
A.K.S. Lobato
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
R.C.L. Costa
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
C.F. Oliveira Neto
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
B.G. Santos Filho
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
F.J.R. Cruz
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
J.M.N. Freitas
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
F.C. Cordeiro
Instituto de Ciencias Agrarias, Universidade Federal Rural da Amazonia, Belem, Brazil
International Journal of Botany
Year: 2008 | Volume: 4 | Issue: 2 | Page No.: 231-235







ABSTRACT
The aim of the experiment was to investigate and evaluate morphological changes provoked by the progressive water stress on Leaf Relative Water Content (LRWC), height of plant, number of leaves, shoot dry matter, root dry matter in plants of soybean cultivar sambaiba. The experimental design was at randomized entirely factorial, with 2 hydric conditions (stress and control) and 4 stress points (0, 2, 4 and 6 days). It were showed in the plants under water stress drop in LRWC of 81.5 until 60.9% due water deficiency in substrate, the number of leaves increase at both the treatments, it being showed after of 6 days the increase of 24.2 and 44.8% in the plants under stress and control, respectively. It was showed different behavior of the root dry matter in plants under water stress, with higher root matter until the 2 days and lower matter after of this point, when compared with control plants. The study with soybean cultivar sambaiba under progressive water stress reveal that occurred reduction in the parameters from the experiment beginning, however significant morphological changes in the number of leaves and root dry matter were showed only after 6 days of water restriction.
PDF Abstract XML References Citation
How to cite this article
A.K.S. Lobato, R.C.L. Costa, C.F. Oliveira Neto, B.G. Santos Filho, F.J.R. Cruz, J.M.N. Freitas and F.C. Cordeiro, 2008. Morphological Changes in Soybean under Progressive Water Stress. International Journal of Botany, 4: 231-235.
DOI: 10.3923/ijb.2008.231.235
URL: https://scialert.net/abstract/?doi=ijb.2008.231.235
DOI: 10.3923/ijb.2008.231.235
URL: https://scialert.net/abstract/?doi=ijb.2008.231.235
INTRODUCTION
The soybean is one of the more important crops in the world, it is source of protein in the human food and has been utilized in the formulations of the animals rations, besides utilization of the grain oil. The worldwide yield is leadered by USA, followed of Brazil, Argentina and China, it are responsible for about 90% of the world yield.
It is currently the grain more exported from Brazil, with yield of about 56 millions of ton in 2005, besides it be 33% of Brazilian yield exported mainly to European market (Agrianual, 2006). This crop is economically fundamental to the Brazil by the significant internal consumption and exportation rate greats (Dias, 2004).
The mechanisms utilized by the plants for overcome the water stress effects might be biochemicals, through of the changes in the proline levels (Vendruscolo et al., 2007), hormones in which detach the abscisic acid transport (ABA) from root on direction to shoot (Liu et al., 2005) and physiologics that provoke the stomatal closure and changes on chlorophyll tenors (Chandrasekar et al., 2000), in which it are interconnected and act with the objective of decrease the damages that this abiotic factor can cause for the plant.
According Van Heerden and Krüger (2000), abiotic stresses might are extremely damaging to Glycine max (L.) Merril, it being more sensitive when compared with others food legume as Vigna unguiculata e Phaseolus vulgaris (Roy-Macauley et al., 1992; Silveira et al., 2003) and also with others species as Gossypium hirsutum e Sorghum bicolor (Inamullah and Isoda, 2005; Younis et al., 2000). The water stress might provoke reduction strong in the yield, mainly during the growth and development period (Van Heerden and Krüger, 2002). The aim of the experiment was investigate and evaluate morphological changes provoked by the progressive water stress on Leaf Relative Water Content (LRWC), height of plant, number of leaves, shoot dry matter, root dry matter in plants of soybean cultivar sambaiba.
