
Wafaa A. Al-Kherb
Department of Biology, College of Science and Arts, Qassim University, 1300, Burydah, Qassim, Saudi Arabia
International Journal of Agricultural Research
Year: 2014 | Volume: 9 | Issue: 2 | Page No.: 110-118







ABSTRACT
This study aims to evaluate the effect of different diets on the biology and life table parameters of the minute pirate bug Orius albidipennis (Rueter) for the first time in Saudi Arabia. Immature stages of the aphid species Aphis fabae Scop., A. craccivora (Koch), Myzus persicae (Sulzer) as well as two-spotted spider mite Tetranychus urticae Koch were chosen as preys and offered to nymphs and adults of O. albidipennis at 26°C and 65% RH. Males lived for shorter time than females. The shortest duration of development was achieved when predatory insect was fed A. fabae and A. craccivora. Both aphid species shortened pre-oviposition period and prolonged oviposition which resulted in a higher rate of fecundity. A. fabae was the most suitable food source where it caused the shortest mean generation Time (T) and Doubling Time (DT) while it gave the highest rate of net Reproduction (Ro), intrinsic rate of increase (rm) and finite rate of increase (erm), followed by A. craccivora, M. persicae and T. urticae. These results should contribute to the development of mass rearing programs for biological control agents in developing countries such as Saudi Arabia.
PDF Abstract XML References Citation
Received: November 09, 2013;
Accepted: December 23, 2013;
Published: March 08, 2014
How to cite this article
Wafaa A. Al-Kherb, 2014. Biological Characteristics of Orius albidipennis (Hemiptera: Anthocoridae)
Reared on Insect and Mite Preys. International Journal of Agricultural Research, 9: 110-118.
DOI: 10.3923/ijar.2014.110.118
URL: https://scialert.net/abstract/?doi=ijar.2014.110.118
DOI: 10.3923/ijar.2014.110.118
URL: https://scialert.net/abstract/?doi=ijar.2014.110.118
INTRODUCTION
In commercial greenhouses, aphids (Homoptera: Aphidoidea) are economically one of the most important agricultural pests worldwide, causing direct damage by plant sucking and indirectly as vectors of plant viruses. Their importance is mostly based on their destructiveness to agricultural and ornamental plants. Aphis fabae (Scopoli), Aphis craccivora (Koch) and Myzus persicae (Sulzer) are very important pest species of many greenhouse plants Saudi Arabia. They are phytophagous, cosmopolitan and polyphagous species. Moreover, these aphids are the vectors of many virus diseases in a very large range of plants (Mendes and Bueno, 2001; Bokina, 2008; Sobhy et al., 2010). On the other hand, the two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) is one of the most destructive mites attacking ornamentals and vegetable crops in all continents (Zaher, 1986; Takafuji et al., 2000; Fouly et al., 2011).
Until recent years, frequent sprays with pesticides were needed for pest control especially in greenhouses which resulted in pesticide resistance. Pesticide resistance makes pest control difficult and pesticides create other problems such as the presence of residues in the crops, contamination of the environment and risks for the growers and consumers (El-Maghraby et al., 1994; Abdel-Rahman and Fouly, 2001; Sengonca et al., 2008).
Therefore, biological control as an important component of Integrated Pest Management (IPM) is a reliable crop protection strategy and is economically profitable endeavors for growers of greenhouse crops, mainly in Europe, Asia, North Africa and America (Mendes and Bueno, 2001; Sengonca et al., 2008; Fouly et al., 2011).
The main natural enemies of aphids and spider mites found in greenhouses are the minute pirate bugs Orius spp. (Hemiptera: Anthocoridae) (Lattin, 1999; Silveira et al., 2004). Orius species are polyphagous predators that feed on a wide range of arthropod pests such as aphids, thrips and spider mites in agricultural fields. Their characteristics make them very efficient predators, because they frequently inhabit the same sites as their prey and they can survive sometime without the presence of prey and they relatively easy to mass produce. Furthermore, the interest for Orius spp. as biological control agents has greatly increased (Cocuzza et al., 1997). Orius albidipennis is a commonest Orius species in Mediterranean countries and several studies of its behavior and biological characteristics have been conducted under greenhouse conditions (El-Maghraby et al., 1994; Sobhy et al., 2010; El-Basha et al., 2012). However, no previous information or published data have been recorded on biological characteristics of O. albidipennis with different prey species in Saudi Arabia. Therefore, the objective of the present laboratory study is to determine the developmental duration, mortality, longevity and reproduction of the pirate bug O. albidipennis when fed on A. fabae, A. craccivora, M. persica and spider mite T. urticae as preys. Furthermore, the suitability of these prey species for this predator and their influence on life table parameters as a preliminary step to evaluating its utilization as an efficient biological control agent was tested. Moreover, it is hoped that the present study can elucidate some of the factors responsible for efficacy of O. albidipennis to control different aphid, thrips and mite species on greenhouse plants in Saudi Arabia.
