
Shiferaw Demissie
Department of Biology, Jimma University, P.O. Box 378, Jimma, Ethiopia
Diriba Muleta
Department of Biology, Jimma University, P.O. Box 378, Jimma, Ethiopia
Gezahegn Berecha
Department of Horticulture and Plant Science, Jimma University, P.O. Box 307, Jimma, Ethiopia
International Journal of Agricultural Research
Year: 2013 | Volume: 8 | Issue: 3 | Page No.: 123-136







ABSTRACT
The aim of this study was to evaluate effect of phosphate solubilizing bacteria on seed germination and seedling growth of Faba bean (Vicia faba L.) under lath-house condition. Phosphorous is an essential macronutrient next to nitrogen required by the plants for vital biosynthesis. But often unavailable for plants because of adsorbed by Ca2+, Mg2+, Fe2+ and Al3+ ions through legend exchange. Although there is mounting information that phosphorus solubilizing bacteria as inoculants increases P uptake of plants. This was not yet tested on Faba bean in Ethiopia. A total of 183 phosphate solubilizing bacteria were isolated from 150 samples collected from rhizosphere soil and root nodules. From these isolates based on their solublization index and mobilization efficacy, two isolates (JURB48 and JURMB69) were selected and tested on Faba bean. The co-inoculants (JURB48+JURMB69) increased the percentage of seed germination (21.4%), vigor index (46.2%), radicle (25.3%) and plumule (50%) lengths of germinated seeds of Faba bean over the non-inoculated. Faba bean shoot fresh, root weight, leaf number, flower number, root dry weight and total dry matter were significantly increased compared to non-inoculated as a result of co-inoculants (JURB48+JURMB69). Plant height, root length, phosphorus content, P uptake and Nodule number and weight were enhanced due to inoculation with JURB48 and JURMB69, respectively, compared to non-inoculated either in the presence or absence of phosphate sources. The present study suggests the potential of JURB48 and JURMB69 isolates as biofertilizers for Faba bean cultivation.
PDF Abstract XML References Citation
Received: April 02, 2013;
Accepted: July 30, 2013;
Published: September 07, 2013
How to cite this article
Shiferaw Demissie, Diriba Muleta and Gezahegn Berecha, 2013. Effect of Phosphate Solubilizing Bacteria on Seed Germination and Seedling Growth of Faba Bean (Vicia faba L.). International Journal of Agricultural Research, 8: 123-136.
DOI: 10.3923/ijar.2013.123.136
URL: https://scialert.net/abstract/?doi=ijar.2013.123.136
DOI: 10.3923/ijar.2013.123.136
URL: https://scialert.net/abstract/?doi=ijar.2013.123.136
INTRODUCTION
Faba bean (Vicia faba L.) is commonly named as broad, field and bell bean among most of the world population (Duc et al., 2010). Even if the exact origin of the Faba bean is not known, central Asia, Mediterranean basin and South America were reported as the possible centers of diversity (Muehlauer and Tullu, 1997). Faba bean is widely cultivated in Ethiopia following China in the world (Yirga et al., 2010), in Ethiopia, especially within the range of 1800-2400 m above sea level (Telaye, 1985). It was the third-largest export crop of Ethiopia following coffee and oil seed, between the years 1998-2000. Nearly, Ethiopia exported 88 tons of Faba bean each year to Djibouti, Yemen and Israel (Rashid et al., 2010).
Faba bean is considered as multipurpose crops for its use as human food in developing countries and as animal feed in industrialized countries (Mulualem et al., 2012). It plays a great role in agricultural production as a driver for economic growth and food security by contributing to smallholder income, as a high value crop compared to cereals and to diet as a cost-effective (affordable for smallholders) source of protein, fat, carbohydrate, fibre and minerals like iron, magnesium, potassium, zinc, copper, selenium and various vitamins (Haciseferogullari et al., 2003). In addition, Faba bean contains a chemical called Levodopa which is used for controlling the Parkinson’s symptoms disease (brain neurons disease that affects body movement) (Holden, 2009).
In addition to play a great role in food consumption and therapeutic value, Faba bean is flourish soil fertility due to fixing atmospheric nitrogen which is contribute for decreasing fertilizer expenditure and environmental costs of chemical fertilizers because of the efficacy of Faba bean form association with potential nitrogen fixing bacteria such as Rhizobium leguminosarum biovar viciae (Danso, 1992). Particularly, the augmentation of soil fertility because of Faba bean and Rhizobial association is more significance in extensive area of farming land to boost agronomical product (Habtegebrial and Singh, 2006). Moreover, crop rotation that incorporates the Faba bean will decrease soil-borne disease such as wheat and barley diseases because of Faba bean association with various types of phytobeneficial microorganisms (Agegnehu et al., 2006). In addition to hosting nitrogen fixing bacteria, Faba bean also have a potential to form association with Phosphate Solubilizing Bacteria (PSB) (Keneni et al., 2010). Potent PSB including Pseudomonas spp., Rhizobium spp. and Bacillus spp. are mainly solubilized phosphate pool by production of organic acid which is easily uptake by the plant (Khan et al., 2007; Awasthi et al., 2011).