MATERIALS AND METHODS
Growth conditions and plant material: The experiment was carried out in greenhouse under natural conditions day/night (minimum/maximum air temperature and relative humidity were: 22.4/37.6°C and 68/79%, respectively, verified during the experiment), where the average photoperiod was 12 h and the maximum active photosynthetical radiation of 623 μmol-2 sec-1 (at 12:00 h), located at the Instituto de Ciências Agrária (ICA) of the Universidade Federal Rural da Amazônia (UFRA), city of Belém, state of Pará, Brazil (01°27`S and 48°26`W) during the months of September and October of 2006.
It was used seed of Glycine max (L.) Merril of cultivar sambaiba collected in the 2006 season, from city of Paragominas, state Pará, Brazil (03°00`S and 47°21`W) and stored until carrying out of the experiment. The substrate utilized to the plant growth and evaluation was composed by black potting soil and sand at 3:1 ratio, respectively and the plants grown in pots with 6 L capacity.
Experiment design and treatments: The experimental design was at randomized entirely factorial, with 2 hydric conditions (stress and control) and 4 stress points (0, 2, 4 and 6 days), the experiment was composed by 8 repetitions and 64 experimental units, as well as each repetition had one plant.
Three seeds were placed into each pot and after 7 days, the plants were thinned to one per pot only. The plants remained in greenhouse for 40 days, watered daily and received macro and micronutrients every 5 days, using the nutritive solution of Hoagland and Arnon (1950). Starting 40th day after the implementation of the experiment, the plants from the treatment under stress were submitted to the period of 6 days without irrigation, simulating the water stress until the 46th day. After this period the plants were take away to the Laboratório de Fisiologia Vegetal Avançada belonging to Instituto de Ciências Agrárias (ICA) of Universidade Federal Rural da Amazônia (UFRA) for measure the morphological parameters.
Measurements: It was measured the height of plant and number of leaves, besides it be determined the Leaf Relative Water Content (LRWC) performed with 10 mm disks in diameter, it being calculated as: LRWC = [(FW-DW)/(TW-DW)] x100, where FW is the fresh weight, TW is the turgid weight measured after 24 h of saturation on deionised water at 4°C in the dark and DW is the dry weight determined after 48 h in an oven at 80°C (Slavick, 1979). It was carried out the washing of the root with distilled water and after the plants were divided at shoot and root, it being the fresh matters placed for dehydrate at 65°C for 72 h, for quantification of the root dry matter and shoot dry matter.
Data analysis: The standard error were calculated for each point, it being applied the variance analysis in the results and the averages of the treatments were compared following Tukey test at the 5% significance level, using SAS (SAS, 1996) and based on statistical theories by Gomes (2000).
RESULTS
Leaf relative water content: Occurred significant decrease in the LRWC and it were showed different behaviors among the control and stress treatments, as well as the control treatment was kept with oscillation small and varying of 81.2 until 83.4%. However, the treatment under water deficiency had progressive fall of 81.5 to 60.9% in the LRWC of the 0 to 6 days, respectively. Moreover, it was possible show (Fig. 1) significant difference after of 2, 4 and 6 days under water deficit, when compared with the control plants.
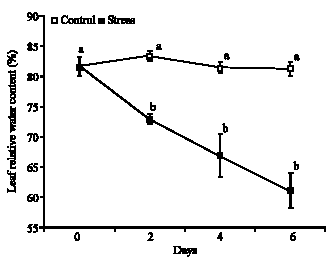 | |
| Fig. 1: | Leaf relative water content in plants of Glycine max cultivar sambaiba under 0, 2, 4 and 6 days of water stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
Height of plant: Significant difference was not observed, according to ANOVA, in which both treatment had increase on height of plant, the plants under water stress had increase of 22.7, 12.8 and 25.2% in the periods from 0 to 2, 2 to 4 and 4 to 6 days, respectively, in total of 60.7% during the 6 days, moreover, the Fig. 2 reveal that of 0 to 6th day, under the same assessment point, the control treatment had higher values that the found in the stress treatment.