MATERIALS AND METHODS
Aphid cultures: Dry seeds of broad bean Vicia fabae, cowpea Vigna unguiculata subspecies Unguiculata and seedlings (15 cm height) of green pepper Capsicum annuum were transplanted in plastic pots (25 cm in diameter) filled with sandy loam soil and irrigated with water as needed. Thirty days after transplantation, the seedlings were infested with the aphids, A. fabae, A. craccivora and M. persicae, respectively. The infested plants were left for 10 days to encourage the aphids to build up their population and used after that as prey for the predatory pirate bug. Plants were kept under greenhouse conditions of 26±3°C, 65±10% RH.
Spider mite culture: The two spotted spider mite T. urticae individuals were collected from eggplant plants growing in a Greenhouse in Onyzah district in autumn 2012. Mites were reared on bean plants, Phaseolus vulgaris L. potted in plastic pots (25 cm in diameter) in a greenhouse at 26±3°C, 65±10% RH. Spider mites were left to build up their population for 30 days and used after that as prey for the predatory pirate bug.
Orius albidipennis culture: A pure culture of the pirate bug O. albidipennis was established from nymph and adult individuals collected from eggplants (Solanum melongena L.) infested with spider mites and aphids in Onyzah district. Insect population was kept in plastic boxes (20H10H10 cm height) at the same environmental conditions and provided with surplus quantity of preys every other day. Insects were supplied with sufficient amount of spider mites as food source. In each box, a piece of cotton soaked in sugar solution (1:1) and bean pods as a resting and egg laying site (Lundgren and Fergen, 2006). Newly deposited eggs which are partially embedded in plant tissues were collected daily and replaced with new leaves. Rearing boxes were examined daily where number of deposited eggs, incubation period and hatchability were recorded.
Experiment technique: A group of 80 newly hatched nymphs of O. albidipennis were maintained in plastic Petri dishes (10 cm diameter) and covered with covers having a piece of muslin of (2H2 cm) which have been fixed by silicon. This group was subsequently divided into four subgroups, 20 nymphs each and singly provided with surplus of A. craccivora, A. fabae, M. persica and T. urticae. Duration of nymph development, mortality and sex ratio were recorded. As soon as reaching maturity, each female was provided with male where adult longevity, life span, daily rate of deposited eggs were counted and subjected to statistical analysis. In all cases, each rearing arena was provided every other day with a plant leaf infested with one of the aforementioned prey types and kept at the same environmental conditions.
Data analysis: Incubation period, duration during nymph stages and adults, fecundity as well as male and female longevity of O. albidipennis were statistically analyzed by one way ANOVA. Duncan’s Multiple Range Test was used to compare means of different treatments using Costat Software (1990) and followed by Duncan Multiple Range Test.
Life table parameters: Life table parameters of the predatory mite O. albidipennis which was fed on three different prey species, were calculated according to Birch (1948) and Laing (1968) by using the Basic Computer Program Abou-Setta et al. (1986):
Where:
| L | = | No. of alive females |
| x | = | Actual female age (time from egg stage till death) (in days) |
| Mx | = | No. of female progeny/female (mothers)/dayH(specific fecundity rate) |
| Lx | = | Rate of survival at day x |
| Ro | = | Net reproductive rate (ΣLxMx) |
| T | = | Mean generation time (ΣLxMx. x)/(ΣLxMx) |
| DT | = | Time required for the population to double its number |
| GRR | = | Gross reproductive rate |
| rm | = | Intrinsic rate of natural increase |
| erm | = | Finite rate of increase |
RESULTS
Data represented in Table 1-2 showed that there were no significant differences between egg incubation periods for male and female predatory insect, where it ranged from 3.46-4.07 days, respectively. The first nymph of males lived for a longer time when it fed on M. persicae and T. urticae while the second nymphs didn=t show any significant difference among tested prey species. In all cases, immature stages of male O. albidipennis lived for a significant longer time when they were provided with spider mites as prey (Table 1). Moreover, data showed that there were no significant differences between total development of predatory males fed on A. craccivora, A. fabae and M. persica. Similar trend was noticed in the development of female immature stages, where nymphs completed their development feeding on A. fabae significantly faster than those fed on A. craccivora and then followed by T. urticae and M. persica (Table 2). In general, O. albidipennis males completed their development and reached maturity before females.