Currently, the productivity of Faba bean in Ethiopia is far below the expected potential because of low input usage, natural disasters like snowstorm, depletion of macronutrient from cultivable land and unavailability of essential nutrients such as phosphorus. To solve this problem the Ethiopia government has been promoting the practice of using chemical (inorganic) and organic fertilizers with less emphasis to bio-inoculums (Zelleke et al., 2010). However, bio-inoculums are the most important fertilizers that building organic and sustainable agriculture unlike to chemical fertilizers which affect human health and cause environmental pollution (Savci, 2012). Hence, the side effects of chemical fertilizers and problem of low soil fertility could be averted by use of bio-inoculums. Among these, PSB are great gear to pace the turnover of phosphorus from various pools of phosphate to enhance growth and development of crop under question (Khan et al., 2009). However, the potential of PSB inoculation in simultaneous increase of phosphorus uptake and growth of Faba bean is not yet evaluated. To this effect, the present study was designed to evaluate the effects of selected PSB on in vitro seed germination of Faba bean under laboratory condition and seedling growth of Faba bean under lath house condition.
MATERIALS AND METHODS
Description of the study area: The study was carried out at Jimma University, located at 353 km to the southwest of Addis Ababa the capital city of Ethiopia. The microbial analysis and seed germination were conducted in Postgraduate and Research Laboratory of the Department of Biology, Jimma University. The lath house experiment was conducted at Jimma University College of Agriculture and Veterinary Medicine (JUCAVM), Jimma, Ethiopia, the study area is located at 7°, 33’N and 36°, 57’E at an altitude of 1710 m.
Sample size and collection: The rhizosphere soil sample and root nodule were collected from Mana district, Oromia regional state, Ethiopia. We sampled rhizosphere soil and nodule from roots of five plant species; Vicia faba (Faba bean), Pisum sativum (Pea), Phaseolus vulgaris (Haricoat bean), Medicago sativa (Alfalfa) and Sasbania spp. we sampled 30 plants from each of the five plants species. The plant samples were uprooted along with non-rhizosphere soil using shovel. Intact roots along with adhered soil from all of the 5 host plants were transferred to polythene bag and transported to Postgraduate and Research Laboratory, Department of Biology, Jimma University and stored at low (4°C) temperature for further analysis.
Isolation of phosphate solubilizing bacteria: The loosely adherent soil was removed by gentle shaking and the roots were cut into 1-2 cm pieces with strongly adhering soil using sterile scissors. Ten grams of each sample was placed in flasks containing 90 mL of saline solution and kept on rotary shaker at 125 rpm for 30 min. A ten-fold serial dilution was made to obtain appropriate dilution factor. From appropriate serial dilution, a 0.1 mL aliquots was spread on the petriplates containing Pikovskaya’s Agar Medium (PVK) (Glucose 10 g; Ca3 (PO4)2 5 g; NH4(SO4)2 0.5 g; Yeast extract 0.5 g; MgSO4.7H2O 0.1 g; NaCl 0.2 g; KCl 0.2 g; MnSO4 0.002 g; FeSO4 0.002 g; Agar 15 g; distilled water 1000 mL) (Pikovskaya, 1948). The petri plates were incubated at 32°C for 2-7 days. The isolates that showed discrete halo zone around colonies were assumed to be phosphate solubilizers.
Determination of phosphate solubilization index (SI): Phosphate solubilization index (SI) was calculated using the formula outlined in Premono et al. (1996). A loopful of 24 h old cultures was spotted at two point of equidistant on Pikovskaya’s agar plate and incubated at 30±2°C for 2-7 days. The diameter of colony and halo zone was measured using transparent ruler:
Quantification of mobilized phosphate: From 24 h old culture of potent PSB, two loopful of bacterial suspension were inoculated into 200 mL of sterilized National Botanical Research Institute's phosphate growth medium broth (10 g of Glucose, 5 g of Ca3 (PO4)2, 5 g of MgCl2.6H2O, 0.25 g of MgSO4.7H2O, 0.2 g of KCl, 0.1 g of (NH4)2SO4) in 250 mL capacity of Erlenmeyer flask (Nautiyal, 1999). All the inoculated flasks were placed on rotary shaker at 150 rpm for 8 h within days and incubated at 30±2°C. The amount of mobilized phosphate in the broth was estimated from triplicate flasks at 0, 5, 10, 15 and 20 days of incubation with a set of uninoculated control using phosphomolybdic blue color method (Yasmin and Bano, 2011).