Number of leaves: In agreement with variance analysis occurred significant difference among the treatments, it being showed increase on number of leaves in the stress and control treatments, besides not have occurred foliar abscission during the measured period, in which the plants under stress had increase on number of leaves of 20.7, 0.0 and 3.5% during the period between 0 to 2, 2 to 4 and 4 to 6 days, respectively, moreover this treatment have total increase of 24.2%. However, the control treatment had accumulated increase of 44.8%, in which this treatment had the higher number of leaves during the period of 2 to 6 days, when compared with stress, as well as only in the 6th day of water restriction occurred significant difference among the treatments (Fig. 3).
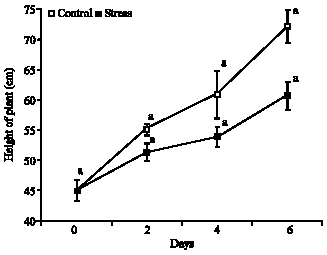 | |
| Fig. 2: | Height of plants on Glycine max cultivar sambaiba under 0, 2, 4 and 6 days of water stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
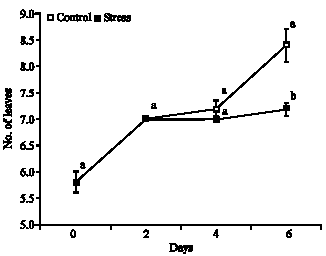 | |
| Fig. 3: | Number of leaves in plants of Glycine max cultivar sambaiba under 0, 2, 4 and 6 days of water stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
Shoot dry matter: It were observed increase in the dry matter of the aerial part in the treatments control and stress, moreover, the ANOVA reveal that occurred not significant difference among the treatments. The control treatment had matter than the stress treatment (Fig. 4), from the beginning until the assessment final, its being showed in the control the accumulated increase de 101.4% during the 6 days, this increase occurred with the values of 8.3, 41.7 and 51.4% in the periods from 0 to 2, 2 to 4, 4 to 6 days, respectively. In the plants under water stress was observed the total increase of 87.4% during the period of 6 days, proving that same under water deficit the studied species had increase of aerial dry mass.
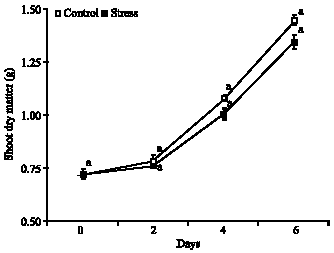 | |
| Fig. 4: | Shoot dry matter in plants of Glycine max cultivar sambaiba under 0, 2, 4 and 6 days of water stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
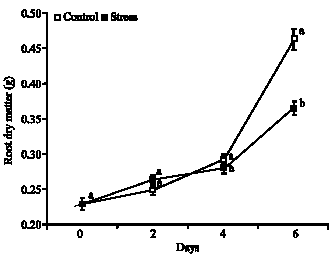 | |
| Fig. 5: | Root dry matter in plants of Glycine max cultivar sambaiba under 0, 2, 4 and 6 days of water stress. Averages followed by the same letter do not differ among themselves by the Tukey test at 5% of probability and the bars represent the mean standard error |
Root dry matter: It were showed significant increases on dry matter of the root of the stress and control plants, moreover, it was showed significant difference only in the 6th assessment day. The plants under water stress had higher root dry matter than the control treatment, until 2 days after the application of the restriction water, in which occurred the increases of the 14.9 and 8.8% on stress and control treatments, respectively, provoked by the higher growth of the root in the plants under light water stress (Fig. 5). From 2nd until 6th day, the root dry matter of the stress plants were smaller, with growth during the periods between 2 to 4 and 4 to 6 days of only 7.0 and 37.7% in plants under water deficit, however the plants kept under irrigation (control) had increase on root dry matter of 18.4 and 75.4% in the interval of 2 to 4 and 4 to 6 days, respectively, its being higher in the two intervals, when compared with the stress treatment. Proving that from 2 days of water deficit the root dry matter of this species is affected negatively.