| Table 1: | Duration (days) (Means±SE) of male developmental stages, life cycle, longevity and life span of Orius albidipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
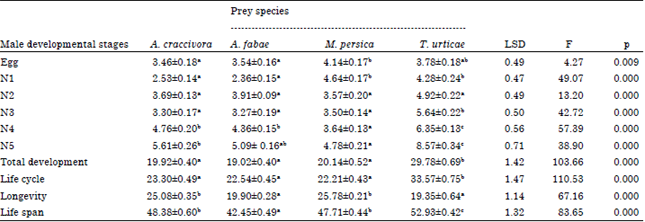 | |
| N1-N5 nymphal stages, means have the same letter in each raw haven=t significant differences (p = 0.05) by Duncan multiple range test, where df = 3, 52 | |
| Table 2: | Duration (days) (Means±SE) of female developmental stages, life cycle, longevity and life span of Orius albidipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
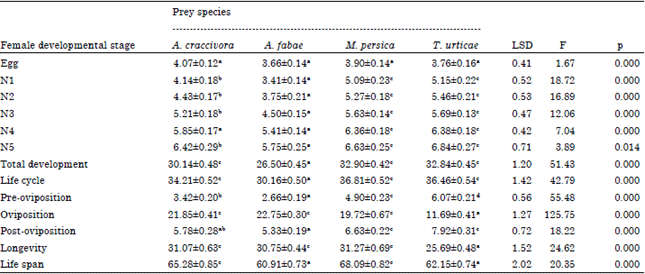 | |
| N1-N5 nymphal stages, means have the same letter in each raw haven=t significant differences (p = 0.05) by Duncan multiple range test, where df = 3, 52 | |
In other words, male longevity averaged 25.08, 19.90, 25.78 and 19.35 days in comparison with females that lived for 31.07, 30.75, 31.27 and 25.69 days when predatory insects fed on A. craccivora, A. fabae, M. persica and T. urticae, respectively (Table 1-2). Adult female of O. albidipennis fed on A. fabae needed 2.66 days before laying its first egg and this period significantly prolonged to 3.42, 4.90 and 6.07 days when it was provided with A. craccivora, M. persica and T. urticae, respectively (Table 2). During an oviposition period of 22.75, 21.85, 19.72 and 11.69 days, each female laid a total number of 66.25, 51.35, 34.27 and 19.23 eggs /female with a daily rate of 2.91, 2.28, 1.72 and 1.61 egg/female/day when it was provided with the aforementioned preys, respectively (Table 3).
Life table parameters: Concerning the life table parameters of O. albidipennis, data represented in Table 4 clarified that sex ratio and nymph survival was 92% when the predator fed on A. fabae and A. craccivora and followed by 86 and 82% when fed M. persica and T. urticae, respectively.
| Table 3: | Mean±SE of total and daily rate of egg deposition of Orius albidipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
 | |
| Means have the same letter in each raw haven=t significant differences (p = 0.05) by Duncan multiple range test, where df = 3, 52 | |
| Table 4: | Life table parameters of Orius albdipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
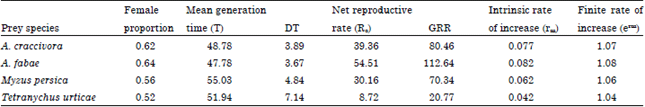 | |
| GRR: Gross reproductive rate, DT: Time required for the population to double its number | |
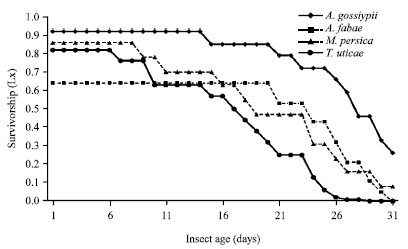 | |
| Fig. 1: | Age specific survivorship of Orius albdipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
The survivorship of O. albidipennis, Lx curves followed I pattern, where most death of O. albidipennis females started after the longest egg laying period (Fig. 1).