Experiment 1 Seed germination bioassay: Twenty one seeds of Faba bean (either single or co-inoculants) were dipped into nutrient broth culture for Pseudomonas sp. (JURB48) and yeast extract mannitol broth for Rhizobium (JURMB69) for 5 h, which contain the bacterial suspension (108 CFU mL-1) while 21 seeds were dipped in distilled water served as control. Seven seeds per plate of inoculated Faba beans were placed in Petri dishes with one layer of Whatman No. 1 filter paper and moist cotton. Both treated and untreated plate containing Faba bean seeds were arranged in Complete Randomized Design (CRD) in triplicate and incubated at 30±2°C for 3-7 days. Then, the percent of germinated seeds for 1-3 days, radicle and plumule length of geminated seeds were taken up to 7 days (Gholami et al., 2009). The vigor index of germinating seeds was also determined using the formula:
Experiment 2 pot experiment: A pot experiment was carried out to investigate the effects of phosphate solubilizing bacteria (JURMB69 and JURB48) with single and dual inoculations in the presence or absence of phosphate sources (Orga and RP) on the Faba bean vegetative growth under lath house condition.
The experiment was performed according to Kucey (1987). Unsterilized soil samples were collected from research site of JUCAVM with no history of Faba bean cultivation for the last 20 years. The soil samples were air dried and busted down to small size. The Bikilal Rock phosphate (BRP) which was brought from western Ethiopia (Wollega) incorporated as source of inorganic phosphate was crushed using hammer. First, the soil and BRP was sieved using 2 mm mesh-size sieve. Second, the sieved soil was filled into 4 kg capacity plastic pots. Then, 2 mm mesh-size of 22 g BRP and 4.4 g of Orga phosphate fertilizer which is made up of bones, stomach paunch, horns and hooves by the action of PSB that contains mainly 1% nitrogen, 23% P2O5, 20.98% organic matter and 21% calcium oxide per 100 kg were added per pot and placed under lath conditions (NAFMAC, 2002). Faba bean variety ‘Dagaga’ obtained from Holeta Research Center, Ethiopia were used. The inoculants in yeast extract mannitol and nutrient broth were mixed with gum arabic in 1:3 ratios and coated onto the seeds (Anonymous, 2012). Sixty seeds of Faba bean for each (JURB48, JURMB69 and JURB48+JURMB69) soaked separately in 60 mL of inoculants-gum arabic mixed suspension for 5 h and pelleting was done by CaCO3. The treated seeds were kept overnight in the laminar flow hood for drying but 60 seeds were sown without inoculants (control). After seed inoculation, 5 seeds were sown into each pot and thinned to three per pot after 7 days of emergence. Three plants per pot were kept upto 45 days. After 15 days of emergence, each seedling of Faba bean was inoculated with 1 mL (about 108 cells mL-1) of their respective inoculants and watered every 2 days until 45 days. The experimental design for the study was Randomized Completely Blocks Design (RCBD) having sixteen treatments replicated three times.
Data collected: Faba bean plants were harvested after 45 days of sowing and the data on plant height, root length, number of nodules and leaves, plant biomass and phosphorus content were recorded following standard procedures (Tennakoon, 2007).
Biomass measurements: At the end of the experiments the root length of Faba bean was carefully uprooted and washed to remove soil. Then, the root length was recorded by measuring the length from base of stem to the tip of longest root. The fresh weight (g) of root and nodules were also measured. Moreover, the fresh weight (g), height (cm), leaves and flower numbers were recorded. Nodules, roots and shoot portions were separated from plants and air dried. The air dried plant samples further oven dried at 72°C for 72 h. The nodule, root and shoot dry weights were measured separately and expressed in g per plant.
Phosphorus content: The oven dried plant samples were ground to fine powder for estimation of phosphorus contents. To 500 mg of plant sample, 10 mL of conc. HNO3 was added in a 250 mL conical flask. The flask was swirled to moisten the entire sample and placed on a hot sand bath for 30 min on the electric hot plate at 180-200°C. The suspension was boiled until nearly to dryness. Five milliliter of tri acid mixture (Conc. HNO3, H2SO4 and 60% HClO4 (10:1:4 ratio)) was added to pre-digested sample and further digested at 180-200°C on a digestion mantle until the residue in the flask became clear white. Ten milliliter of 6 N HCl was added to residue and swirled well. The acid digest was transferred to 50 mL volumetric flask and the volume was adjusted to 50 mL with distilled water. Then, 5 mL of wet oxidized digested sample was taken in a 50 mL of volumetric flask and 10 mL of vanadomolybdate reagent was added. The volume was topped up to 50 mL with distilled water and allowed to react for 30 min. The development of yellow color was read at 490 nm using spectrophotometer.