DISCUSSION
The LRWC decrease provoked by the water deficiency in soil, as well as the root might assimilate lower water amount/volume with increase the period under water stress (Taiz and Zeiger, 1998). Similar results it were found by Sánchez-Rodríguez et al. (1999) investigating plants of Casuarina equisetifoli. The water stress simulated artificially in this experiment cause as direct consequence changes in LRWC, because during the transpiration process and photosynthesis occur water loss through of the stomata and the assimilation/ reposition rate is strongly affected, occurred probably decrease of the conductance stomatal for reduce the water loss to the atmosphere (Verslues et al., 2006).
The lower height and shoot dry matter in the plants under water deficiency occurred, probably due the ABA action, in which it is produced in the cells under abnormal conditions and this way inhibit the cell division and/or DNA synthesis. Similar results on the height reduction in plant under water stress were described by Martínez et al. (2004) studying Atriplex halimus, as well as Lacerda et al. (2001) working with genotypes of Sorghum bicolor under salt stress. Studies carried out by Wang et al. (1998) reveal that the ABA induce the gene expression that codify the inhibited protein of the cyclin-dependent activity (ICK1), coinciding with the results found by Jakoby et al. (2006) on extreme important of this metabolic in cell division process and consequently development and growth plant.
The smaller number of leaves showed in the plants under water stress it is due the drop in LRWC, in which reduce the leaf turgor, the assimilation of water and nitrogen compounds (Reddy et al., 2003) occurred with consequence a lower or void extension rate of the leaf area existent in the plant, moreover probably increase in the abscisic acid levels (ABA) in roots, in which it will are transported from roots to shoot and it will act in the apical region of the plant with antagonist of the auxin and cytokinin, responsible for growth and cell division, respectively (Taiz and Zeiger, 1998), through these hormonal mechanisms the buds remain dormancies and develop not the leaf news. The reduction in the number of leaves in plants under water stress agree with the results got by Ismail et al. (2004) working with Musa sp.
The weight higher of the root dry matter showed in plants under water deficit, in the period between 0 and 2 days of water stress, it can are attribute to root length increased provoked by the search of water in substrate, because with this behavior the plant tempt balance the water supplement, it can make normal the growth rates and tissue developments, besides organize the metabolic activity affected by the water restriction (Pimentel, 2004). These results are similar with found by Sankar et al. (2007) studying the water stress effects on root behavior in Abelmoschus esculentus, in which occurred increase of the root length and consequently weight major under moderate water deficit, however it was showed decrease of the root length under strong water stress, when compared with control plants. According to Kerbauy (2004) studies with gene modified plants describe that occur decrease of the ethylene levels and increase of the abscisic acid (ABA) in roots of the plants under water stress, when compared with plants normally irrigated, it prove the different behavior these hormonals, besides it are attribute at ABA the capacity of the remain ethylene normal levels produced in root of plants under normal conditions. However, the higher root dry matter of the control plants, in finish of the experiment, occurred due at growth of way constant, it being higher at growth of the plants under water stress after of 2 days.
The study with soybean cultivar sambaiba under progressive water stress reveal that occurred reduction in the parameters from the experiment beginning, however significant morphological changes in the number of leaves and root dry matter were showed only after 6 days of water restriction.
ACKNOWLEDGMENTS
The present study make part of Project funded by the Fundação de Apoio à Pesquisa, Extensão e Ensino em Ciências Agrárias (FUNPEA) and counting with the infra-structure of the Universidade Federal Rural da Amazônia (UFRA).
REFERENCES
- Chandrasekar, V., R.K. Sairam and G.C. Srivastava, 2000. Physiological and biochemical responses of hexaploid and tetraploid wheat to drought stress. J. Agron. Crop Sci., 185: 219-227.
CrossRefDirect Link - Inamullah and A. Isoda, 2005. Adaptive responses of soybean and cotton to water stress. Plant Prod. Sci., 8: 16-26.
CrossRefDirect Link - Ismail, M.R., M.K. Yusoff and M. Mahmood, 2004. Growth, water relations, stomatal conductance and proline concentration in water stressed banana (Musa sp.) plants. Asian J. Plant Sci., 3: 709-713.
