Antarctica. The genus having highly ornamental inflorescence and/or foliage are Anthurium, Caladium, Dieffenbachia, Monstera, Philidendron and Spathyphyllum (Gryum, 1990). A vast number of Anthurium species are duly produced and traded internationally as cut-flowers, flowering potted plants and landscape plants among which most of the cut-flower Anthuriums are believed to be hybrids of Anthurium andraeanum Linden ex André with several closely related species in the section Calomystrium (Croat and Sheffer, 1983) and have been referred to as Anthurium andraeanum Hort. (Kamemoto and Kuehnle, 1996). According to the report of Kuehnle et al. (1992), the breeding of two species, A. andreanum Linden ex André and A. scherzerianum Schott., has resulted in many cultivars which are used as potted plants as well as cut flowers, grown commercially all over the world. Until now, the identification of the cultivars has mainly been based on the spathe colour (Kobayashi et al., 1987). A large number of Anthurium cultivars displaying an array of spathe colours (ranging from red, orange, pink, coral and white) and belonging to the three spathe categories- namely, standard (single-coloured heart-shaped), obaki (bicolours of green with another major anthocyanin colour) and tulip-type are cultivated in order to meet the demand of the various market preferences in terms of colours, shades, floral sizes and shapes. The production of a single flower from each leaf axil takes place during the sympodial phase. In fact, the commercial flower is an inflorescence, composed of the spadix (a spike of minute flowers closely arranged round a fleshy axis) and the spathe (a large sheathing bract enclosing the flower cluster) (Higaki et al., 1984). The Anthurium gains its fame and respected status by exhibiting its striking ensemble, which is created unitedly by its spadix and its spathe, within the economically essential ornamentals, allowing its use in interior and exterior decoration and also to its use as a cut flower (Lopes and Mantovani, 1980; Castro et al., 1986). In the worldwide market, the Anthuriums are flowers with intense importance. The commercial value of Orchids and Anthuriums among the tropical cut flowers are first and second, respectively (Dofour and Guerin, 2003; Chen et al., 2003). Due to the increasing demand for cut flowers in the global market, large numbers of novel Anthurium cultivars are continually being imported from the Netherlands and Hawaii for commercial cultivation (Nowbuth et al., 2005).
PROBLEMS ASSOCIATED WITH CONVENTIONAL PROPAGATION
Propagation of Anthurium can be possible by both asexual and sexual methods. Conventional vegetative or asexual propagation of Anthurium is performed through terminal and single node cuttings along with the division of newly emerging suckers or offshoots, which can also be induced by topping (Mahanta and Paswan, 2001). In spite of producing homogenious materials, vegetative propagation of Anthurium is a very measured process, taking it years to develop commercial quantities of elite clones (De Lima et al., 2006) and thus not proving very advantageous (Pierik et al., 1974; Higaki and Ramussen, 1979; Geier, 1990). The traditional propagation of hybrid is challenging due to the growth of a low number of new plants from the base (Martin et al., 2003). In other way Anthurium is sexually propagated by seed (Dofour and Guerin, 2003), although, some taxa fail to produce viable seeds due to incompatibilities (Sheffer and Kamemoto, 1976). Moreover, the seed propagation leads to a very heterogeneous progeny due to cross pollination (Bejoy et al., 2008). A notable variation in the form of colour, quality, yield and the time of first flowering can be observed in the plants which are derived from seeds (Jahan et al., 2009), where the seeds cannot be conserved and must be collected immediately after fruit maturation. Besides, the phase from pollination to seed maturity (approximately 6 to 7 months) and the ensuring development of the plants, which takes a further three years, is too lengthy (Hamidah et al., 1997; Viegas et al., 2007). Seed viability, contamination and germination often creates difficulty and to record high level of germination, seed requires continuous light condition which is hardly available in natural field condition (Gantait et al., 2008), apart from that the seeds are viable only for two to three days and germination is very low (20 to 30%) (Jahan et al., 2009).
PLANT REGENERATION SYSTEMS IN VITRO
Plant regeneration in vitro is an alluring alternative for mass multiplication of outstanding cultivars at faster rates than conventional methods. A unique concept proposed by Haberlandt (1902) and logically proved for the first time by Steward et al. (1958), states that, in vitro culture is one of the key tools of plant biotechnology that exploits the totipotent nature of plant cells. In vitro culture promotes accelerated multiplication of superior clones and is a pre-requisite for the improvement of plants via genetic engineering technique. Tissue culture has been exploited to generate genetic variability by producing haploids, somaclonal and gametoclonal variants from which crop plants can be improved. It is also utilized for enriching the health status of the plant material and to amplify the number of desirable germplasm available to the plant breeder. Tissue culture, in union with molecular tools and techniques, has been successfully used to integrate specific traits through genetic transformation. The culture of single cells and meristems can be effectively used to clean out the pathogens from planting material and thereby dramatically improve the produce of established cultivars. By using certain laboratory materials, typically taking one or two decades to attain the trade market all the way through plant breeding, this technology can be expected to have an ever accelarating impact on crop improvement as we pass through the new millennium (Kothari et al., 2010).
Anthurium is an ornamental plant used for its cut flower significance from commercial to household decoration. Commercial production of the Anthurium plants is growing globally to meet the increasingly high demand of its flowers. Its economic value has extensively increased over the last two decades and there is a growing prospective for continued further growth in both domestic and international markets. To accomplish this purpose and satisfy the increasing demand of innumerable quality propagules, a collection of research work has been carried out to explore morphogenic potential and attain regeneration of Anthurium and constant efforts are still in advancement to achieve further improvement in the biotechnological aspects. Anthurium plant regeneration has been achieved chiefly through direct or indirect organogenesis, but the other modes of regeneration such as somatic and zygotic or somatic embryogenesis have also been explored.