The P content was obtained by standard curve. To prepare a standard curve, 0.439 g of KH2PO4 oven dried was dissolved in distilled water and adjusted to 1000 mL in a volumetric flask (100 ppm P-solution). Aliquots of 1-10 mL were transferred to 50 mL volumetric flask and 10 mL vanadomolybdate reagent was added to each flask including blank. The volume was made up to 50 mL with distilled water. The development of yellow color was read at 490 nm using spectrophotometer. The standard curve was obtained by plotting a graph as concentration along X-axis and corresponding absorbance along Y-axis (Jackson, 1973).
Phosphorus uptake: Phosphorus uptake in plants was calculated according to Tennakoon (2007):
Statistical analysis: For each of the response variables, the validity of model assumption was verified by examining the residuals. The data of seed germination, fresh and dry biomasses of Faba bean were analyzed using SAS version 9.0. Moreover, the data of radicle and plumule length of germinated seeds didn’t meet model assumption and transformed using Arcsine x. while vigor index of seed germination was transformed by square root. Means separation were performed for significant parameters at p<0.05 using Tukey’s test.
RESULTS
Isolation of phosphate solubilizing bacteria: A total of 183 PSB were isolated from 150 rhizosphere samples and root nodules. Based on their SI and phosphate mobilization efficiency the two isolates namely JURB48 (Pseudomonas sp.) and JURMB69 (Rhizobium sp.) were selected for seed germination and pot experiment.
Effect of PSB inoculation on seed germination: The results of germination percentage, radicle and plumule length of Faba bean inoculation with co-inoculants (JURB48+JURMB69), JURMB69 (Rhizobium sp.) and JURB48 (Pseudomonas sp.) showed significant (p<0.05) increment over control (Fig. 1). Accordingly, co-inoculants and JURMB69 increased seed germination over control by 21.4 and 19% and radicle by 25.3 and 8.8%, respectively. On the other hand, the plumule length increased by co-inoculants (50%) and JURB48 (37.5%) over non-inoculated (Table 1).
 | |
| Fig. 1: | Germinated seeds of Faba bean; JURB48+JURMB69 (co-inoculants), JURB48 and JURMB69 (single inoculants) and control |
| Table 1: | Effect of phosphate solubilizing bacteria on germination of Faba bean seeds |
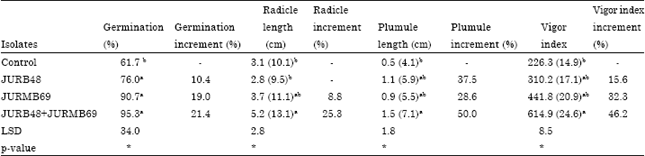 | |
| Values in the parentheses are transformed data, Means with the same letter in the columns are not significantly different, LSD: Least significant difference, *Significance p<0.05, JURB48, isolates from rhizosphere of Faba bean, and JURMB69, isolates from Faba bean root nodule. For all isolates the leading letter JU: Jimma University | |
Effect of PSB inoculation on seed germination and seedling growth of Faba bean fresh biomass: The results of Faba bean inoculation with Pseudomonas sp. and Rhizobium sp. isolates in the presence and absence of phosphate sources on growth parameters are presented in Table 2. All of the inoculated treatments significantly (p<0.05) increased plant height compared to the control (Fig. 2). Among the treatments, OG (Orga)+JURB48 (Pseudomonas sp.) gave the maximum plant height (46.22 cm) followed by BRP (Bikilal Rockphosphate)+OG+JURMB69 (Rhizobium sp.) (44.89 cm). In case of shoot weight, the plant supplemented with OG+JURB48+JURMB69 gave the maximum weight of shoot (23.38 g plant-1) compared to the other treatments (Table 2).
Most of the plants inoculated with PSB produced the higher number of leaves compared to non-inoculated. The maximum number of leaves were observed on the plant inoculated with BRP+OG+JURB48+JURMB69 (43.6) followed by JURMB69 (43.33) without any phosphate sources (Table 2). Significant (p<0.05) differences in both root length and fresh weight of Faba bean were observed due to inoculation with PSB. The plant inoculated with BRP+JURB48 and JURB48+JURMB69 showed the highest root length (36.61 cm) and weight (15.19 g plant-1), respectively.
| Table 2: | Effect of PSB inoculants on growth of Faba bean |
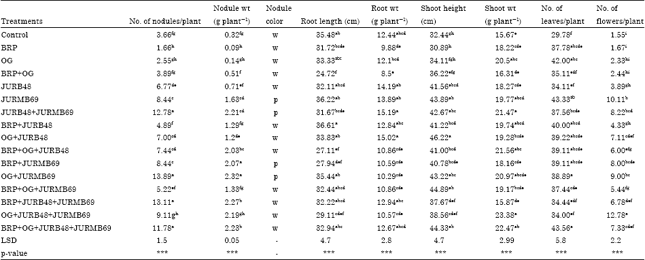 | |
| Means with the same letter in the columns are not significantly different, LSD: Least significant difference, ***:Significant at p<0.001, BRP: Bikilal rock phosphate, OG: Orga, JURB48: Pseudomonas isolated from Faba bean rhizosphere, JURMB69: Rhizobium isolated from nodule of Faba bean, W: White nodule, P: Pink nodule, for all isolates the leading letter JU: Jimma university | |
 | |
| Fig. 2(a-b): | Growth of Inoculated Faba bean (a) Inoculated and (b) Control |
 | |
| Fig. 3(a-b): | Effect of PSB inoculation on Faba bean flowering, (a) Inoculated and (b) Control |
All the inoculated treatments gave the highest number of nodule compared to control (Table 2). Similarly, most of the treatments gave higher nodule fresh weight than the control. The plants inoculated with OG+JURMB69 showed the largest number of nodule (13.89/plant) and weight (2.32 g plant-1) compared to the other treatments and control.