CrossRefDirect Link - Jakoby, M.J., C. Weinl, S. Pusch, S.J.H. Kuijt, T. Merkle, N. Dissmeyer and A. Schnittger, 2006. Analysis of the subcellular localization, function and proteolytic control of the Arabidopsis Cyclin-dependent kinase inhibitor ICK1/KRP1. Plant Physiol., 141: 1293-1305.
CrossRefDirect Link - Lacerda, C.F., J. Cambraia, M.A.O. Cano and H.A. Ruiz, 2001. Plant growth and solute accumulation and distribution in two sorghum genotypes, under NaCl stress. Rev. Bras. Fisiol. Veg., 13: 270-284.
CrossRefDirect Link - Liu, F., C.R. Jensen, A. Shahanzari, M.N. Andersen and M.N. Jacobsen, 2005. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci., 168: 831-836.
CrossRefDirect Link - Martinez, J.P., S. Lutts, A. Schanck, M. Bajji and J.M. Kinet, 2004. Is osmotic adjustment required for water stress resistance in the Mediterranean shrub Atriplex halimus L? J. Plant Physiol., 161: 1041-1051.
CrossRefDirect Link - Reddy, T.Y., V.R. Reddy and V. Anbumozhi, 2003. Physiological responses of groundnut (Arachis hypogea L.) to drought stress and its amelioration: A critical review. Plant Growth Regul., 41: 75-88.
CrossRefDirect Link - Roy-Macauley, H., Y. Zuily-Fodil, M. Kidric, A.T. Pham Thi and J.V. de Silva, 1992. Effect of drought stress on proteolytic activities in Phaseolus and Vigna leaves from sensitive and resistant plants. Physiol. Plant., 85: 90-96.
CrossRefDirect Link - Sanchez-Rodriguez, J., P. Perez and R. Martinez-Carrasco, 1999. Photosynthesis, carbohydrate levels and chlorophyll fluorescence‐estimated intercellular CO2 in water‐stressed Casuarina equisetifolia Forst. & Forst. Plant Cell Environ., 22: 867-873.
CrossRefDirect Link - Sankar, B., C.A. Jaleel, P. Manivannan, A. Kishorekumar, R. Somasundaram and R. Panneerselvam, 2007. Drought-induced biochemical modifications and proline metabolism in Abelmoschus esculentus (L.) Moench. Acta Bot. Croatica, 66: 43-56.
Direct Link - Silveira, J.A.G., R.C.L. Costa, R.A. Viegas, J.T.A. Oliveira and M.V.B. Figueiredo, 2003. N-Compound accumulation and carbohydae shortage on N2 fixation in drought-stressed and rewatered cowpea plants. Spanish J. Agric. Res., 1: 65-75.
Direct Link - Van Heerden, P.D.R. and G.H.J. Kruger, 2000. Photosynthetic limitation in soybean during cold stress. S. Afr. J. Sci., 96: 201-206.
Direct Link - Van Heerden, P.D.R. and G.H.J. Krüger, 2002. Separately and simultaneously induced dark chilling and drought stress effects on photosynthesis, proline accumulation and antioxidant metabolism in soybean. J. Plant Physiol., 159: 1077-1086.
CrossRefDirect Link - Verslues, P.E., M. Agarwal, S. Katiyar-Agarwal, J. Zhu and J.K. Zhu, 2006. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J., 45: 523-539.
CrossRefPubMedDirect Link - Wang, H., Q. Qi, P. Schorr, A.J. Cutler, W.L. Crosby and L.C. Fowke, 1998. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsis thaliana interacts with both Cdc2a and CycD3 and its expression is induced by abscisic acid. Plant J., 15: 501-510.
CrossRefDirect Link - Younis, M.E., O.A. El-Shahaby, S.A. Abo-Hamed and A.H. Ibrahim, 2000. Effects of water stress on growth, pigments and 14CO2 assimilation in three sorghum cultivars. J. Agron. Crop Sci., 185: 73-82.
CrossRef