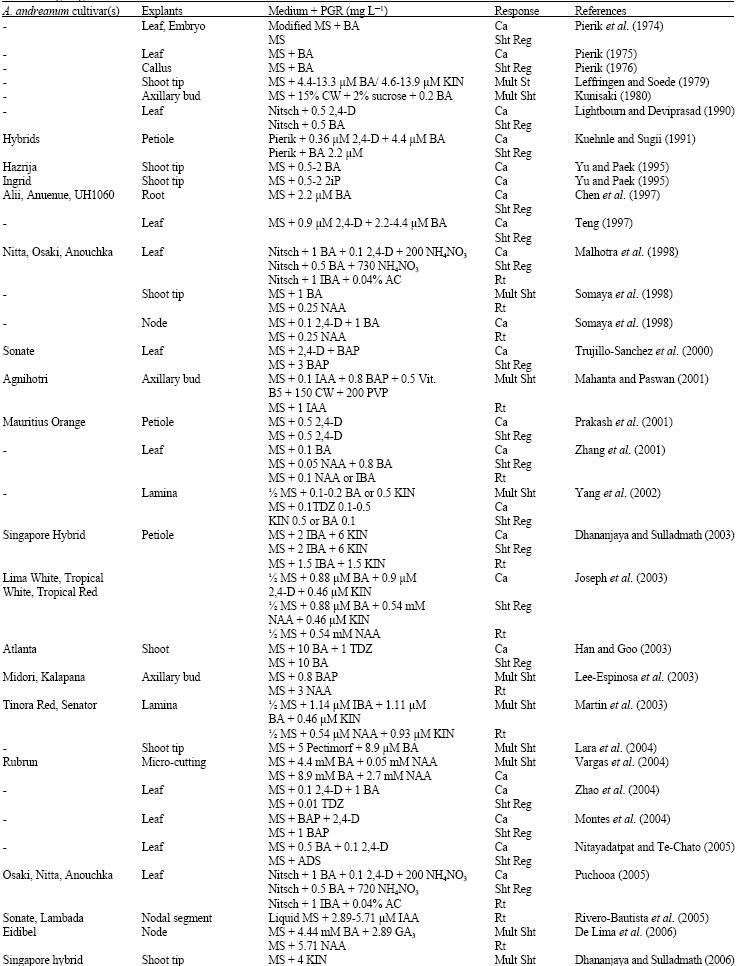
Organogenesis in vitro: Organogenesis in vitro is a versatile inclination concerning the de novo formation of organs (shoots or roots). Shoots can be derived either all the way through differentiation of non-meristematic tissues known as adventitious shoot formation or through pre-existing meristematic tissues known as axillary shoot formation. Both these approaches bring about a synergistic interaction of physical and chemical factors. A triumphant plant regeneration protocol requires an apt choice of explant, age of the explant, specific media formulations, definite growth regulators, genotype, carbon source, gelling agent and other physical factors which includes light regime, temperature and humidity. An array of research work has been performed to explore the morphogenic potential and achieve regeneration of A. andreanum (Table 1).
| Table 1: | Organogenesis in vitro from Anthurium andreanum |
 | |
 | |
| Mult Sht: Multiple shoot; Rt: Root; CW: Coconut water; AC: Activated charcoal; Rt: Root; Ca: Callus; PLB: Protocorm-like bodies; Sht Reg: Adventitious shoot regeneration; CW: Coconut water; AC: Activated charcoal; Nitsch: Nitsch’s basal medium (Nitsch, 1969), Pierik: Pierik medium (Pierik, 1987) | |
Effect of cultivars in regeneration: Cultivars are one of the key attributes that have left their impact on the organogenic response of the cultures in different plant species. The specificity of the cultivars in terms of their individual regeneration capacity stands out as the limiting factor resulting in a condition which demands for specific protocols for each cultivar. The fact concerning the reliability on cultivar specificity for in vitro regeneration acts as one of the most important factors for Anthurium tissue culture.
The outcome of cultivar difference on in vitro morohogenesis has been reported as an obvious result among the Anthurium andreanum cultivars (Pierik et al., 1974). Pierik (1976) examined 38 cultivars of Anthurium andreanum and observed moderate to strong callus formation from leaf explants in 31 responsive cultivars, where callus induction was very meager in four with no response from rest of the three. Performance variation of different cultivars on callus induction and regeneration of Anthurium was studied comprehensively in total 15 different cultivars (Chen et al., 1997; Malhotra et al., 1998; Puchooa, 2005; Duong et al., 2007; Atak and Celik, 2009) using altered explant sources and culture media compositions. In the study of Duong et al. (2007), 10 cultivars namely, Neon, Choco, Sonate, Midori, Pistache, Tropical, Safari, Arizona and Cancan were evaluated on the basis of their performance over callus induction and adventitious shoot formation. In terms of callus induction and proliferation, cultivar Pistache proved as the best resultant, whereas, Carnaval and Cancan didn’t reveal any response. The potentiality of shoot regeneration was also varying from cultivar to cultivar, where, the Tropical type resulted maximum number of shoots than the others. The most recent report by Atak and Celik (2009) in Anthurium andreanum cv. Arizona and Sumi, describes the significantly different impact of cultivars on indirect organogenesis. The callus induction rate of Arizona was significantly higher than that of the Sumi. The average number of adventitious shoots regenerated from callus for Arizona cultivar also increased significantly as compared to Sumi.
Direct regeneration of multiple shoots and roots was notably influenced by the cultivars Midori, Kalapana, Tinora Red, Senator, Sonate and Lambada, as revealed by the literatures (Lee-Espinosa et al., 2003; Joseph et al., 2003; Martin et al., 2003; Rivero-Bautista et al., 2005). In the study of Martin et al. (2003) two commercially elegant cultivars of Anthurium Tinora Red and Senator revealed their differential performance in direct organogenesis from lamina explants. Cultivar Tinora Red proved to be more regenerative than Senator in term of number and growth of meristemoids and multiple shoots per explant though there was no significant difference recorded in rooting in vitro.Kuehnle et al. (1992) were the first to report the influence of Anthurium hybrid cultivars UH780, UH965 and UH1060 on somatic embryogenesis. UH780 and UH956, the two intraspecific hybrids including the tulip-type hybrid UH1060, were cultured on MS medium with varied concentration ratio of 2,4-dichlorophenoxyacetic acid (2,4-D) and 6-furfurylaminopurine (KIN). Impact of different cultivars in production of somatic embryos was quite prominent as UH1060 was the best performer in comparison to the other two.