Female proportion (females/females+males) was 0.64, 0.62, 0.56 and 0.52, respectively and these values were included in calculating the life table parameters (Table 4).
Mean generation time (T) of O. albidipennis and the time required for the population to double its number (DT) were shorter by feeding on A. fabae and followed by A. craccivora in comparison with M. persica and T. urticae. Net reproductive rate (Ro), the Gross Reproductive Rate (GRR), intrinsic rate of increase (rm) and finite rate of increase (erm) were higher by feeding on A. fabae and A. craccivora while, they were significantly lower when O. albidipennis fed M. persica but T. urticae occupied the last rank (Table 4).
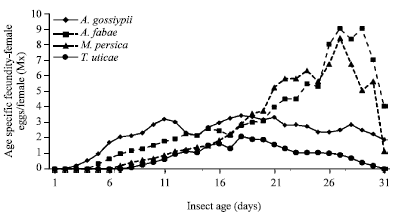 | |
| Fig. 2: | Age specific fecundity of Orius albdipennis fed on Aphis craccivora, A. fabae, Myzus persica and Tetranychus urticae and incubated at 26°C and 65% RH |
As shown in Fig. 2, the age specific fecundity (Mx) of O. albidipennis was highly affected by prey species. However, the net reproductive rates (Ro) averaged 54.51 when the predatory insect fed A. fabae and declined to 39.36 on A. craccivora. These values decreased to 30.16 when M. persica was provided as prey and sharply decreased to reach its lowest value of 8.72 for spider mites, respectively (Table 4). Accordingly, the intrinsic rate of increase rm, finite rate of increase erm increased steadily when predatory insect fed A. fabae, A. craccivora, M. persica and T. urticae (Table 4).
DISCUSSION
Life history of O. albidipennis: However, little is known worldwide and no information or published data concerning the effect of different prey types especially aphids on development and reproduction of O. albidipennis compared with other species of Orius (Sobhy et al., 2010). The present results showed that prey species affected the duration of developmental stages of O. albidipennis male and female. Aphid species shortened significantly the development as compared with spider mite as food source. That may be due to the nutritional value of prey species especially when O. albidipennis nymphs fed A. fabae. These results agree with the findings of Sengonca et al. (2008) who found that the total developmental duration of O. similis Zheng was significantly shorter when feeding on A. pomi (DeGeer) as compared with A. gosypii (Glover) or M. persica. Developmental duration of O. albidipennis male was shorter than female but contradictory, Mendes and Bueno (2001) stated that nymphal development of male and female of O. insidiosus (Say) was equal.
During developmental time, the total mortality percentage of O. albidipennis was strongly affected by prey type. Mortality averaged 8, 8, 14 and 18% when nymphs of O. albidipennis fed A. craccivora, A. fabae, M. persica and T. urticae, respectively. Similarly, Sengonca et al. (2008) found that mortality percentage of O. similis during embryonic development was highly by different prey species. They demonstrated the prey with high nutritional quality increased the weight of eggs which contained a large quantity of yolk and that consequently increased egg hatching. Moreover, Kiman and Yeargan (1985) found that the mortality of O. insidiosus nymphs feeding on thrips was higher than those fed aphids or lepidopterous eggs. According to Tommasini et al. (2004), Ephestia kuehinella (Zeller) eggs induced a significant higher mortality in population of O. majuscules (Reuter) than Frankliniella occidentalis (Pergamde). In all previous studies on Orius species, the highest mortality percentages were recorded by Mendes and Bueno (2001) who found that 31% of 2nd nymphs of O. insidiosus were killed when the insects fed Caliothrips phaseoli (Hood). That means, developmental period of Orius species may be attributed to several factors such as insect species, prey species as well as other environmental conditions.
During adulthood, the present investigations proved that longevity of O. albidipennis was also affected by different prey species where female lived longer than male. The shortest male and female longevity was achieved when adults continue fed spider mite T. urticae while the longest was with M. persica. These results agree with those of Mendes et al. (2002) with O. insidiosus but contradicted with the findings of Fritsche and Tamo (2000) and Tommasini et al. (2004) who found that males of the same species lived longer than females. They attributed the shorter longevity of adult female of O. insidiosus to the higher energy consumption required for egg production and maturation in the ovaries. In contrast, Sengonca et al. (2008) didn't find significant differences between longevity of male and female of O. similis. That means adult longevity of Orius due to species and also may be due to prey type and host plant.