The inoculants also enhanced early flowering and more number of flowers per plant in all inoculated treatments than control (Fig. 3). Particularly OG+JURB48+JURMB69 (12.78/plant) increased number of flowers four fold over the uninoculated plant.
| Table 3: | Effect of PSB inoculants on dry matter content and nutrient uptake of Faba bean |
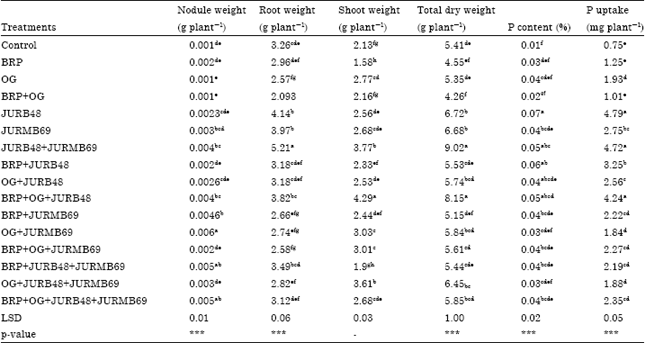 | |
| Mean with the same letter are not significantly different in the columns, LSD: Least significant difference, ***Significant at p<0.001, BRP: Bikilal rock phosphate, OG: Orga, JURB48: Pseudomonas isolated from Faba bean rhizosphere, JURMB69: Rhizobium isolated from nodule of Faba bean. For all isolates the leading letter JU: Jimma university | |
Dry matter biomass: The results on shoot, root, nodule, total dry weight, P content and up take of plants are presented in Table 3. Most of the inoculated treatments recorded significantly (p<0.05) higher shoot dry weight over uninoculated control. BRP+OG+JURB48 recorded the maximum shoot dry weight (4.29 g plant-1). On the other hand, majority of the inoculated treatments enhanced root dry weight significantly over the uninoculated control. The highest root dry weight was recorded for plants inoculated with JURB48+JURMB69 (5.2 g plant-1). Almost all inoculated treatments gave significantly (p<0.05) higher dry mass of root nodules compared to the control. However, BRP+JURB48 and BRP+OG+JURMB69 are at par with the control. The treatments inoculated with OG+JURMB69 recorded the maximum nodule dry weight (0.006 g plant-1) compared to the control.
All the inoculated treatments gave significantly higher total dry matter than uninoculated control, except BRP+JURMB69 which is at par with control. Among the inoculated treatments, JURB48+JURMB69 showed the maximum total dry matter content (9.02 g plant-1) (Table 3). Most of the inoculated treatments gave significantly (p<0.05) the greatest P content in Faba bean compared to the control. Among the inoculated treatments, the highest percentage of P content was recorded in the presence of JURB48 (Table 3). Similar to P content, most of the inoculated treatments showed the greatest P up take in Faba bean. The maximum P uptake was recorded in plants inoculated with JURB48 (4.79 mg plant-1) followed by JURB48+JURMB69 (4.72 g plant-1) (Table 3).
DISCUSSION
In the present study, the co-inoculants (JURB48+JURMB69) increased the percent of seed germination, which is comparable with the result of Gholami et al. (2009) who reported the increment of maize seeds germination by (18.5%) over control due to plant growth promoting rhizobacteria inoculants. The increment of seed germination with inoculants could be due to the isolates ability to synthesis seed germination hormone. Gholami et al. (2009) demonstrated the enhancement of seed germination by inoculants because of the synthesis of seed germination hormone like gibberellins which triggered the activity of specific enzymes that promoted early germination, such as α-amylase that increase the availability of starch assimilation. Furthermore, the co-inoculants increased vigor index, radicle and plumule length compared to control. This result is in accordance with the report of Sharma et al. (2007) in which the PSB co-inoculants (P. fluorescens and B. megaterium) increased percent of radicle and plumule length ranging from 13.8-59.7 and 19-56.4%, respectively. Aipova et al. (2010) also reported the co-inoculation of PSB enhanced the radicle and root length of wheat as compared to individuals. This could be because of higher amount of growth promoting substances and biocontrol substances released by inoculants. Egamberdiyeva (2007) and Sharma et al. (2007) attributed the enhancement of seed parameters to auxins (IAA) and growth promoting substance produced by PSB inoculants. In general, in addition to phosphate solubilizing, both genera of bacteria isolated from legume plants have ability to improve seed germination.