Effect of explants in regeneration: Direct or indirect in vitro organogenesis of the plant tissue is mostly governed by the suitable selection of the explant. In support to this statement, Anthurium regeneration is also reliant on the age and type of the explant concerned. Different explants including explants such as in vitro seedling, axillary bud, shoot tip, node, leaf, lamina, petiole, spathe and spadix (Pierik et al., 1974; Leffringen and Soede, 1979; Kunisaki, 1980; Kuehnle and Sugii, 1991; Teng, 1997; Joseph et al., 2003; Vargas et al., 2004; De Lima et al., 2006) initiate the purpose efficiently in Anthurium tissue culture.
For direct organogenesis into multiple shoot and root regeneration, shoot tip explants were successfully used (Somaya et al., 1998; Lara et al., 2004; Dhananjaya and Sulladhmath, 2006). The pioneering effort of successful shoot multiplication of Cancan cultivar of Anthurium using apical shoot bud was carried out by Gantait et al. (2008) (Fig. 1a, b). Nodal explant was recognized to be the other most responsive explant to regenerate in vitro roots (Rivero-Bautista et al., 2005). Shoot multiplication was witnessed in three cultivars Agnihotri, Midori and Kalapana, initiating the culture with axillary buds (Mahanta and Paswan, 2001; Lee-Espinosa et al., 2003). Comparable results have been demonstrated by other authors using micro-cuttings of in vitro seedlings explants (Vargas et al., 2004; Maira et al., 2009). Yang et al. (2002) and Martin et al. (2003) fascinatingly studied the efficiency of direct shoot regeneration from lamina explants.
Adventitious shoot and root regeneration subsequent to successful callus induction were studied in detail using leaf explants (Pierik et al., 1974; Lightbourn and Deviprasad, 1990; Malhotra et al., 1998; Trujillo-Sanchez et al., 2000; Zhang et al., 2001; Zhao et al., 2004; Montes et al., 2004; Puchooa, 2005; Nitayadatpat and Te-Chato, 2005; Duong et al., 2007; Viegas et al., 2007; Yang et al., 2008; Beyramizade et al., 2008).
 | |
| Fig. 1: | In vitro propagation of Anthurium andreanum (a) Initiation of shoot bud (b) Multiple shoot regeneration (c) Root induction and elongation (d) Ex vitro acclimation of micropropagated plantlets (e) Micropropagated plants at bloom stage and (f) Gel electrophoresis using ISSR marker [(GT)8A] showing the monomorphic bands ensures clonal fidelity (Lane M- 50 bp ladder, C1 to C7- in vitro regenerated clones and P- mother plant) |
Pierik et al. (1974) led the way out in this aspect. Later, Teng (1997) in a novel approach employed homogenised leaf inoculums to regenerate induced callus and then adventitious shoots along with roots using liquid or raft culture. As stated in the study of Bejoy et al. (2008), callus development was restricted along the cut margins and mid rib zone of leaf lamina explants in the cultivar Agnihotri. Jahan et al. (2009) reported the advantage of leaf explants over spadix both in terms of callus induction and development of multiple shoots. Very recently, efficiency of leaf explants in indirect morphogenesis was assessed by Atak and Celik (2009) in two cultivars of Anthurium andreanum Arizona and Sumi. Callus induction, proliferation and regeneration of shoots were consistently excellent. Efficacy of leaf and petiole over callus-mediated prorocorm-like body (PLB) induction supported by histological details was reported by Yu et al. (2009) in cultivar Valentino. Totipotent callus induction on ½ MS supplemented with 2,4-D and BA was achieved following growth and differentiation of PLBs. In vitro roots also proved to be an attractive explants source for Anthurium micropropagation as they are readily obtainable. Recovery of adventitious plantlets from in vitro root cuttings was efficiently achieved by Chen et al. (1997) in UH1060, Alii and Anuenue cultivars of Anthurium under weak light condition and modified MS with BA supplementation. Apart from substantial achievement of leaf as explants, successful appliance of shoot tip, micro-cuttings, petiole and node as explants, for indirect organogenesis was also reported by several authors (Kuehnle and Sugii, 1991; Somaya et al., 1998; Dhananjaya and Sulladmath, 2003; Han and Goo, 2003; Vargas et al., 2004; Yu and Paek, 1995). Prakash et al. (2001) studied on the regeneration efficiency of petiole explants of A. andreanum cv. Mauritius Orange. Callus was produced initially at the cut ends and later on the entire surface of petiole explants in almost six to eight weeks of culture. Subsequent development of shoots from shoot primordia and later regeneration of roots under in vitro shoots was achieved with effective results.
For somatic embryo development and plant regeneration in A. andreanum, leaf and calli were used as explants to initiate the process (Kuehnle et al., 1992; Prakash et al., 2002; Xin et al., 2006; Sontikul and Te-chato, 2007; Beyramizade et al., 2008). Earlier Kuehnle et al. (1992) developed a successful in vitro protocol of production of somatic embryos and subsequent regeneration of plants from Anthurium hybrids using whole leaf blade. Later on, Xin et al. (2006) have also postulated the superiority of leaf over the other explants like petioles and roots in terms of embryogenic potentiality. Further, this report on efficiency of leaf to develop somatic embryo has been supported by the study of Beyramizade et al. (2008) in an elegant Anthurium cultivar Tera.
It is significant to evaluate key events in somatic embryogenesis with zygotic embryogenesis to establish whether similar developmental events take place in somatic embryos and to consider their quality (Flinn et al., 1991). At present, there are insufficient information on zygotic embryogenesis of Anthurium to make such comparisons and Anthurium genetic improvement. Matsumoto et al. (1998) reported the first work on the ovule or zygotic embryo culture from Anthurium andreanum cv. Kalapana using floral parts as explants. They also analyzed an array of morohological, anatomical and histochemical aspects of zygotic embryogenesis.