Pre-oviposition period was affected by prey species where O. albidipennis female started laying eggs 2.66 days when fed A. fabae after emergence. This period prolonged significantly by 28.57, 84.21 and 128.19% when female was fed A. craccivora, M. persica and T. urticae. These results agree with those of Mendes et al. (2002) who found that O. insidiosus had a shorter pre-oviposition duration with eggs of A. kuehinella. Contradictory, Sengonca et al. (2008) found that feeding on A. gossypii accelerated the pre-oviposition of O. similis from 4.4-6.2 days with A. fabae as prey species. During an oviposition period, O. albidipennis showed a capability of laying eggs for a longer time when fed on A. fabae which resulted in the largest quantity of eggs. Prey species of A. craccivora, M. persica and T. urticae caused a reduction percentage of fecundity by an average of 22.49, 48.27 and 70.97%, respectively. Similar results were obtained by Chyzik et al. (1995) who reported that fecundity of O. albidipennis was higher when adults of Thrips tabaci (Lindeman) or eggs of E. kuehinella were offered than T. urticae. Similarly, Mendes and Bueno (2001) noticed that female of O. insidiosus fed on C. phaseoli laid an average of 77.8 egg/female, However, a year later, 2002, they found that it considerably increased this value by feeding on eggs of A. kuehinella (195.25 eggs/female) and decreased to 22.50 eggs/female when it fed A. gossypii. On the other hand, although, there are significant differences between the total quantities of eggs laid by O. albidipennis fed on three aphids and spider mites, it was noticed that these variations were limited with regard to the daily rate of fecundity. That may be due to the long of oviposition period. From the previous results, it can be concluded that a suitable prey such as aphid species may allow a high nutritive content which might accelerate the maturation of predator ovarioles which possibly resulted in the short pre-oviposition and long oviposition periods. Moreover, A. fabae showed that it is the most favorable prey to O. albidipennis because it gave the shortest developmental duration, the longest oviposition period and the highest rate of fecundity as compared with other aphid species while spider mite occupied the last rank.
Life table parameters of O. Albidipennis: The percentage of daughter females in the total population of O. Albidipennis averaged 64, 62, 56 and 52% when fed A. fabae, A. Craccivora, M. Persica and T. urticae. The present results also showed that the mean generation time T of O. Albidipennis was affected by prey species when it was incubated at 26°C and 65% RH. The shortest and longest T values were obtained when predatory bug was fed A. fabae and T. urticae. Therefore, the insects needed to 3.67, 3.89, 4.84 and 7.14 days for the population to double its number (DT) when fed A. fabae, A. craccivora, M. persica and T. urticae, respectively. Also, A. fabae provided the pirate bug with the highest rate of net reproductive rate (Ro), intrinsic rate of natural increase (rm), finite rate of increase (erm) and Gross Reproductive Rate (GRR) and followed by A. craccivora, M. persica and finally T. urticae. Thes results showed that A. fabae was the most suitable food source to obtain the best life table parameters which are directly and clearly indicate to the efficiency of the predatory insect as a biological control agent. Similar results were obtained by Mendes et al. (2002) with O. insidiosus, Sengonca et al. (2008) with O. similis and Sobhy et al. (2010) with O. albidipennis.
The present results showed that the different types of prey examined point to the nutritional superiority of aphid species, especially A. fabae, as the prey choice for development and reproduction of O. albidipennis. This is probably due to the high nitrogen content present in such preys which guarantees a more adequate diet for the development of this predator under laboratory conditions. However, in nature, the diversified availability of prey and other factors, in addition to nutritional quality of food, can define the selection process of the predator insect.
ACKNOWLEDGMENTS
The author wishes to thank Dr. Ahmed H. Fouly, Professor of Biological Control and Acarology, Department of Plant Production and Protection, College of Agriculture and Veterinary Medicine, Qassim University to analyze and discuss the life table parameters of O. albidipennis.
REFERENCES
- Abou-Setta, M.M., R.W. Sorrell and C.C. Childers, 1986. Life 48: A basic computer program to calculate life table parameters for an insect or mite species. Florida Entomol., 69: 690-697.