In pot experiment, the selected inoculants improved significantly the growth of Faba bean compared to uninoculated plant either individually or co-inoculation. Among the treatments, OG+JURB48 (Pseudomonas sp.), BRP+OG+JURMB69 (Rhizobium sp.) and BRP+OG+JURB48+JURMB69 were superior in enhancing shoot height over the other treatments. This could be due to high P-solubilizing and mineralizing ability from P-sources, production of growth promoting substances such as IAA and production of biocontrol compounds like HCN (Aditya et al., 2009; Akhtar and Siddiqui, 2009). The fresh weight and leaves numbers of Faba bean were enhanced with co-inoculants (JURB48+JURMB69) supplemented with OG and BRP. The highest in fresh shoot weight and leaves number could be due to the synergistic effect of co-inoculants for releasing of growth substances as well as mineralization and solubilization of P-sources (Kumari et al., 2009).
The highest root length of Faba bean was attained by JURB48 (Pseudomonas sp.) along with Rock phosphate which recorded 36.6 cm long, this result support our in vitro evaluation of isolates in which high amount of IAA was produced by JURB48 (Pseudomonas sp.). Hence, the enhancement in root length might be due to higher amount of IAA produced by JURB48 (Pseudomonas sp.), mobilization of phosphate from inorganic pool and production of bioactive compound that suppress the Faba bean pathogen (Vassilev et al., 2006).
The co-inoculants (JURB48+JURMB69) significantly increased the root weight of Faba bean than the rest of treatments and this could be due to positive relationship of the inoculated bacteria for improving root weight by accumulation of balanced nutrients in the roots and production of growth regulators hormone like IAA and biocontrol compounds that improved root development, which enhanced water and nutrient uptake and ultimately helped better root development. Our results confirmed the previous findings of Afzal and Bano (2008) who reported large amount of wheat root mass due to co-inoculation of PSB. But, by far less than the result of Qureshi et al. (2011), who recorded 50.54 cm root length and 77.1 g plant-1 root mass of Mung bean (Vigna radiata L.) from the co-inoculation of phosphate solubilizing R. phaseoli and B. megaterium.
The plants inoculated with OG+JURMB69 gave high number of nodules (13.9/plant) and nodule weight (2.32 g plant-1). This could be due to the symbiotic effect of Rhizobium on Faba bean along with organic phosphate. In addition, the nodules showed pink color. The number of nodules recorded in the present finding was less than the report of Workalemahu (2009), who recorded 96/plants. This showed that the isolates were weak in nodule formation.
All of the inoculated plants produced highest number of flowers and flowering at the 38 days after sowing compared to control. Specially, OG+JURB48+JURMB69 treated plants gave significant number flowers (12.78/plant) and this was four times higher than the uninoculated plant. This finding is congruent with the investigation of Kumari et al. (2009), who recorded up to 21 flowers/plants from Lens culinaris inoculated by PSB inoculants. The co-inoculants (OG+JURB48+JURMB69) enhancing the uptake of nutrient from organic phosphate amendment, production of phytohormones and higher biocontrol agent from two sides. Similarly, the earliness of bud initiation in PSB-inoculated plants could be due to easy uptake of nutrients and simultaneous transport of growth promoting substances such as cytokinins to the auxiliary buds resulting in breakage of apical dominance, ultimately this has resulted in a better sink for faster mobilization of photosynthates and early transformation of plant parts from vegetative to reproductive phase as earlier demonstrated by Jayamma (2008).
The overall results on dry matter showed that, most of the treatments receiving inoculants gave highest dry biomass of plant compared to control. This could be due to accumulation of nutrient in the plant tissue either from organic and inorganic source of phosphate or soils due to the activities of PSB, which could have contributed to improve the dry weight of shoot. Most of the inoculated treatment significantly increased dry weight of root over uninoculated control. The highest root dry weight was recorded in plant inoculated with JURB48+JURMB69 (5.2 g plant-1). This could be due to co-inoculants promote the growth of root through nitrogen fixation, phosphate solubilization and production of growth promoting substances which are known to change the root morphology and increase the biomass of Faba bean root. These observations are less than the earlier findings of Qureshi et al. (2011) who reported that co-inoculation of PSB such as Bacillus and Rhizobium sp. significantly increased (231.3 g pot-1) the root biomass of Mung bean (Vigna radiata L.).