Scanning the literature we can conclude that leaf explants have been found to be the most effective for indirect organogenesis (adventitious shoot formation) and plant regeneration in Anthurium, whereas apical or axillary meristems have been most frequently used for clonal multiplication. In addition to this, other explants have also proved their potentiality for regeneration depending upon the parameters like cultivar, plant growth regulator regime and others.
Impact of culture conditions: Kuehnle et al. (1992) explored the importance of light regime on somatic embryogenesis of Anthurium andreanum. They observed that the initiation of somatic embryo requires typical dark period with 23°C temperature, whereas, conservation and maturation of embryos preferred a 16 h photoperiod of 54 μmol/m2/sec and 25±2°C temperature.
Effect of low light intensity in direct regeneration from in vitro root was assessed by Chen et al. (1997), where as low as 4 μmol/m2/ sec illumination successfully incubated the root explants. Martin et al. (2003) observed that the effectiveness of direct plant regeneration was significantly influenced by the pH of the medium. The effect of pH was studied by culturing young brown-coloured lamina explants on the optimal medium. Of the different regimes of pH (5.0 to 6.0) tested in their study, pH at 5.5 developed a higher number of shoots directly from the lamina explant. Explants cultured on medium with lower or higher regimes of the optimal pH exhibited a higher percentage of yellowing and browning.
Duong et al. (2007) observed the effect of pH on different phases of organogenesis, where pH 6.0 was optimum for all stages except in vitro rooting in pH 6.2. In their study, both callus multiplication and plantlet regeneration were achieved at 10 h photoperiod with 45 μmol/m2/sec illumination at 25±2°C and 70-75% RH.
In contrast to that Bejoy et al. (2008) reported the effectiveness of continuous dark period over alternative light and dark period in callus induction of Anthurium. Callus induction frequency was quite high in dark regime, while the explants kept in light didn’t develop callus and turned brown rapidly. In accordance with the above finding, Atak and Celik (2009) also preferred the role of continuous dark phase of one month for callus initiation.
A favourable effect of controlled culture condition during Anthurium organogenesis in vitro (pH 5.8), with a temperature range of 25±2°C, 16 h photoperiod, 60% RH and 1500-3000 lux light intensity was studied (Mahanta and Paswan, 2001; Gantait et al., 2008; Jahan et al., 2009). For PLB induction and regeneration, a specific condition at 25±2°C with 12 h photoperiod at 25-30 μmol/m2/sec proved to be favourable according to the report of Yu et al. (2009).
Initiation of aseptic cultures: Sterilization: For commencement of aseptic cultures, meticulous information on the physiological status and the susceptibility of the plant species to diverse pathological contaminants are obligatory. In favour of most of the explants, the universally adopted system involves surface disinfection of initial explants with 70% (v/v) ethanol for 1 min followed by 0.1% mercuric chloride (HgCl2) for 2 min (Mahanta and Paswan, 2001) or 0.1% HgCl2 alone for 7-12 min and rinsing in sterile distilled water (Martin et al., 2003; Duong et al., 2007; Bejoy et al., 2008) or treating with 1-3% sodium hypochlorite (NaOCl) for 15-20 min (Teng, 1997; Vargas et al., 2004).
However, Gantait et al. (2008) sterilized the shoot tips using antifungal solution cetrimide for 5 min followed by NaOCl and 0.1% HgCl2. Later, Jahan et al. (2009) effectively used 70% (v/v) ethanol for 1 min, 1.5% NaOCl for 8 min as disinfectant and added 0.01% Tween-20 as surfactant.
Most recently, to diminish the contamination caused by fungus, endogenous and exogenous bacteria, Atak and Celik (2009) surface sterilized the explants for 1 min in 70% (v/v) ethanol, soaked in gentamicin solution for 30 min and then again soaked in 20% (v/v) commercial bleach [containing 5% (v/v) NaOCl] for 12 min. However, soaking in solution of gentamycin after surface sterilization resulted in the maximum percentage of disinfected explants. Such discrepancy could be clarified as all through surface sterilization, the bare ends of the explants are separated leaving the fresh conducting tissue, which allows the antibiotic solutions to infiltrate deep within the tissue ensuing in superior rates of disinfection.
Experiment with plant growth regulators: Plant Growth Regulators (PGR) signify a crucial role to pave the way in which plants grow and develop. They control the pace of growth of the individual organs and integrate these organs to generate the plants. There is a synergistic requirement of both auxins and cytokinins to induce cell division and growth in plant tissue cultures. Experiments on whole plants and excised tissues have largely established the existence of antagonistic and additive interactions linking these two types of plant growth regulators (Kothari et al., 2010). Shoot regeneration in A. andreanum is also highly influenced by the media formulations containing plant hormones and other growth regulators.
Pierik et al. (1974) for the first time tested different hormonal regimes for indirect organogenesis in Anthurium. Leffringen and Soede (1979) were the first authors to report the efficiency of cytokinin alone to induce multiple shoots from a single shoot tip explant. MS medium was supplemented with either 4.4-13.3 μM benzyleadenine (BA) or KIN. Soon after, Kunisaki (1980) used Coconut Water (CW) as a growth enhancer successfully to induce multiple shoot buds. Axillary buds were cultured on MS basal with 15% CW and 0.2 mg L-1 BA and a high frequency multiple shoot induction and proliferation were recorded. In accordance with the above findings, later, the report of Mahanta and Paswan (2001) also favoured the role of CW (150 mg L-1) in a combination with vitamin B5 (0.5 mg L-1), Poly Vinyl Pyroledon (PVP) (200 mg L-1) and indole-3-acetic acid (IAA) (0.1 mg L-1) in direct shoot multiplication from axillary bud explants of A. andreanum cv. Agnihotri.