CrossRefDirect Link - Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26.
CrossRefDirect Link - Bokina, I.G., 2008. On the biology of Anthocorid bugs (Heteroptera, Anthocoridae), predators of grass aphids in the forest-steppe zone of West Siberia. Entomol. Rev., 88: 1060-1063.
CrossRef - Chyzik, R., M. Klein and Y. Ben-Dov, 1995. Reproduction and survival of the predatory bug Orius albidipennis on various arthropod prey. Entomologia Experimentalis Applicata, 75: 27-31.
CrossRefDirect Link - Cocuzza, G.E., P. de Clercq, M. van de Veire, A. de Cock, D. Degheele and V. Vacante, 1997. Reproduction of Orius laevigatus and Orius albidipennis on pollen and Ephestia kuehniella eggs. Entomol. Exp. Appl., 82: 101-104.
Direct Link - El-Basha, N.A., M.S. Salman and M.A. Osman, 2012. Functional response of Orius albidipennis (Hemiptera: Anthocoridae) to the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). J. Entomol., 9: 248-256.
CrossRef - El-Maghraby, M.M.A., M.A. El-Tantawy, E.A.A. Gomaa and M. Nada, 1994. Effect of pesticidal application during the early and late cotton season on the abundance of certain predators associated with cotton plants. Anzeiger fur Schadlingskunde Pflanzenschutz Umweltschutz, 67: 60-62.
CrossRef - Fouly, A.H., M.A. Al-Deghairi and N.F. Abdel Baky, 2011. Biological aspects and life tables of Typhlodromips swirskii (Acari: Phytoseiidae) fed Bemisia tabaci (Hemiptera: Aleyroididae). J. Entomol., 8: 52-62.
CrossRefDirect Link - Fritsche, M.E. and M. Tamo, 2000. Influence of thrips prey species on the life-history and behaviour of Orius albidipennis. Entomologia Experimentalis Applicata, 96: 111-118.
Direct Link - Kiman, Z.B. and K.V. Yeargan, 1985. Development and reproduction of the predator Orius insidiosus (Hemiptera: Anthocoridae) reared on diets of selected plant material and arthropod prey. Ann. Entomol. Soc. Am., 78: 464-467.
Direct Link - Laing, J.E., 1968. Life history and life table of Phytoseiulus persimilis Athias-Henriot. Acarologia, 10: 578-588.
PubMedDirect Link - Lundgren, J.G. and J.K. Fergen, 2006. The oviposition behavior of the predator Orius insidiosus: Acceptability and preference for different plants. BioControl, 51: 217-227.
CrossRef - Mendes, S.M. and V.H.P. Bueno, 2001. Biology of Orius insidiosus (Say) (Hemiptera: Anthocoridae) fed on Caliothrips phaseoli (Hood) (Thysanoptera: Thripidae). Neotropica Entomol., 30: 423-428.
CrossRef - Mendes, S.M., V.H. Bueno, V.M. Argolo and L.C.P. Silveira, 2002. Type of prey influences biology and consumption rate of Orius insidiosus (Say) (Hemiptera, Anthocoridae). Revista Brasileira Entomologia, 46: 99-103.
Direct Link - Sengonca, C., K. Ahmadi and P. Blaeser, 2008. Biological charactristics of Orius similis Zheng (Heteroptera, Anthocoridae) by feeding on different aphid species as prey. J. Plant Dis. Protect., 115: 32-38.
Direct Link - Silveira, L.C.P., V.H.P. Bueno and J.C. Van Lenteren, 2004. Orius insidiosus as biological control agent of thrips in greenhouse chrysanthemums in the tropics. Bull. Insctol., 57: 103-109.
Direct Link - Sobhy, I.S., A.A. Sarhan, A.A. Shoukry, G.A. El-Kady, N.S. Mandour and S.R. Reitz, 2010. Development, consumption rates and reproductive biology of Orius albidipennis reared on various prey. BioControl, 55: 753-765.
CrossRef - Takafuji, A., A. Ozawa, H. Nemoto and T. Gotoh, 2000. Spider mites of Japan: Their biology and control. Exp. Applied Acarol., 24: 319-335.
CrossRef - Tommasini, M.G., J.C. van Lenteren and G. Burgio, 2004. Biological traits and predation capacity of four Orius species on two prey species. Bull. Insectol., 57: 79-93.
Direct Link