The total dry weight of the Faba bean inoculated with PSB was significantly increased over control. For instance the plant inoculated with co-inoculant (JURB48+JURMB69) recorded the maximum dry weight (9.02 g plant-1) of Faba bean compared to the other treatments. This result is in line with the report of El-Komy (2005), who reported increase in the total dry weight of wheat due to the co-inoculation of Azospirillum lipoferum and Bacillus megaterium. These isolates possessed high P-solubilizing ability, nitrogen fixing capacity as well as production of growth hormone and production of biocontrol, which might have contributed to the enhanced plant vigor. Increase in plant height, number of leaves and root growth in inoculated treatments might have led to the increased total dry matter production of Faba bean plants as earlier suggested by Afzal et al. (2005).
The two isolates (JURB48 and JURMB69) either in combination or separately enhanced both P content and uptake of Faba bean plants significantly over uninoculated treatments. The increase in P uptake due to PSB inoculation could be because of the ability to solubilize insoluble inorganic and mineralize organic phosphate sources as a result higher amount of P accumulated in plant tissue. The significant increase in the percentage of P content (0.07) and uptake (4.79 mg plant-1) were observed due to inoculation with isolates of JURB48 (Pseudomonas sp.). It was clearly evident that inoculants which solubilized higher amount of phosphorus enhanced content and uptake of P over the rest of the treatments. Improved in P content due to inoculation with fluorescent pseudomonas was reported by Zargar et al. (2005) in other crop plants. This investigation has clearly indicated that PSB enhanced the P up take in Faba bean plants through multitrophic beneficial traits. However, El-Komy (2005) reported that the inoculation of PSB increased the P content of wheat plant upto 53%. In general, the growth parameters of Faba bean were enhanced either single or co-inoculate when amended with Orga. This enhancement could be due to mineralization of Orga than solubilization of inorganic phosphate. This could be substantiated by the findings of Ekin (2010) who reported that the growth and yield of Sunflower was enhanced due to phosphate fertilizers amended with PSB.
CONCLUSION
In the current study, the co-inoculants (JURB48 and JURMB69) enhanced seed germination, radicle and plumule length significantly compared to the other treatments. From the two inoculants, JURMB69 (Rhizobium sp.) is more effective than JURB48 in enhancing seed germination parameters. Most of the plant biomass such as fresh weight of shoot, root and total dry weight, flowering and leave number were significantly increased as a result of co-inoculants (JURB48 and JURMB69) either with or without of phosphate sources. In contrast to in vitro situation, JURB48 (Pseudomonas sp.) highly increased most biomass and nutrient content of Faba bean compared to JURMB69 (Rhizobium sp.) inoculant. Both single and co-inoculants are more effective with Orga to enhance the growth of Faba bean rather than rockphosphate. Moreover, if the effectiveness of our PSB isolates will be tested on other crops and vegetables, it would be better to reduce dependance on commercial fertilizers.
ACKNOWLEDGMENTS
We would like to thank Jimma University for financial support and arranging all the logistics required for the execution of this study. We gratefully credit the farmer owners of the Faba bean farms for their permission and guidance.
REFERENCES
- Aditya, B., A. Ghosh and D. Chattopadhyay, 2009. Co-inoculation effects of nitrogen fixing and phosphate solubilising micro-organisms on Teak (Tectona grandis) and Indian redwood. J. Biol. Sci., 1: 23-32.
Direct Link - Afzal, A. and A. Bano, 2008. Rhizobium and phosphate solubilizing bacteria improve the yield and phosphorus uptake in Wheat (Triticum aestivum). Int. J. Agric. Biol., 10: 85-88.
Direct Link - Afzal, A., M. Ashraf, S.A. Asad and M. Farooq, 2005. Effect of phosphate solubilizing microorganisms on phosphorus uptake, yield and yield traits of Wheat (Triticum aestivum L.) in rainfed area. Int. J. Agric. Biol., 7: 207-209.
Direct Link - Agegnehu, G., A. Ghizaw and W. Sinebo, 2006. Yield performance and land-use efficiency of barley and faba bean mixed cropping in Ethiopian highlands. Eur. J. Agron., 25: 202-207.
CrossRef - Aipova, R., S.A. Aitkeldiyeva, A.A. Kurmanbayev, A.K. Sadanov and O.B. Topalova, 2010. Assessment of biotechnological potential of phosphate solubilizing bacteria isolated from soils of Southern Kazakhstan. Nat. Sci., 2: 841-845.
CrossRefDirect Link - Akhtar, M.S. and Z.A. Siddiqui, 2009. Use of plant growth-promoting rhizobacteria for the biocontrol of root-rot disease complex of chickpea. Australasian Plant Pathol., 38: 44-50.
CrossRefDirect Link - Awasthi, R., R. Tewari and H. Nayyar, 2011. Synergy between plants and P-Solubilizing microbes in soils: Effects on growth and physiology of crops. Int. Res. J. Microbiol., 2: 484-503.
Direct Link - Duc, G., S.Y. Bao, M. Baum, B. Redden and M. Sadiki et al., 2010. Diversity maintenance and use of Vicia faba L. genetic resource. Field Crop Res., 115: 270-278.