Lightbourn and Deviprasad (1990) introduced Nitsch basal medium (Nitsch, 1969) replacing MS for indirect organogenesis of A. andreanum. A low level of 2,4-D (0.5 mg L-1) alone efficiently induced in vitro callus and its subsequent proliferation, whereas, exactly similar concentration of BA was supplemented alone for successful regeneration of adventitious shoots. With the advancement of the previous idea Kuehnle and Sugii (1991) employed a relatively high level of cytokinin source in combination with low level of auxin to produce more number of calli with a higher proliferation rate in Anthurium hybrids. Pierik basal medium (Pierik, 1987) was fortified with 0.36 μM 2,4-D + 4.4 μM BA when petiole explants were inoculated for dedifferentiation of tissue. For regeneration of shoots from calli, their study supports the reports of their predecessors, BA at 2.2 μM proved to be the best for single cytokinin source. Yu and Paek (1995) established two different protocols in other way to specify cytokinin sources in callus culture for two different cultivars. For Hazrija, MS supplemented with 0.5-2 mg L-1 BA proved most efficient in callus induction as well as growth. On the other hand, MS plus 0.5-2 mg L-1 iP was the best combination for Ingrid cultivar. The effectiveness of cytokinin alone in both callus induction as well as adventitious shoot regeneration was once again proved by the study of Chen et al. (1997) from root explants of Alii, Anuenue and UH1060 cultivars of Anthurium. BA at the rate of 2.2 μM supplementation to MS proved to be the ideal PGR, although response was varied from cultivar to cultivar. Teng (1997) carried out the pioneer work in Anthurium introducing PGRs in liquid or raft culture to improve indirect organogenesis. MS with 0.9 μM 2,4-D + 2.2-4.4 μM BA influenced the inoculums size and regeneration frequency to a great extent. Larger (1000 μm) inoculums produced 1.8-3.3 times higher number of shoots than the smaller ones (500-1000 μm).
Young leaf explants of three A. andreanum cultivars (Nitta, Osaki and Anouchka) were cultured on MS medium with the auxin 2,4-D, indole-3-butyric acid (IBA) and the cytokinin (BA), with an additional supplementation of ammonium nitrate (NH4NO3) and Activated Charcoal (AC). In this unique study of Malhotra et al. (1998) effect of NH4NO3 and AC apart from the regular PGRs was significantly observed particularly for callus induction, adventitious shoot regeneration and in vitro rooting respectively. 200 mg L-1 NH4NO3 in combination with BA and 2,4-D in Nitsch successfully enhanced callus growth and relatively higher level of NH4NO3 (730 mg L-1) induced adventitious shoots from callus. On the other hand, 0.04% AC stimulated a huge success in rhizogenesis in vitro when combined with 1 mg L-1 IBA. The similar study on the same cultivars was carried out by Puchooa (2005) and the details absolutely support the findings of the previous report of Malhotra et al. (1998) regarding the usefulness of 2,4-D, NH4NO3 and AC, except in the recent study it was found that the relatively low level of NH4NO3 (720 mg L-1) was sufficient for shoot regeneration. In a more advanced report of Yang et al. (2008), the noteworthy impact of NH4NO3 on callus induction and proliferation has been observed when supplemented in permutation with CaCl2. In accordance to these reports successive studies of Duong et al. (2007), Gantait et al. (2008) and Atak and Celik (2009), treatment with AC was necessary for root induction and elongation occurring on medium. Combinations of AC and auxin (IAA or IBA) have been very often used as root elongation promoters. However, the addition of AC seems to provide additional beneficial advantage. It is reported that AC eliminates light and provides a reasonable physical environment for the rhizosphere and helps rooting in A. andreanum (Gantait et al., 2008) (Fig. 1c). This statement was further supported by Gantait et al. (2009a, b) in Dendrobium and vanilla respectively, where they observed the enhanced effect of AC in rooting in vitro.
Employment of Adenine Sulphate (ADS), an additive to PGRs, has been shown to promote induction and elongation of shoot buds in A. andreanum (Nitayadatpat and Te-Chato, 2005). MS medium plus ADS devoid of any PGR fruitfully stimulated shoot regeneration. It seems that ADS acts indirectly on bud induction as an elicitor or enhancer of growth in combination or synergism with endogenous or exogenously added growth regulators. Addition of ADS to the medium significantly improved the multiple shoot elongation. Recently, Gantait et al. (2008) emphasized the importance of ADS in the medium for enhanced multiple shoot bud induction and elongation from the cultured shoot tip of A. andreanum cv. Cancan (Fig. 1a, b). The highest number of shoot buds per explant was obtained when MS medium was fortified with 0.5 mg L-1 BAP + 60 mg L-1 ADS.
Yu et al. (2009) reported the upshot of low auxin and high cytokinin levels on callus induction to subsequent PLB production. A half strength of MS with 0.9 μM 2,4-D plus 8.88 μM BA successfully induced callus growth. When the level of BA was decreased to 4.44 μM, it hastened the formation of PLB.
Bud regeneration from somatic embryos cultured in vitro of three hybrid cultivars (UH965, UH780 and UH1060) of Anthurium was studied in detail by Kuehnle et al. (1992). They reported 1-4 mg L-1 2,4-D compiled with 0.33-1 mg L-1 KIN to be the best for somatic embryo development and 0.2 mg L-1 BA to be ideal for adventitious shoot bud regeneration. In support to this, Xin et al. (2006) and recently Beyramizade et al. (2008) reported the similar significance of 2,4-D and KIN combination in embryogenesis and BA alone in shoot regeneration.
Experiments with carbon sources: Plant regeneration in vitro requires the carbon source for induction and proliferation of differentiated or undifferentiated organs. 3% sucrose is the frequent carbohydrate supply in Anthurium in vitro culture (Leffringen and Soede, 1979; Somaya et al., 1998; Yang et al., 2002; Viegas et al., 2007; Gantait et al., 2008; Maira et al., 2009). Muraghige and Skoog (1962) have postulated that, generally the supplementation of 3% sucrose is better than 2 or 4% for the purpose of tissue culture. Nevertheless, there are many accounts on the use of other carbohydrate sources and concentrations for direct or indirect organogenesis in Anthurium. During somatic embryo development of Anthurium Kuehnle et al. (1992) used supplementation of both 1% glucose with 2% sucrose or 3% sucrose, or 4% sucrose and 2% glucose as carbon sources. In this study it was found that a combination of glucose and sucrose in place of sucrose alone significantly improved the somatic embryogenesis of Anthurium, which was entirely supported by the advanced report of Xin et al. (2006) in the similar morphogenetic (somatic embryogenesis) study.
Somatic embryogenesis: Somatic embryogenesis acts as a key component of in vitro propagation when conjugated with a traditional breeding approach and molecular techniques. According to Stasolla and Yeung (2003), somatic embryogenesis provides a precious implement to boost the pace of genetic enhancement of commercial crop species.
Somatic embryos morphologically resemble zygotic embryo but regenerates from somatic cells through a systematic sequence of distinctive morphological stages and is considered profitable over other in vitro propagation systems as it curtails the multiplication time span and proves to be potential as an efficient regeneration system with comparatively high genetic integrity (Kothari et al., 2010). The aforesaid statement certainly is a significant issue in plant tissue culture, which is still progressive in case of A. andreanum (Table 2).
In Anthurium micropropagation, using bulking up via a callus stage, followed by adventitious bud formation, has been proposed as an interesting possibility for large scale production by Pierik et al. (1974) but according to Geier (1990) this method results in formation of off-types. For many applications, somatic embryos have powerful advantages for mass propagation; that is, the high multiplication rate, the ease of use of liquid medium, the handling of enormous numbers of embryos at one time and the possible use of bioreactors (Merkle et al., 1990). As an alternative, most commercial tissue culture laboratories now try to avoid adventitious propagation and favour axillary bud proliferation techniques for Anthurium (Hamidah et al., 1997). However, there are still grave problems with weaning of this material and moreover, the rate of the propagation is time-consuming. As a remedy to the aforesaid problems with conventional in vitro and in vivo propagation techniques, an embryogenic-like callus of A. andraeanum, cultured on medium containing 2,4-D and BA was developed by Kuehnle and Sugii (1991). Plants were readily obtained from that callus but regeneration from somatic embryos was not demonstrated. Afterwards, Kuehnle et al. (1992) reported somatic embryogenesis and plant regeneration in A. andraeanum hybrids using an induction medium containing 2,4-D and KIN.
Direct or indirect somatic embryogenesis and plant regeneration from leaf or in vitro calli using as explants have been achieved by several authors (Prakash et al., 2002; Xin et al., 2006; Sontikul and Te-chato, 2007; Beyramizade et al., 2008). Nitsch’s basal medium (Nitsch, 1969) supplemented with natural additives like coconut water, banana pulp, wheat malt and ragi malt has been observed to promote somatic embryos formation and subsequent shoot regeneration in Liver Red cultivar of Anthurium from leaf explants (Prakash et al., 2002). A higher level of auxin (2,4-D) with comparatively lower level of cytokinin (BA) supplementation in MS have significant effect or even they accelerate the pace of somatic embryogenesis in A. andreanum (Xin et al., 2006). In accordance to the previous report, germination of somatic embryos has been induced by higher 2,4-D and lower KIN level with modified MS, as revealed by the study of Sontikul and Te-chato (2007) and Beyramizade et al. (2008). As understood by all these statements, a very low concentration of BA alone as cytokinin source is established to be potent for shoot regeneration.
| Table 2: | Somatic embryogenesis in Anthurium andreanum |
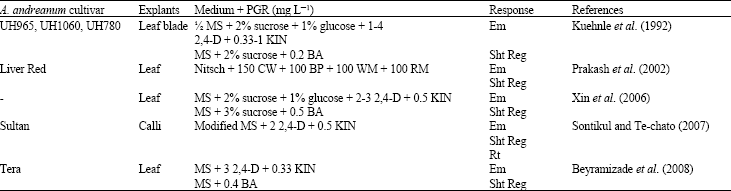 | |
| Rt: Root; Em: Somatic embryo; Sht Reg: Adventitious shoot regeneration; CW: Coconut water; AC: Activated charcoal; WM: Wheat malt; BP: Banana pulp; RM: Ragi (Eleusine coracana) malt; Nitsch: Nitsch’s basal medium (Nitsch, 1969) | |
ACCLIMATIZATION
The in vitro plant regeneration system, an integral part of plant biotechnology, still carries the difficulty of ex vitro acclimatization of plants to triumph over. Acclimatization virtually denotes the climatic or environmental adjustments of an organism, particularly a plant that has been transferred to a new atmosphere. Ex vitro adaptation of a micropropagated plant to a greenhouse or a field setting is indispensable because there is, in general, a noteworthy discrepancy between the in vitro environment and the greenhouse or field condition. Unbeaten acclimatization measures offer most advantageous circumstances for a high proportion of endurance of plants; they diminish the percentage of dead and injured plants in the in vitro process and boost the plant growth and establishment. Ultimately, an efficient acclimatization procedure saves the resources of time, labour, money and cuts off the cost of production of competent and deliverable produce (Gantait, 2009).
Substantial efforts have been directed to optimize the conditions for in vitro stages of micropropagation, but the process of acclimatization of micropropagated plants to the soil environment has not been studied exhaustively in A, andreanum. Accordingly, the transplantation stage continues to be a major bottleneck in the micropropagation of Anthurium. Plantlets regenerated in vitro have been incessantly exposed to a unique microenvironment that has been standardised to offer nominal stress and best possible conditions for plant multiplication. Hazarika (2003) proposed the gradual systematic acclimatization of in vitro generated plantlets in accordance to their physiological and anatomical characteristics.