Direct Link - Egamberdiyeva, D., 2007. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol., 36: 184-189.
CrossRefDirect Link - Ekin, Z., 2010. Performance of phosphate solubilizing bacteria for improving growth and yield of sunflower (Helianthus annuus L.) in the presence of phosphorus fertilizer. Afr. J. Biotechnol., 9: 3794-3800.
Direct Link - Gholami, A., S. Shahsavani and S. Nezarat, 2009. The effect of Plant Growth Promoting Rhizobacteria (PGPR) on germination, seedling growth and yield of maize. World Acad. Sci. Eng. Technol., 49: 19-24.
Direct Link - Habtegebrial, K. and B.R. Singh, 2006. Wheat responses in semiarid Northern Ethiopia to N2 fixation by Pisum sativum treated with phosphorous fertilizers and inoculant. Nutrient Cycling Agroecosyst., 75: 247-255.
CrossRef - Haciseferogullari, H., I. Gezer, Y. Bahtiyarca and H.O. Menges, 2003. Determination of some chemical and physical properties of Sakiz faba bean (Vicia faba L. var. major). J. Food Eng., 60: 475-479.
CrossRefDirect Link - El-Komy, H.M.A., 2005. Coimmobilization of Azospirillum lipoferum and Bcillus megaterium for successful phosphorus and nitrogen nutrition of wheat plants. Food Technol. Biotechnol., 43: 19-27.
Direct Link - Jackson, M.L., 1973. Soil Chemical Analysis. 1st Edn., Prentice Hall Ltd., New Delhi, India, Pages: 498.
Direct Link - Keneni, A., F. Assefa and P.C. Prabu, 2010. Isolation of phosphate solubilizing bacteria from the rhizosphere of faba bean of Ethiopia and their abilities on solubilizing insoluble phosphates. J. Agric. Sci. Technol., 12: 79-89.
Direct Link - Khan, M.S., A. Zaidi and P.A. Wani, 2007. Role of phosphate-solubilizing microorganisms in sustainable agriculture-A review. Agron. Sustain. Dev., 27: 29-43.
CrossRefDirect Link - Khan, A.A., G. Jilani, M.S. Akhtar, S.M.S. Naqvi and M. Rasheed, 2009. Phosphorus solubilizing bacteria: Occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci., 1: 48-58.
Direct Link - Kucey, R.M.N., 1987. . Increased phosphorus uptake by wheat and field beans inoculated with a phosphorus-solubilizing Penicillium bilaji strain and with vesicular-arbuscular mycorrhizal fungi. Applied Environ. Microbiol., 52: 2699-2703.
Direct Link - Kumari, M., D. Vasu, Z. Ul-Hasan and U.K. Dhurwe, 2009. Effects of PSB (Phosphate Solubilizing Bacteria) morphological on characters of Lens culinaris Medic. Med. Biol. Forum Int. J., 1: 5-7.
Direct Link - Mulualem, T., T. Dessalegn and Y. Dessalegn, 2012. Participatory varietal selection of faba bean (Vicia faba L.) for yield and yield components in Dabat district, Ethiopia. Wudpecker J. Agric. Res., 7: 270-274.
Direct Link - Nautiyal, C.S., 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett., 170: 265-270.
CrossRefPubMedDirect Link - Qureshi, M.A., M.A. Shakir, A. Iqbal, N. Akhtar and A. Khan, 2011. Co-inoculation of phosphate solubilizing bacteria and rhizobia for improving growth and yield of mungbean (Vigna radiata L.). J. Anim. Plant Sci., 21: 491-497.
Direct Link - Savci, S., 2012. An agricultural pollutant: Chemical fertilizer. Int. J. Environ. Sci. Dev., 3: 77-80.
Direct Link - Sharma, K., G. Dak, A. Agrawal, M. Bhatnagar and R. Sharma, 2007. Effect of phosphate solubilizing bacteria on the germination of Cicer arietinum seeds and seedling growth. J. Herbal Med. Toxicol., 1: 61-63.
Direct Link - Vassilev, N., M. Vassileva and I. Nikolaeva, 2006. Simultaneous P-solubilizing and biocontrol activity of microorganisms: Potentials and future trends. Applied Microbiol. Biotechnol., 71: 137-144.
CrossRefDirect Link - Workalemahu, A., 2009. The effect of indigenous root nodulating bacteria on nodulation and growth of faba bean (Vicia faba) in low input agricultural systems of tigray highlands, Northern Ethopia. Momona Ethiopian J. Sci., 1: 30-43.
Direct Link - Yasmin, H. and A. Bano, 2011. Isolation and characterization of phyosphate solubilizaing bacteria from rhizosphere soil of weeds of khewra salt range and attock. Pak. J. Bot., 43: 1663-1668.
Direct Link