Mahanta and Paswan (2001) successfully transferred in vitro Anthurium plantlets in the plastic pots containing the growing medium of soilrite-perlite at the ratio of 10:1 and reported 60% survival rate after four weeks of transfer. Effect of vermicompost and sand mixture (1:3 v/v) in ex vitro establishment under green house followed by net house condition was reported by Martin et al. (2003), where 95% survival rate was achieved. Maximum survival rate of 98% was reported by Han and Goo (2003) in cultivar Atlanta using a combination of vermiculite and perlite (1:1 v/v) as growth substrate. Vargas et al. (2004) studied the acclimatization of cultivar Rubrun using soil and organic humus (1:1 v/v). In their study micropropagated plantlets ensured 80% success rate within four weeks. In an exceptional investigation, De Lima et al. (2006) evaluated the associated impact of arbuscular mycorrhizal fungi (AMF) on acclimatization of Anthurium. They reported the symbiotic favour to the plantlets with higher water content and vigorous growth than the control due to multiple inoculations of three AMF species (Glomus etunicatum, Gigaspora albida and Acaulospora longula).Bejoy et al. (2008) used coarse river sand and charcoal (3:1 v/v) mixture for ex vitro transfer and acclimatization of tissue cultured plantlets of Agnihotri which regained growth within four to five weeks with 89% chance of survival. Accordingly, Gantait et al. (2008) studied the effectiveness of organics like charcoal and coconut fibre in a 1:1:1:1 (v/v) combination with sand and soil. The two-step (with six week duration) process of acclimatization, with the success rate of 85% was standardised. The initial sand and soil mixture (1:1 v/v) ensured the primary acclimatization and set the plants to be transferred further (Fig. 1d, e). They also reported the utility of intermittent water spraying to ensure high humidity during this period. More recently, Jahan et al. (2009) established the acclimatization procedure of callus, derived from Anthurium plantlets, on sand, loamy soil and coco-peat (1:1:1 v/v) ensuring 85% survival rate.
Clonal fidelity assessment: There are three major modes of developing an adventitious system suitable for mercantile scales, namely, -organ culture, somatic embryogenesis and nodule culture. George (1996) reviewed these and the other strategies in detail. A key contemplation in using an adventitious system is the prospective of recuperating extraordinarily high numbers of genetic variants (somaclonal variation). In an industrial scenario, this risk is often serious enough to eradicate any further concern of micropropagation as a cloning strategy. This is especially true for suspension/callus-based systems which seed to generate the higher incidences of somaclonal variation in recovered propagules. One of the foremost basis of such a genetic crisis, however, is the unintended occurrence of adventitious bud formation within the shoot cultures (Gantait, 2009).
There are inadequate reports available on clonal fidelity assessment of in vitro raised propagules of A. andreanum. Sontikul and Te-chato (2007) reported the evaluation of genetic identity using isozyme markers. In their study different banding patterns of esterase revealed the somaclonal variation of plantlets regenerated through somatic embryogenesis.
In an exceptional study Gantait et al. (2008) reported the appraisal of genetic clonality of micropropagated plantlets with the aid of molecular markers for the first time in Anthurium. They used ten inter simple sequence repeats (ISSR) markers to assess the uniformity of in vitro generated clones along with their mother plant, where monomorphic ISSR banding pattern revealed that all the micropropagated propagules are genetically true to type (Fig. 1f). The direct organogenesis seems to be the potent factor to restore the genetic integrity. In the recent most studies of Gantait et al. (2010) the efficiency of ISSR in clonal fidelity study on micropropagated Allium ampeloprasum was confirmed successfully.
CONCLUSION AND FUTURE PROSPECT
Noteworthy escalation intended towards in vitro improvement of A. andreanum propagation has been made, but still there is an extensive way to go in this track. Important advances have been materialized in regeneration systems in vitro. Significant efforts have been made to combat various types of hindrances with special emphasis on restoration of genetic integrity and in vitro conservation through long term shoot culture, but the results have not been quite satisfactory as more exhaustive study is expected in this aspect.
We believe that future investigations on the other modes of regeneration (e.g., haploid production, anther culture and protoplast culture etc.) and use of advanced biochemical and molecular markers to assess the differential stages will help to unknot novel mechanisms for the development of more proficient strategies with improved in vitro methods.
REFERENCES
- Atak, C. and O. Celik, 2009. Micropropagation of Anthurium andreanum from leaf explants. Pak. J. Bot., 41: 1155-1161.
Direct Link - Bejoy, M., V.R. Sumitha and N.P. Anish, 2008. Foliar regeneration in Anthurium andreanum Hort. cv. Agnihotri. Biotechology, 7: 134-138.
Direct Link - Beyramizade, E., P. Azadi and M. Mii, 2008. Optimization of factors affecting organogenesis and somatic embryogenesis of Anthurium andreanum Lind. Tera. Propagation Ornamental Plants, 8: 198-203.
Direct Link - Chen, F.C., A.R. Kuehnle and N. Sugii, 1997. Anthurium roots for micropropagation and Agrobacterium tumefaciens-mediated transfer. Plant Cell Tissue Org. Cult., 49: 71-74.
CrossRef - Croat, T.B. and R.D. Sheffer, 1983. The sectional grouping of Anthurium (Araceae). Aroideana, 6: 85-123.
Direct Link - Dhananjaya, M.V. and V.V. Sulladmath, 2006. Rapid and efficient clonal propagation of Anthurium andreanum cv. Singapore hybrid. Ind. J. Hort., 63: 59-61.
Direct Link - Dofour, L. and V. Guerin, 2003. Growth, developmental feature and flower production Anthurium andreanum Lind. in tropical conditions. Sci. Hort., 98: 25-35.
CrossRef - Flinn, B.S., D.R. Roberts, D.T. Webb and B.S. Sutton, 1991. Storage protein changes during zygotic embryogenesis in interior spruce. Tree Physiol., 8: 71-81.
Direct Link - Gantait, S., N. Mandal, S. Bhattacharyya and P.K. Das, 2008. In vitro Mass Multiplication with pure genetic identity in Anthurium andreanum Lind. Plant Tissue Cult. Biotech., 18: 113-122.
CrossRef - Gantait, S., N. Mandal, S. Bhattacharyya and P.K. Das, 2010. Determination of genetic integrity in long-term micropropagated plantlets of Allium ampeloprasum L. using ISSR markers. Biotechnology, 9: 218-223.
CrossRefDirect Link - Gryum, M.H., 1990. Evolution and phylogeny of Araceae. Ann. Miss. Bot. Gard., 77: 628-697.
Direct Link - Hamidah, M., P. Debergh and A.G.A. Karim, 1997. Cyclic somatic embryogenesis of Anthurium scherzerianum Schott. Acta Hort., 447: 123-124.
Direct Link - Han, B.H. and D.H. Goo, 2003. In vitro propagation of Anthurium andreanum 'Atlanta' developed for pot culture. Korean J. Plant Biotech., 2: 179-184.
Direct Link - Hazarika, B.N., 2003. Acclimatization of tissue-cultured plants. Curr. Sci., 85: 1704-1712.
Direct Link - Higaki, T. and H.P. Ramussen, 1979. Chemical induction of adventitious shoots in Anthurium. HortSci., 14: 64-65.
Direct Link - Higaki, T., H.P. Ramussen and W.J. Carpenter, 1984. A Study of Some Morphological and Anatomical Aspects of Anthurium andreanum Lind. University of Hawaii, HITAHR Res. Ser., Honolulu, pp: 12.
Direct Link - Joseph, M., K.P. Martin, J. Mundasssery and V.J. Philip, 2003. In vitro propagation of three commercial cut flower cultivars of Anthurium andraeanum Hortic. Indian J. Exp. Biol., 41: 154-159.
Direct Link - Kobayashi, R.S., J.L. Brewbaker and H. Kamemoto, 1987. Identification of Anthurium andreanum cultivars by gel electrophoresis. J. Am. Soc. Hort. Sc., 112: 164-167.
Direct Link - Kothari, S.L., A. Joshi, S. Kachhwaha and N. Ochoa-Alejo, 2010. Chilli peppers- A review on tissue culture and transgenesis. Biotechnol. Adv., 28: 35-48.
CrossRef - Kuehnle, A.R. and N. Sugii, 1991. Callus induction and plantlet regeneration in tissue cultures of Hawaiian anthuriums. HortScience, 26: 919-921.
Direct Link - Kuehnle, A.R., F.C. Chen and N. Sugii, 1992. Somatic embryogenesis and plant regeneration in Anthurium andraeanum hybrids. Plant Cell Rep., 11: 438-442.
CrossRefDirect Link - Mahanta, S. and L. Paswan, 2001. In vitro propagation of Anthurium from auxillary buds. J. Ornamental Hort., New Seri., 4: 17-21.
Direct Link - Maira, O., M. Alexander and T.E. Vargas, 2009. Micropropagation and organogenesis of Anthurium andraeanum cv. Rubrun. Methods Mol. Biol., 589: 3-14.
CrossRefDirect Link - Martin, K.P., D. Joseph, J. Madassery and V.J. Phillip, 2003. Direct shoot regeneration from lamina explants of two commercial cut flower cultivars of Anthurium andraeanum Hort. In vitro Cell. Dev. Biol. Plant, 39: 500-504.
CrossRefDirect Link - Matsumoto, T.K., A.R. Kuehnle and D.T. Webb, 1998. Zygotic embryogenesis in Anthurium (Araceae). Am. J. Bot., 85: 1560-1568.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nitayadatpat, R. and S. Te-Chato, 2005. Enhanced efficiency for propagation of Anthurium by tissue culture technique. Songkalanakarin J. Sc. Technol., 27: 1003-1008.
Direct Link - Nowbuth, P., G. Khittoo, T. Bahorun and S. Venkatasamy, 2005. Assessing genetic diversity of some Anthurium andraeanum Hort. cut-flower cultivars using RAPD Markers. Afr. J. Bot., 4: 1189-1194.
Direct Link - Pierik, R.L.M., 1976. Anthurium andraeanum plantlets produced from callus tissues cultivated in vitro. Plant Physiol., 37: 80-82.
CrossRef - Pierik, R.L.M., H.H.M. Steegmans and J.A.J. van der Meys, 1974. Plantlet formation in callus tissues of Anthurium andraeanum Lind. Sci. Hort., 2: 193-198.
CrossRef - Puchooa, D., 2005. In vitro mutation breeding of Anthurium by gamma radiation. Int. J. Agric. Biol., 7: 11-20.
Direct Link - Stasolla, C. and E.C. Yeung, 2003. Recent advances in conifer somatic embryogenesis: Improving somatic embryo quality. Plant Cell Tissue Org. Cult., 74: 15-35.
CrossRef - Steward, F.C., M.O. Mapes and J.S. Mears, 1958. Growth and organized development of cultured cells. II. Organization in cultures grown from freely suspended cells. Am. J. Bot., 45: 705-708.
Direct Link - Teng, W.L., 1997. Regeneration of Anthurium adventitious shoots using liquid or raft culture. Plant Cell Tissue Org. Cult., 49: 153-156.
CrossRef - Vargas, T.E., A. Mejias, M. Oropeza and E. de Garcia, 2004. Plant regeneration of Anthurium andraeanum cv. Rubrun. Electron. J. Biotech., 7: 282-286.
Direct Link - Xin, W., B. Xu, G.D. Wang, W.M. Guo, F.D. Wen and J.P. Jin, 2006. Somatic embryogenesis and plant regeneration of Anthurium andraeanum. Acta Hort. Sinica, 33: 1281-1286.
Direct Link - Yang, X.L., Z.F. Hou, J. Ji, S.J. Gui and G. Wang, 2008. Effect of culture medium and temperature on the ratio of callus of Anthurium leaf. J. Shenyang Agric. Univ., 39: 15-18.
Direct Link - Yu, Y., L. Liu, J. Liu and J. Wang, 2009. Plant regeneration by callus-mediatedprotocorm-like body induction of Anthurium andreanum Hort. Agric. Sci. China, 8: 572-577.
CrossRef
sivakumar Reply
this journal is open access so it is very useful
sena bandara Reply
by for tissue culture of anthurium